Towards Improved Use of Vaccination in the Control of Infectious Bronchitis and Newcastle Disease in Poultry: Understanding the Immunological Mechanisms

 , , ,
, , ,  ,
,  , and
, and
Abstract
1. Infectious Bronchitis (IB) and Newcastle Diseases (ND) in Poultry
1.1. Introduction
1.2. Public Health Importance of IB and ND in Poultry
1.3. Economic Importance of IB and ND in Poultry
2. Immunity and Immune Response against IB and ND in Poultry
2.1. Immune System in Birds Relative to Viral Diseases
2.2. Host Immune Responses to IB and ND
2.2.1. Host Immune Responses to IBV
2.2.2. Host Immune Responses to NDV
2.3. Vaccine-Mediated Immunity against IB and ND
2.3.1. Vaccine-Mediated Immunity against IBV
2.3.2. Vaccine-Mediated Immunity against NDV
2.4. Immunopathology in the Hosts Resulting from IBV and NDV
2.4.1. Immunopathology in the Hosts from IBV
2.4.2. Immunopathology in the Hosts from NDV
3. Vaccination against IB and ND
3.1. Available Vaccines against IB and ND
3.1.1. Available Vaccines against Infectious Bronchitis
3.1.2. Available Vaccines against Newcastle Disease (ND)
3.2. Recent Advances in Vaccine Development against IB and ND
3.2.1. Recent Advances in IB Vaccine Development
3.2.2. Recent Advances in ND Vaccine Development
3.3. IB and ND Vaccine Development
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bande, F.; Arshad, S.S.; Omar, A.R.; Bejo, M.H.; Abubakar, M.S.; Abba, Y. Pathogenesis and diagnostic approaches of avian infectious bronchitis. Adv. Virol. 2016, 2016, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Dimitrov, K.M.; Ferreira, H.L.; Pantin-Jackwood, M.J.; Taylor, T.L.; Goraichuk, I.V.; Crossley, B.M.; Killian, M.L.; Bergeson, N.H.; Torchetti, M.K.; Afonso, C.L.; et al. Pathogenicity and transmission of virulent Newcastle disease virus from 2018–2019 California outbreak and related viruses in young and adult chickens. Virology 2019, 531, 203–218. [Google Scholar] [CrossRef] [PubMed]
- Duffy, S.; Shacketton, L.A.; Holmes, E.C. Rates of evolutionary change in viruses: Patterns and determinants. Nat. Rev. Genet. 2008, 9, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Miller, P.J.; Decanini, E.L.; Afonso, C.L. Newcastle disease: Evolution of genotypes and the related diagnostic challenges. Infect. Genet. Evol. 2000, 10, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Moreno, A.; Franzo, G.; Massi, P.; Tosi, G.; Blanco, A.; Antilles, N.; Biarnes, M.; Majó, N.; Nofravías, M.; Dolz, R.; et al. A novel variant of the infectious bronchitis virus resulting from recombination events in Italy and Spain. Avian Pathol. 2017, 46, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Crinion, R.A.; Hofstad, M.S. Pathogenicity of four serotypes of avian infectious bronchitis virus for the oviduct of young chickens of various ages. Avian Dis. 1972, 16, 351–363. [Google Scholar] [CrossRef]
- Müller, S.; Schubert, A.; Zajac, J.; Dyck, T.; Oelkrug, C. IgY antibodies in human nutrition for disease prevention. Nutr. J. 2015, 14, 109. [Google Scholar] [CrossRef]
