
Pigs Immunized with the Virus-like Particle Vaccine Are Protected against the Hepatitis E-3 Virus

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Selection of the HEV-3-239 Capsid Antigen and Producing Recombinant Baculovirus
2.2. Purification of HEV-239 VLP
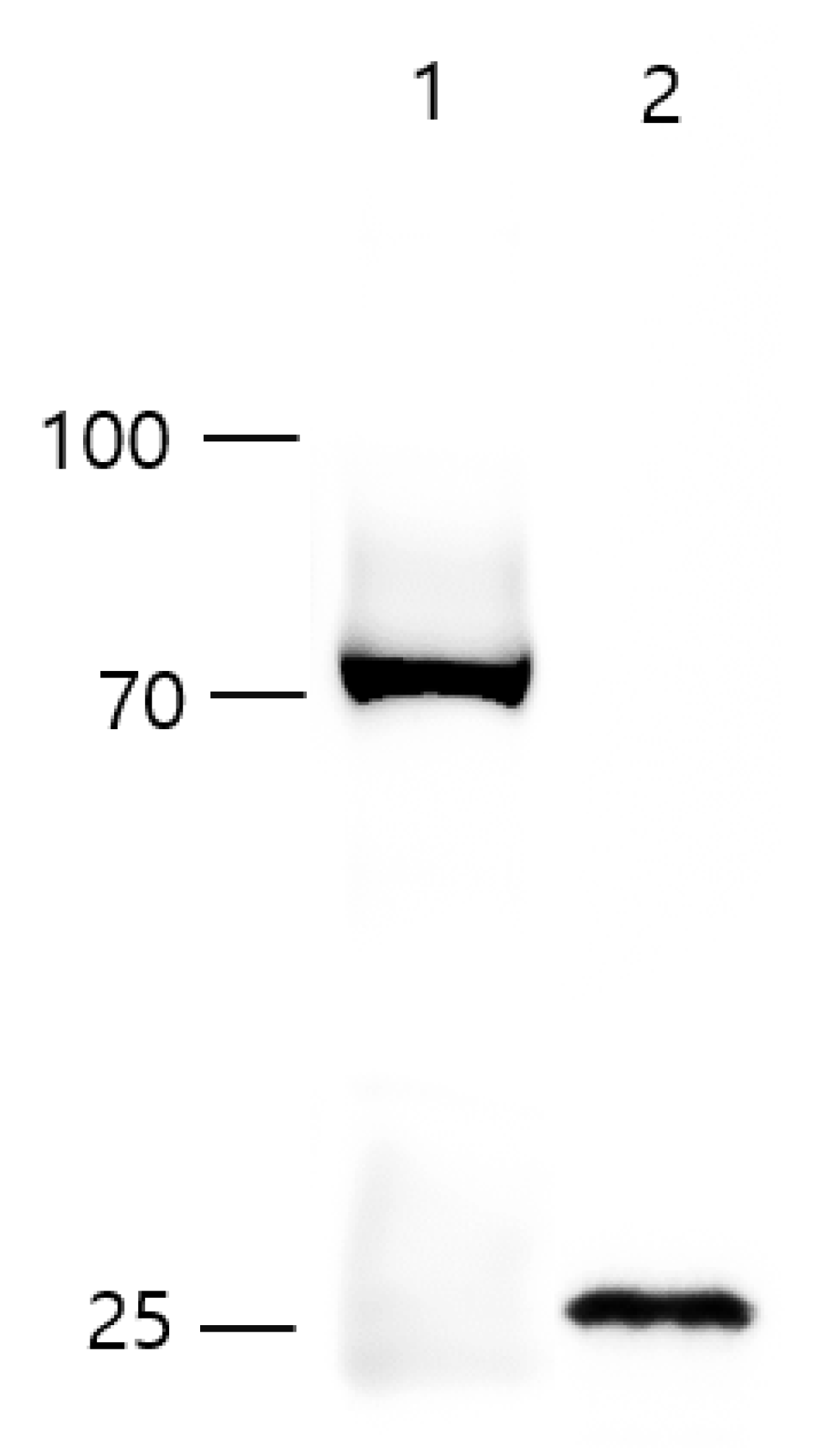
2.3. Western Blot Analysis
2.4. Electron Microscopy
2.5. Animals, Immunization, and Challenge Schedule
2.6. HEV RNA, Anti-HEV IgG Antibody Analysis
2.7. Examination of ALT, AST in the Porcine Serum
2.8. Histopathology
2.9. Statistical Analysis
3. Results
3.1. Icosahedral Shaped, 20–30 nm-Sized, HEV-3-239-VLPs Was Assembled
3.2. Fecal Viral Shedding and Viremia Were Not Detected in the Pigs Vaccinated with 200 μg of VLPs
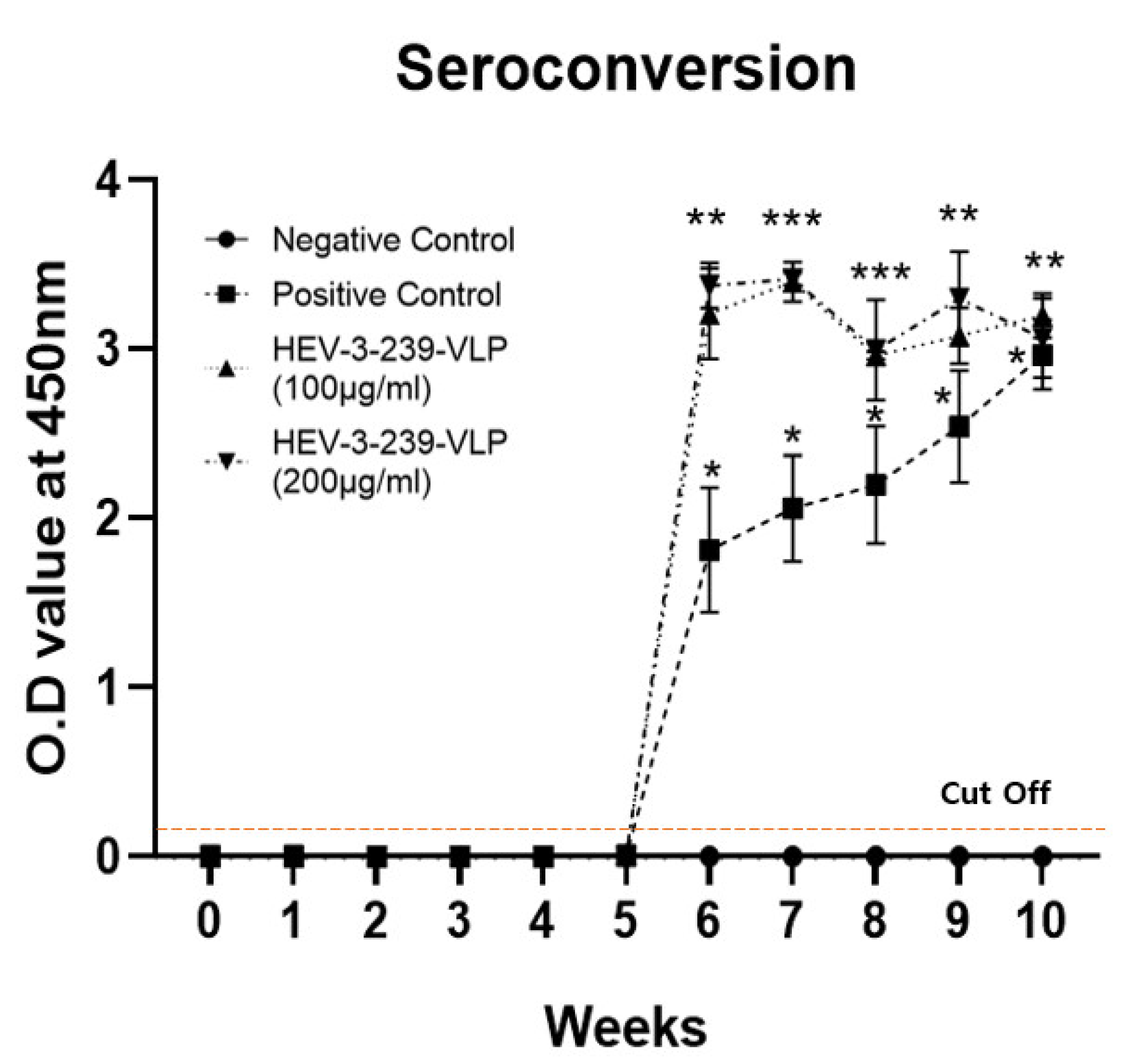
3.3. Vaccination with the HEV-3-239-VLP Induced the Anti-HEV Antibodies
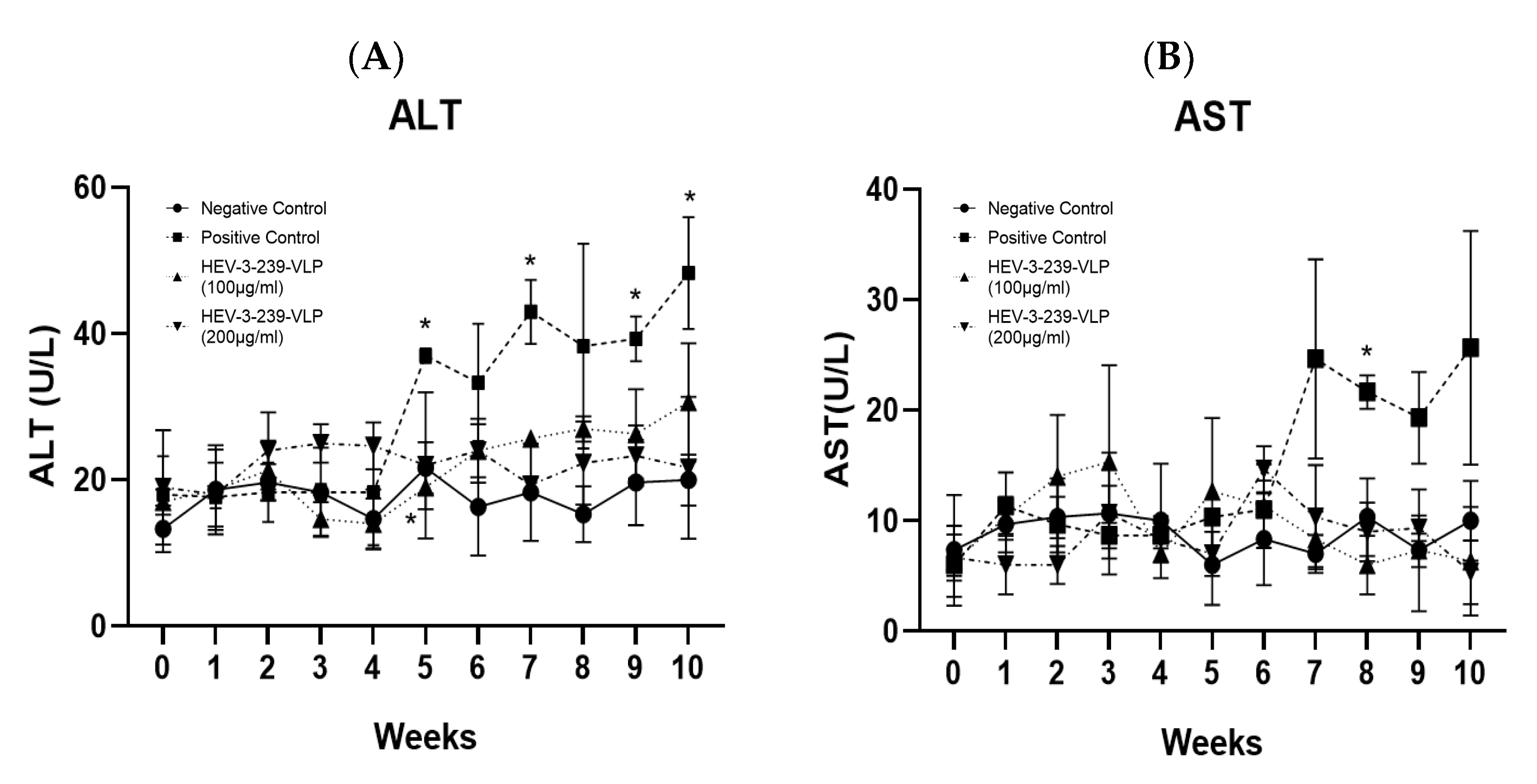
3.4. HEV-3-239 VLP Prevents Hepatitis
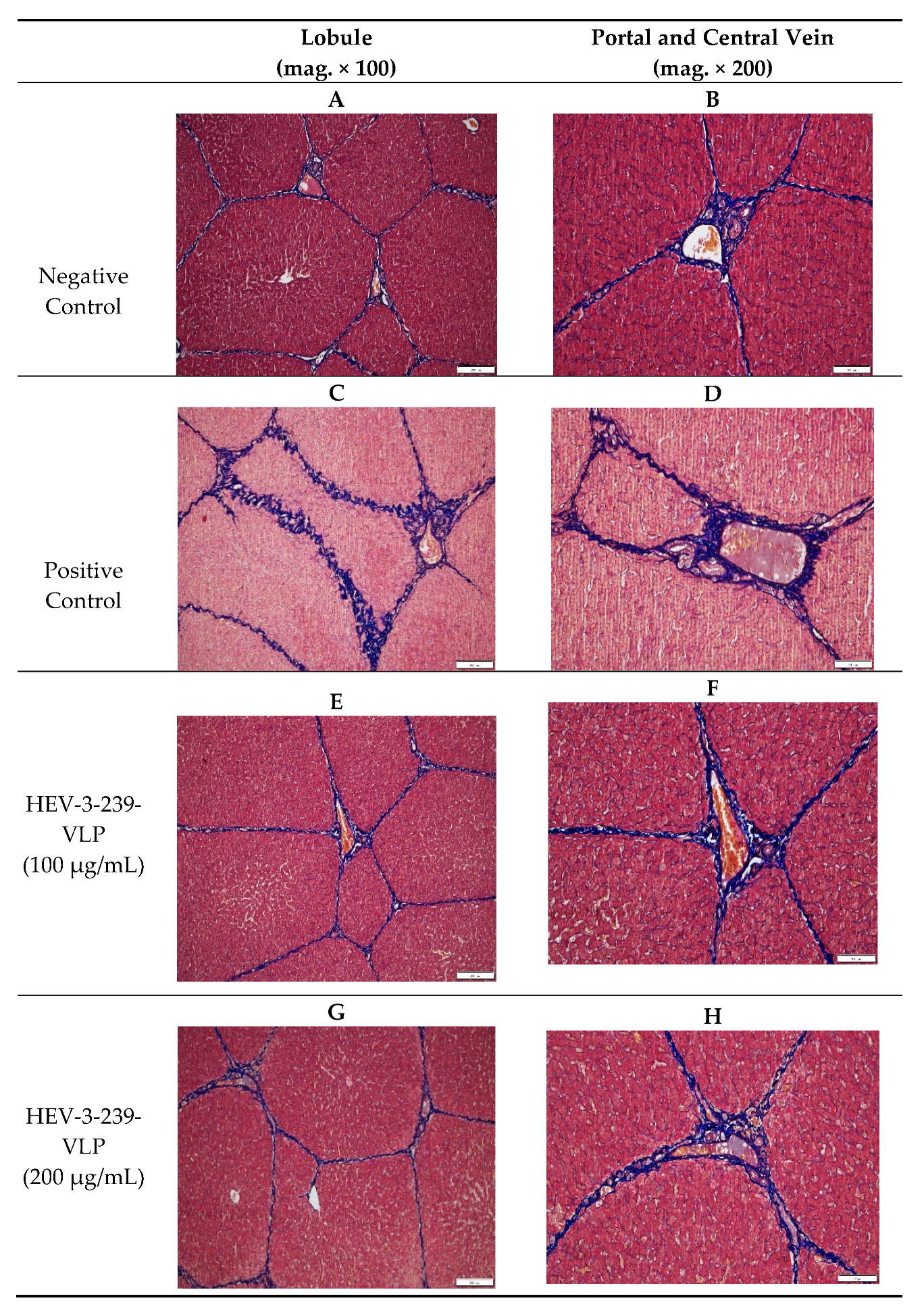
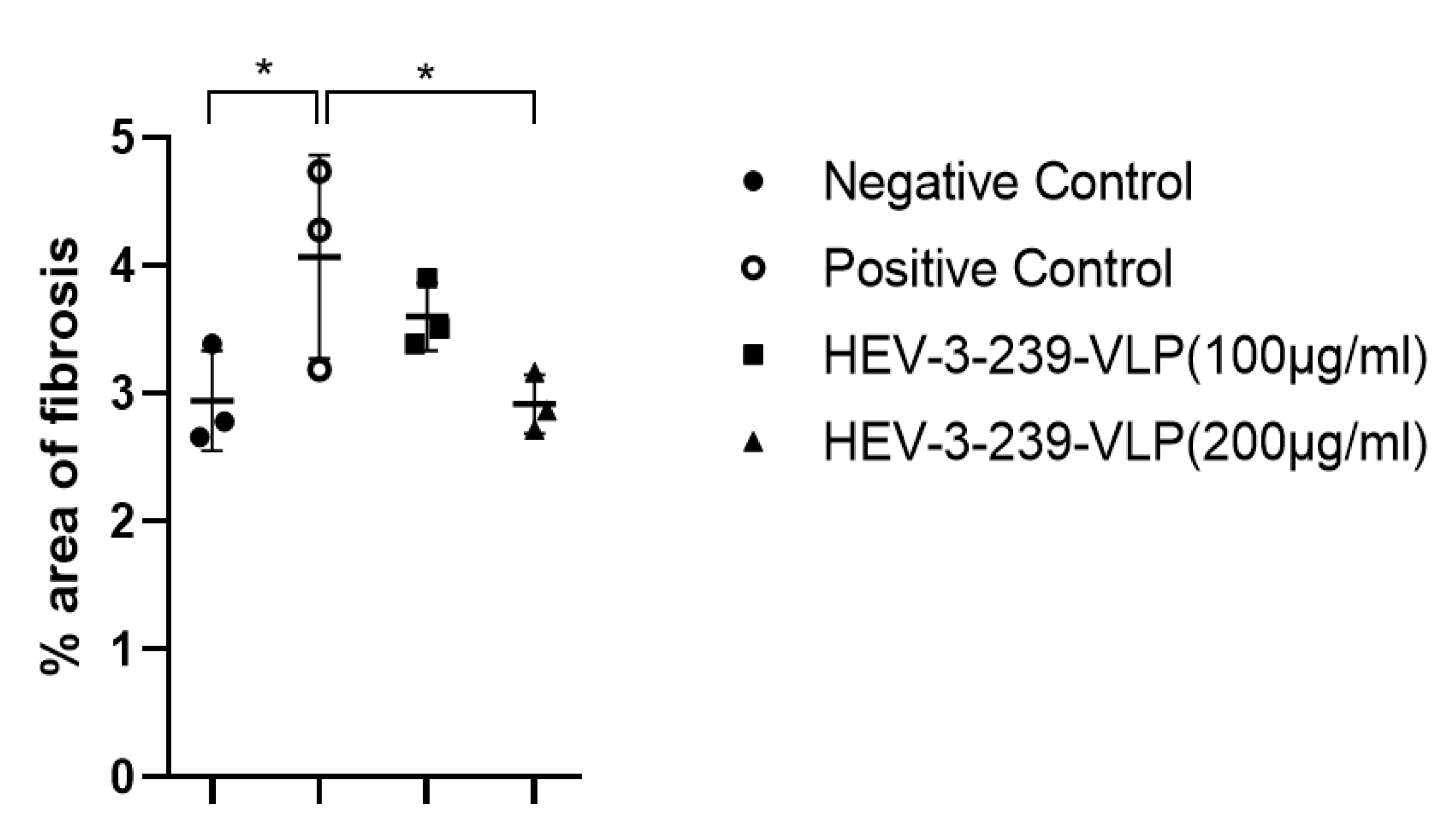
3.5. Histopathological Analysis Indicates the Inhibition of Fibrosis in the Livers of Pigs Vaccinated with VLPs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Smith, D.B.; Simmonds, P.; Jameel, S.; Emerson, S.U.; Harrison, T.J.; Meng, X.-J.; Okamoto, H.; Van Der Poel, W.H.M.; Purdy, M.A.; Members of the International Committee on the Taxonomy of Viruses Study Group. Consensus proposals for classification of the family Hepeviridae. J. Gen. Virol. 