- Tan, L.; Zhang, Y.; Liu, F.; Yuan, Y.; Zhan, Y.; Sun, Y.; Qui, X.; Meng, C.; Song, C.; Ding, C. Infectious bronchitis virus poly-epitope-based vaccine protects chickens from acute infection. Vaccine 2016, 34, 5209–5216. [Google Scholar] [CrossRef]
- Miller, P.J.; Kim, L.M.; Ip, H.S.; Afonso, C.L. Evolutionary dynamics of Newcastle disease virus. Virology 2009, 391, 64–72. [Google Scholar] [CrossRef]
- Lin, M.Y.; Liu, H.J.; Ke, G.M. Genetic and antigenic analysis of Newcastle disease viruses from recent outbreaks in Taiwan. Avian Pathol. 2003, 32, 345–350. [Google Scholar] [CrossRef]
- Miller, P.J.; King, D.J.; Afonso, C.L.; Suarez, D.L. Antigenic differences among Newcastle disease virus strains of different genotypes used in vaccine formulation affect viral shedding after a virulent challenge. Vaccine 2007, 25, 7238–7246. [Google Scholar] [CrossRef] [PubMed]
- Emecheta, A.O.; Ike, A.C.; Onu, C.J.; Eze, C.D.; Olovo, C.V. The benefits of supplementation with antibiotic alternatives on Newcastle disease virus titre in poultry. World’s Poult. Sci. J. 2018, 74, 665–673. [Google Scholar] [CrossRef]
- Abdisa, T.; Tagesu, T. Review on Newcastle disease of poultry and its public health importance. J. Vet. Sci. Technol. 2017, 8, 441. [Google Scholar] [CrossRef]
- Keeney, A.H.; Hunter, M.C. Human infection with the Newcastle virus of fowls. AMA Arch. Ophthalmol. 1950, 44, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Schirrmacher, V.; Ahlert, T.; Pröbstle, T.; Steiner, H.H.; Herold-Mende, C.; Gerhards, R.; Hagmüller, E.; Steiner, H.H. Immunization with virus-modified tumor cells. Semin. Oncol. 1998, 25, 677–696. [Google Scholar] [PubMed]
- Moscoso, H.; Raybon, E.O.; Thayer, S.G.; Hofacre, C.L. Molecular detection and serotyping of infectious bronchitis virus from FTA (R) filter paper. Avian Dis. 2005, 49, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Miller, P.J.; Estevez, C.; Yu, Q. Comparison of viral shedding following vaccination with inactivated and live Newcastle disease vaccines formulated with wild-type and recombinant viruses. Avian Dis. 2009, 53, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Ignjatovic, J.; Sapats, S. Avian infectious bronchitis virus. Rev. Sci. Tech. Off. Int. Epizoot. 2000, 19, 493–508. [Google Scholar] [CrossRef] [PubMed]
- Melkamu, S.; Asrat, M. Review on economic importance and current diagnostic techniques of Newcastle disease in poultry. Int. J. Adv. Res. Biol. Sci. 2018, 5, 117–125. [Google Scholar]
- Thomazelli, L.M.; Araujo, J.D.; Ferreira, C.S.; Hurtado, R.; Oliveira, D.B. Molecular surveillance of the Newcastle disease virus in domestic and wild birds on the North-Eastern coast and Amazon biome of Brazil. Braz. J. Poult. Sci. 2012, 14, 1–7. [Google Scholar] [CrossRef]
- Jackwood, M.W.; de Wit, S. Infectious bronchitis. In Diseases of Poultry, 13th ed.; Swayne, D.E., Glisson, J.R., McDougald, L.R., Nolan, L.K., Suarez, D.L., Neil, V., Eds.; John Wiley and Sons, Inc.: Hoboken, NJ, USA, 2013; pp. 139–159. [Google Scholar]
- Khataby, K.; Fellahi, S.; Loutfib, C.; Mustapha, E.M. Avian infectious bronchitis virus in Africa: A review. Vet. Quart. 2016, 36, 71–75. [Google Scholar] [CrossRef] [PubMed]
- McMartin, D.A. Infectious bronchitis. In Virus Infections of Birds; McFerran, J.B., McNulty, M.S., Eds.; Elsevier Science Publishers: Amsterdam, The Netherland, 1993; pp. 249–274. [Google Scholar]
- Assayag, M.S.; Chacón, J.L.; Rocha, P.T.; Kuana, S. Economic impact of infectious bronchitis in a Brazilian poultry integration system. In Proceedings of the 7th Symposium on Avian Corona and Pneumoviruses and Complicating Pathogens, Rauischholzhausen, Germany, 18–21 June 2012; pp. 80–83. [Google Scholar]
- Colvero, L.P.; Villarreal, L.Y.B.; Torres, C.A.; Brandão, P.E. Assessing the economic burden of avian infectious bronchitis on poultry farms in Brazil. Rev. Sci. Tech. Off. Int. Epizoot. 2015, 34, 993–999. [Google Scholar] [CrossRef] [PubMed]
- Dhama, K.; Singh, S.D.; Barathidasan, R.; Desingu, P.A.; Chakraborty, S.; Tiwari, R.; Kumar, M.A. Emergence of avian infectious bronchitis virus and its variants need better diagnosis, prevention and control strategies: A global perspective. Pak. J. Biol. Sci. 2014, 17, 751–767. [Google Scholar] [CrossRef] [PubMed]
- Mengesha, M.; Tamir, B.; Dessie, T. Village chicken constraint and traditional management practices in Jamma district, South Wollo, Ethiopia. Livest. Res. Rural Dev. 2011, 23, 37. [Google Scholar]
- Khatun, M.; Islam, I.; Ershaduzzaman, M.; Islam, H.M.S.; Yasmin, S.; Hossen, A.; Hasan, M. Economic Impact of Newcastle Disease on Village Chickens—A Case of Bangladesh. J. Econ. Bus. 2018, 1, 358–367. [Google Scholar]
- Antipas, B.B.; Bidjeh, K.; Youssouf, M.L. Epidemiology of Newcastle disease and its economic impact in Chad. Eur. J. Exp. Biol. 2012, 2, 2286–2292. [Google Scholar]
- Sadiq, M.B.; Mohammed, B.R. The economic impact of some important viral diseases affecting the poultry industry in Abuja, Nigeria. Sokoto J. Vet. Sci. 2017, 15, 7–17. [Google Scholar] [CrossRef]
- Kumar, B.G.; Joshi, P.K.; Datta, K.K.; Singh, S.B. An Assessment of economic losses due to avian flu in Manipur state. Agric. Econ. Res. Rev. 2008, 21, 37–47. [Google Scholar]
- Cavanagh, D. Coronavirus avian infectious bronchitis virus. Vet. Res. 2007, 38, 281–297. [Google Scholar] [CrossRef]
- Kapczynski, D.R.; Afonso, C.L.; Miller, P.J. Immune responses of poultry to Newcastle disease virus. Dev. Comp. Immunol. 2013, 41, 447–453. [Google Scholar] [CrossRef]
- Hughes, S.; Poh, T.Y.; Bumstead, N.; Kaiser, P. Re-evaluation of the chicken MIP family of chemokines and their receptors suggests that CCL5 is the prototypic MIP family chemokine, and that different species have developed different repertoires of both the CC chemokines and their receptors. Dev. Comp. Immunol. 2007, 31, 72–86. [Google Scholar] [CrossRef] [PubMed]
- Temperley, N.D.; Berlin, S.; Paton, I.R.; Griffin, D.K.; Burt, D.W. Evolution of the chicken Toll-like receptor gene family: A story of gene gain and gene loss. BMC Genom. 2008, 9, 62. [Google Scholar] [CrossRef] [PubMed]
- Cormican, P.; Lloyd, A.T.; Downing, T.; Connell, S.J.; Bradley, D.; O’Farrelly, C. The avian Toll-Like receptor pathway-subtle differences amidst general conformity. Dev. Comp. Immunol. 2009, 33, 967–973. [Google Scholar] [CrossRef] [PubMed]