2014, 95, 2223–2232. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.B.; Simmonds, P.; Izopet, J.; Oliveira-Filho, E.; Ulrich, R.G.; Johne, R.; Koenig, M.; Jameel, S.; Harrison, T.J.; Meng, X.-J.; et al. Proposed reference sequences for hepatitis E virus subtypes. J. Gen. Virol. 2016, 97, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Rein, D.B.; Stevens, G.A.; Theaker, J.; Wittenborn, J.S.; Wiersma, S.T. The global burden of hepatitis E virus genotypes 1 and 2 in 2005. Hepatology 2012, 55, 988–997. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Li, C.; Hagedorn, C.H. Phylogenetic analysis of global hepatitis E virus sequences: Genetic diversity, subtypes and zoonosis. Rev. Med. Virol. 2006, 16, 5–36. [Google Scholar] [CrossRef]
- Colson, P.; Borentain, P.; Queyriaux, B.; Kaba, M.; Moal, V.; Gallian, P.; Heyries, L.; Raoult, D.; Gerolami, R. Pig Liver Sausage as a Source of Hepatitis E Virus Transmission to Humans. J. Infect. Dis. 2010, 202, 825–834. [Google Scholar] [CrossRef] [Green Version]
- Kamar, N.; Selves, J.; Mansuy, J.-M.; Ouezzani, L.; Péron, J.-M.; Guitard, J.; Cointault, O.; Esposito, L.; Abravanel, F.; Danjoux, M.; et al. Hepatitis E Virus and Chronic Hepatitis in Organ-Transplant Recipients. N. Engl. J. Med. 2008, 358, 811–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, I.; Holla, R.P.; Jameel, S. Molecular virology of hepatitis E virus. Virus Res. 2011, 161, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Jameel, S.; Zafrullah, M.; Ozdener, M.H.; Panda, S.K. Expression in animal cells and characterization of the hepatitis E virus structural proteins. J. Virol. 1996, 70, 207–216. [Google Scholar] [CrossRef] [Green Version]
- Kannan, H.; Fan, S.; Patel, D.; Bossis, I.; Zhang, Y.-J. The Hepatitis E Virus Open Reading Frame 3 Product Interacts with Microtubules and Interferes with Their Dynamics. J. Virol. 2009, 83, 6375–6382. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.-H.; Purcell, R.H.; Emerson, S.U. A truncated ORF2 protein contains the most immunogenic site on ORF2: Antibody responses to non-vaccine sequences following challenge of vaccinated and non-vaccinated macaques with hepatitis E virus. Vaccine 2005, 23, 3157–3165. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, C.-B.; Li, R.-C.; Li, Y.-M.; Zheng, Y.-J.; Li, Y.-P.; Luo, D.; Pan, B.-B.; Nong, Y.; Ge, S.-X.; et al. Randomized-controlled phase II clinical trial of a bacterially expressed recombinant hepatitis E vaccine. Vaccine 2009, 27, 1869–1874. [Google Scholar] [CrossRef]
- Shrestha, M.P.; Scott, R.M.; Joshi, D.M.; Mammen, M.P.; Thapa, G.B.; Thapa, N.; Myint, K.S.A.; Fourneau, M.; Kuschner, R.A.; Shrestha, S.K.; et al. Safety and Efficacy of a Recombinant Hepatitis E Vaccine. N. Engl. J. Med. 2007, 356, 895–903. [Google Scholar] [CrossRef]