- Kasier, P. Advances in avian immunology prospects for disease control: A review. Avian Pathol. 2010, 39, 309–324. [Google Scholar] [CrossRef]
- Joiner, K.S.; Hoerr, F.J.; Ewald, S.J.; van Santen, V.L.; Wright, J.C.; van Ginkel, F.W.; Toro, H. Pathogenesis of infectious bronchitis virus in vaccinated chickens of two majorhistocompatibility B complex genotypes. Avian Dis. 2007, 51, 758–763. [Google Scholar] [CrossRef]
- Caron, L.F. Etiology and immunology of infectious bronchitis virus. Braz. J. Poult. Sci. 2010, 12, 115–119. [Google Scholar] [CrossRef]
- Parra, D.; Takizawa, F.; Sunyer, J.O. Evolution of B cell immunity. Ann. Rev. Anim. Biosci. 2013, 1, 65–97. [Google Scholar] [CrossRef]
- Rose, M.E.; Orlans, E.; Buttress, N. Immunoglobulin classes in the hen’s egg: Their segregation in yolk and white. Eur. J. Immunol. 1974, 4, 521–523. [Google Scholar] [CrossRef]
- Kjærup, R.M.; Dalgaard, T.S.; Norup, L.R.; Hamzic, E.; Sørensen, P.; Juul-Madsen, H.R. Characterization of cellular and humoral immune responses after IBV infection in chicken lines differing in MBL serum concentration. Viral Immunol. 2014, 27, 529–542. [Google Scholar] [CrossRef]
- Júnior, A.F.; dos Santos, J.P.; Sousa, I.O.; Martin, I.; Alves, E.G.L.; Rosado, I.R. Gallus gallus domesticus: Immune system and its potential for generation of immunobiologics. Cienc. Rural 2018, 48, e20180250. [Google Scholar]
- Bienenstock, J.; Johnston, N.; Perey, D.Y. Bronchial lymphoid tissue. I. Morphologic characteristics. Lab. Investig. 1973, 28, 686–692. [Google Scholar]
- Jeurissen, S.H.M.; Vervelde, L.; Janse, E.M. Structure and function of lymphoid tissues of the chicken. Poult. Sci. Rev. 1994, 5, 183–207. [Google Scholar]
- Oshima, K.; Hiramatsu, K. Distribution of T-cell subsets and immunoglobulin-containing cells in nasal-associated lymphoid tissue (NAL T) of chickens. Histol. Histopathol. 2000, 15, 713–720. [Google Scholar]
- Nochi, T.; Jansen, C.A.; Toyomizu, M.; Eden, W. The well-developed mucosal immune systems of birds and mammals allow for similar approaches of mucosal vaccination in both types of animals. Front. Nutr. 2018, 5, 60. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.H.; Göbel, T.W.F.; Kubota, T.; Cooper, M.D. T cell development in the chicken. Poult. Sci. 1994, 73, 1012–1018. [Google Scholar] [CrossRef] [PubMed]
- Kaspers, B.; Göbel, T.W.F. The avian immune system. Encycl. Immunol. 2016, 30, 13–17. [Google Scholar] [CrossRef]
- Ike, A.C.; Onu, C.J.; Ononugbo, C.M.; Reward, E.E.; Muo, S.O. Immune response to herpes simplex virus infection and vaccine development. Vaccines 2020, 8, 302. [Google Scholar] [CrossRef]
- Cong, F.; Liu, X.; Han, Z.; Shao, Y.; Kong, X.; Liu, S. Transcriptome analysis of chicken kidney tissues following coronavirus avian infectious bronchitis virus infection. BMC Genom. 2013, 14, 743. [Google Scholar] [CrossRef]
- He, Y.; Xie, Z.; Dai, J.; Cao, Y.; Hou, J.; Zheng, Y.; Wei, T.; Mo, M.; Wei, P. Responses of the toll-like receptor and melanoma differentiation-associated protein 5 signaling pathways to avian infectious bronchitis virus infection in chicks. Virol. Sin. 2016, 31, 57–68. [Google Scholar] [CrossRef]