- Cao, Y.-F.; Tao, H.; Hu, Y.-M.; Shi, C.-B.; Wu, X.; Liang, Q.; Chi, C.-P.; Li, L.; Liang, Z.-L.; Meng, J.-H.; et al. A phase 1 randomized open-label clinical study to evaluate the safety and tolerability of a novel recombinant hepatitis E vaccine. Vaccine 2017, 35, 5073–5080. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Jiang, J.; Zhang, X.; Wang, N.; Wang, K.; Li, Q.; Li, T.; Lin, Q.; Wang, Y.; Yu, H.; et al. Characterization of capsid protein (p495) of hepatitis E virus expressed in Escherichia coli and assembling into particles in vitro. Vaccine 2018, 36, 2104–2111. [Google Scholar] [CrossRef] [PubMed]
- Li, S.W.; Zhang, J.; Li, Y.M.; Ou, S.H.; Huang, G.Y.; He, Z.Q.; Ge, S.X.; Xian, Y.L.; Pang, S.Q.; Ng, M.H.; et al. A bacterially expressed particulate hepatitis E vaccine: Antigenicity, immunogenicity and protectivity on primates. Vaccine 2005, 23, 2893–2901. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.-C.; Zhang, J.; Zhang, X.-F.; Zhou, C.; Wang, Z.-Z.; Huang, S.-J.; Wang, H.; Yang, C.-L.; Jiang, H.-M.; Cai, J.-P.; et al. Efficacy and safety of a recombinant hepatitis E vaccine in healthy adults: A large-scale, randomised, double-blind placebo-controlled, phase 3 trial. Lancet 2010, 376, 895–902. [Google Scholar] [CrossRef]
- Wu, T.; Li, S.-W.; Zhang, J.; Ng, M.-H.; Xia, N.-S.; Zhao, Q. Hepatitis E vaccine development. Hum. Vaccines Immunother. 2012, 8, 823–827. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, X.-F.; Huang, S.-J.; Wu, T.; Hu, Y.-M.; Wang, Z.-Z.; Wang, H.; Jiang, H.-M.; Wang, Y.-J.; Yan, Q.; et al. Long-Term Efficacy of a Hepatitis E Vaccine. N. Engl. J. Med. 2015, 372, 914–922. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Wu, X.; Li, L.; Liu, Z.; Wang, Z. Use of baculovirus expression system for generation of virus-like particles: Successes and challenges. Protein Expr. Purif. 2013, 90, 104–116. [Google Scholar] [CrossRef]
- Li, T.C.; Yamakawa, Y.; Suzuki, K.; Tatsumi, M.; Razak, M.A.; Uchida, T.; Takeda, N.; Miyamura, T. Expression and self-assembly of empty virus-like particles of hepatitis E virus. J. Virol. 1997, 71, 7207–7213. [Google Scholar] [CrossRef] [Green Version]
- Li, T.-C.; Suzaki, Y.; Ami, Y.; Dhole, T.N.; Miyamura, T.; Takeda, N. Protection of cynomolgus monkeys against HEV infection by oral administration of recombinant hepatitis E virus-like particles. Vaccine 2004, 22, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Park, B.-J.; Ahn, H.-S.; Han, S.-H.; Go, H.-J.; Lee, J.-B.; Park, S.-Y.; Song, C.-S.; Lee, S.-W.; Paik, H.-J.; Choi, Y.-K.; et al. Evaluation of the protective effects of a nanogel-based vaccine against rabbit hepatitis E virus. Vaccine 2019, 37, 5972–5978. [Google Scholar] [CrossRef]