- Smith, J.; Sadeyen, J.-R.; Cavanagh, D.; Kaiser, P.; Burt, D.W. The early immune response to infection of chickens with infectious bronchitis virus (IBV) in susceptible and resistant birds. BMC Vet. Res. 2015, 11, 256. [Google Scholar] [CrossRef]
- Zhang, W.-X.; Zuo, E.-W.; He, Y.; Chen, D.-Y.; Long, X.; Chen, M.-J.; Li, T.-T.; Yang, X.-G.; Xu, H.-Y.; Lu, S.-S.; et al. Promoter structures and differential responses to viral and non-viral inducers of chicken melanoma differentiation-associated gene 5. Mol. Immunol. 2016, 76, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Chhabra, R.; Kuchipudi, S.V.; Chantrey, J.; Ganapathy, K. Pathogenicity and tissue tropism of infectious bronchitis virus is associated with elevated apoptosis and innate immune responses. Virology 2016, 48, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Zhang, X.; Wu, T.; Su, J.; Wang, Y.; Wang, Y.; Ruan, B.; Niu, X.; Wu, Y. Avian infectious bronchitis virus disrupts the melanoma differentiation associated gene (MDA5) signaling pathway by cleavage of the adaptor protein MAVS. BMC Vet. Res. 2017, 13, 332. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Li, J.; Liu, H.; Zhang, P.; Chen, D.; Men, S.; Li, X.; Wang, H. Induction of innate immune responses following introduction of infectious bronchitis viruses (IBV) in the trachea and renal tissues of chickens. Microb. Pathog. 2018, 116, 54–61. [Google Scholar] [CrossRef]
- Amarasinghe, A.; Abdul-Cader, M.S.; Almatrouk, Z.; van der Meer, F.; Cork, S.C.; Gomis, S.; Abdul-Careem, M.F. Induction of innate immune responses characterized by production of interleukin (IL)-1β and recruitment of macrophages to the respiratory tract of chickens following infection with infectious bronchitis virus (IBV). Vet. Microbiol. 2018, 215, 1–10. [Google Scholar] [CrossRef]
- Elhamouly, M.; Terade, T.; Nii, T.; Isobe, N.; Yoshimura, Y. Innate antiviral immune response against infectious bronchitis virus and involvement of prostaglandin E2 in the uterine mucosa of laying hens. Theriogenology 2018, 110, 122–129. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, T.; Xu, Q.; Han, Z.; Liang, S.; Shao, Y.; Ma, D.; Liu, S. Differential modulation of avian β-defensin and toll-like receptor expression in chickens infected with infectious bronchitis virus. Appl. Microbiol. Biotechnol. 2015, 99, 9011–9024. [Google Scholar] [CrossRef]
- Gao, P.; Chen, L.; Fan, L.; Ren, J.; Du, H.; Sun, M.; Li, Y.; Xie, P.; Lin, Q.; Liao, M.; et al. Newcastle disease virus RNA-induced IL-1β expression via the NLRP3/caspase-1 inflammasome. Vet. Res. 2020, 51, 53. [Google Scholar] [CrossRef]
- Qian, J.; Xu, X.; Ding, J.; Yin, R.; Sun, Y.; Xue, C.; Ding, C.; Yu, S.; Liu, X.; Hu, S.; et al. Newcastle disease virus-like particles induce DC maturation throughTLR4/NF-κB pathway and facilitate DC migration by CCR7-CCL19/CCL21 axis. J. Vet. Microbiol. 2017, 203, 158–166. [Google Scholar] [CrossRef]
- Seo, S.H.; Collisson, E.W. Specific Cytotoxic T Lymphocytes are involved in In Vivo Clearance of Infectious Bronchitis Virus. J. Virol. 1997, 71, 5173–5177. [Google Scholar] [CrossRef]
- Zhang, W.; Bouwman, K.M.; van Beurden, S.J.; Ordonez, S.R.; van Eijk, M.; Haagsman, H.P.; Verkeije, H.M.; Veldhuizen, E.J.A. Chicken mannose binding lectin has antiviral activity towards infectious bronchitis virus. Virology 2017, 509, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Rue, C.A.; Susta, L.; Cornax, I.; Brown, C.C.; Kapczyriski, D.R.; Suarez, D.L.; King, D.J.; Miller, P.J.; Afonso, C.L. Virulent Newcastle disease virus elicits a strong innate immune response in chickens. J. Gen. Virol. 2011, 92, 931–939. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Chen, Y.; Zhao, W.; Zhang, T.; Liu, C.; Qi, T.; Han, Z.; Shao, Y.; Ma, D.; Liu, S. Infection of Goose with genotype VIId Newcastle disease virus of Goose origin elicits strong immune responses at early stage. Front. Microbiol. 2016, 7, 1587. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, Y.; Takeuchi, O.; Kato, H.; Kumar, H.; Matsui, K.; Morii, E.; Aozasa, K.; Kawai, T.; Akira, S. Alveolar macrophages are the primary interferon-α-producer in pulmonary infection with RNA viruses. Immunity 2007, 27, 240–252. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Ingle, H.; Mishra, S.; Mahla, R.S.; Kumar, A.; Kawai, T.; Akira, S.; Takaoka, A.; Raut, A.A.; Kumar, H. IPS-1 differentially induces TRAIL, BCL2, BIRC3 and PRKCE in type 1 interferons-dependent and independent anticancer activity. Cell Death Dis. 2015, 6, e1758. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Ning, Z.; Sun, M.; Gao, S.; Kang, Y.; Xie, P.; Ren, T. Interferon regulatory factor 7- (IRF7-) mediated immune response affects Newcastle disease virus replication in chicken embryo fibroblasts. Acta Vet. Hung. 2014, 62, 500–511. [Google Scholar] [CrossRef]
- Yang, X.; Arslan, M.; Liu, X.; Song, H.; Du, M.; Li, Y.; Zhang, Z. IFN-γ establishes interferon-stimulated gene-mediated antiviral state against Newcastle disease virus in chicken fibroblasts. Acta Biochim. Biophys. Sin. 2020, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Jiang, L.; Liu, L.; Sun, L.; Zhao, W.; Chen, Y.; Qi, T.; Han, Z.; Shao, Y.; Liu, S.; et al. Induction of avian β-defensin 2 is possibly mediated by the p38 MAPK signal pathway in chicken embryo fibroblasts after Newcastle disease virus infection. Front. Microbiol. 2018, 9, 751. [Google Scholar] [CrossRef]
- Rasoli, M.; Yeap, S.K.; Tan, S.W.; Moeini, H.; Ideris, A.; Bejo, M.H.; Alitheen, N.B.M.; Kaiser, P.; Omar, A.R. Alteration in lymphocyte responses, cytokine and chemokine profiles in chickens infected with genotype VII and VIII velogenic Newcastle disease virus. Comp. Immunol. Microbiol. Infect. Dis. 2013, 37, 11–21. [Google Scholar] [CrossRef]
- Kang, Y.; Li, Y.; Yuan, R.; Feng, M.; Xiang, B.; Sun, M.; Li, Y.; Xie, P.; Tan, Y.; Ren, T. Host innate immune responses of ducks infected with Newcastle disease viruses of different pathogenicities. Front. Microbiol. 2015, 6, 1283. [Google Scholar] [CrossRef]
- Schilling, M.A.; Katani, R.; Memari, S.; Cavanaugh, M.; Buza, J.; Radzio-Basu, J.; Mpenda, F.N.; Deist, M.S.; Lamont, S.J.; Kapur, V. Transcriptional innate immune response of the developing chicken embryo to Newcastle disease virus infection. Front. Genet. 2018, 9, 61. [Google Scholar] [CrossRef] [PubMed]