- Mizuo, H.; Suzuki, K.; Takikawa, Y.; Sugai, Y.; Tokita, H.; Akahane, Y.; Itoh, K.; Gotanda, Y.; Takahashi, M.; Nishizawa, T.; et al. Polyphyletic Strains of Hepatitis E Virus Are Responsible for Sporadic Cases of Acute Hepatitis in Japan. J. Clin. Microbiol. 2002, 40, 3209–3218. [Google Scholar] [CrossRef] [Green Version]
- Blasco-Perrin, H.; Abravanel, F.; Blasco-Baque, V.; Péron, J.M. Hepatitis E, the neglected one. Liver Int. 2016, 36 (Suppl. 1), 130–134. [Google Scholar] [CrossRef]
- Pérez-Gracia, M.T.; Lindemann, M.L.M.; Villalba, M.C.M. Hepatitis E: Current status. Rev. Med. Virol. 2013, 23, 384–398. [Google Scholar] [CrossRef]
- Kamar, N.; Bendall, R.; Abravanel, F.; Xia, N.; Ijaz, S.; Izopet, J.; Dalton, H.R. Hepatitis E. Lancet 2012, 379, 2477–2488. [Google Scholar] [CrossRef]
- Kumar, A.; Beniwal, B.; Kar, P.; Sharma, J.B.; Murthy, N.S. Hepatitis E. in Pregnancy. Obstet. Gynecol. Surv. 2005, 60, 7–8. [Google Scholar] [CrossRef]
- Khuroo, M.S.; Kamili, S. Aetiology, clinical course and outcome of sporadic acute viral hepatitis in pregnancy. J. Viral Hepat. 2003, 10, 61–69. [Google Scholar] [CrossRef]
- Dalton, H.R.; Kamar, N.; Izopet, J. Hepatitis E in developed countries: Current status and future perspectives. Futur. Microbiol. 2014, 9, 1361–1372. [Google Scholar] [CrossRef]
- Sinclair, S.M.; Jones, J.K.; Miller, R.K.; Greene, M.F.; Kwo, P.Y.; Maddrey, W.C. The Ribavirin Pregnancy Registry: An Interim Analysis of Potential Teratogenicity at the Mid-Point of Enrollment. Drug Saf. 2017, 40, 1205–1218. [Google Scholar] [CrossRef]
- Zhao, Q.; Zhang, J.; Wu, T.; Li, S.-W.; Ng, M.-H.; Xia, N.-S.; Shih, J.W.-K. Antigenic determinants of hepatitis E virus and vaccine-induced immunogenicity and efficacy. J. Gastroenterol. 2013, 48, 159–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, X.; Wang, S.; Dai, X.; Shi, C.; Wen, Y.; Zhu, M.; Zhan, S.; Meng, J. Rabbit as a Novel Animal Model for Hepatitis E Virus Infection and Vaccine Evaluation. PLoS ONE 2012, 7, e51616. [Google Scholar] [CrossRef] [PubMed]
- Tsareva, T.S. Recombinant vaccine against hepatitis E: Dose response and protection against heterologous challenge. Vaccine 1997, 15, 1834–1838. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Pigs HEV RNA Detected | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Group | Weeks Post-Inoculation | |||||||||||
| Sample | 0 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |
| Negative Control | Serum | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 |
| Feces | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | |
| Positive Control | Serum | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 1/3 | 2/3 | 1/3 | 2/3 | 0/3 |
| Feces | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 2/3 | 2/3 | 3/3 | 2/3 | 1/3 | |
| HEV-3-239-VLP (100 μg/mL) | Serum | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 2/3 | 0/3 | 0/3 | 0/3 |
| Feces | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 2/3 | 1/3 | 0/3 | 0/3 | |
| HEV-3-239-VLP (200 μg/mL) | Serum | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 |
| Feces | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Go, H.-J.; Park, B.-J.; Ahn, H.-S.; Kim, D.-H.; Kim, D.-Y.; Kim, J.-H.; Lee, J.-B.; Park, S.-Y.; Song, C.-S.; Lee, S.-W.; et al. Pigs Immunized with the Virus-like Particle Vaccine Are Protected against the Hepatitis E-3 Virus. Vaccines 2021, 9, 1265. https://doi.org/10.3390/vaccines9111265
Go H-J, Park B-J, Ahn H-S, Kim D-H, Kim D-Y, Kim J-H, Lee J-B, Park S-Y, Song C-S, Lee S-W, et al. Pigs Immunized with the Virus-like Particle Vaccine Are Protected against the Hepatitis E-3 Virus. Vaccines. 2021; 9(11):1265. https://doi.org/10.3390/vaccines9111265
Chicago/Turabian StyleGo, Hyeon-Jeong, Byung-Joo Park, Hee-Seop Ahn, Dong-Hwi Kim, Da-Yoon Kim, Jae-Hyeong Kim, Joong-Bok Lee, Seung-Yong Park, Chang-Seon Song, Sang-Won Lee, and et al. 2021. "Pigs Immunized with the Virus-like Particle Vaccine Are Protected against the Hepatitis E-3 Virus" Vaccines 9, no. 11: 1265. https://doi.org/10.3390/vaccines9111265
APA StyleGo, H.-J., Park, B.-J., Ahn, H.-S., Kim, D.-H., Kim, D.-Y., Kim, J.-H., Lee, J.-B., Park, S.-Y., Song, C.-S., Lee, S.-W., Choi, Y.-K., & Choi, I.-S. (2021). Pigs Immunized with the Virus-like Particle Vaccine Are Protected against the Hepatitis E-3 Virus. Vaccines, 9(11), 1265. https://doi.org/10.3390/vaccines9111265