- Ecco, R.; Brown, C.; Susta, L.; Cagle, C.; Cornax, I.; Pantin-Jackwood, M.; Miller, P.J.; Afonso, C.L. In vivo transcriptional cytokine responses and association with clinical and pathological outcomes in chickens infected with different Newcastle disease virus isolates using formalin-fixed paraffin-embedded samples. Vet. Immunol. Immunopathol. 2011, 141, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Deist, M.S.; Gallardo, R.A.; Bunn, D.A.; Kelly, T.R.; Dekkers, J.C.M.; Zhou, H.; Lamont, S.J. Novel mechanisms revealed in the trachea transcriptome of resistant and susceptible chicken lines following infection with Newcastle disease virus. Clin. Vaccine Immunol. 2017, 24, e00027-17. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Sun, Y.; Chen, H.; Dai, Y.; Zhan, Y.; Yu, S.; Qiu, X.; Tan, L.; Song, C.; Ding, C. Activation of the PKR/elF2α signaling cascade inhibits replication of Newcastle disease virus. Virol. J. 2014, 11, 62. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.; Bharadwaj, M.S.K.; Gupta, A.; Kumar, R.; Kumar, S. Chicken viperin inhibits Newcastle disease virus infection in vitro: A possible interaction with the viral matrix protein. Cytokine 2019, 120, 28–40. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Han, Z.; Qi, T.; Zhao, R.; Liu, S. Chicken galectin-1B inhibits Newcastle disease virus adsorption and replication through binding to hemagglutinin-neuraminidase (HN) glycoprotein. J. Biol. Chem. 2017, 292, 20141–20161. [Google Scholar] [CrossRef]
- Xiang, X.; Zhu, W.; Li, Y.; Gao, P.; Liang, J.; Liu, D.; Ding, C.; Liao, M.; Kang, Y.; Ren, T. Immune responses of mature chicken bone marrow derived dendritic cells infected with Newcastle disease virus strains with differing pathogenicity. Arch. Virol. 2018, 163, 1407–1417. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, W.; Xu, H.; Liu, J.; Deng, Y.; Cheng, H.; Zhan, T.; Lu, X.; Liao, T.; Guo, L.; et al. Gga-miR-19b-3p Inhibits Newcastle Disease Virus Replication by Suppressing Inflammatory Response via Targeting RNF11 and ZMYND11. Front. Microbiol. 2019, 10, 2006. [Google Scholar] [CrossRef]
- Okino, C.H.; Alessi, A.C.; Montassier, M.D.-F.S.; Rosa, A.J.D.-M.; Wang, X.; Montassier, H.J. Humoral and cell-mediated immune responses to different doses of attenuated vaccine against avian infectious bronchitis virus. Viral Immunol. 2013, 26, 259–267. [Google Scholar] [CrossRef]
- Chhabra, R.; Forrester, A.; Lemlere, S.; Awad, F.; Chantrey, J.; Ganapathy, K. Mucosal, cellular and humoral immune responses induced by different live infectious bronchitis virus vaccination regimes and protection conferred against infectious bronchitis virus Q1 strain. Clin. Vaccine Immunol. 2015, 22, 1050–1059. [Google Scholar] [CrossRef]
- Fan, W.-S.; Li, H.-M.; He, Y.-N.; Tang, N.; Zhang, L.-H.; Wang, H.-Y.; Zhong, L.; Chen, J.-C.; Wei, T.-C.; Huang, T.; et al. Immune protection conferred by three commonly used commercial live attenuated vaccines against the prevalent local strains of avian infectious bronchitis virus in southern China. J. Vet. Med. Sci. 2018, 80, 1438–1444. [Google Scholar] [CrossRef] [PubMed]
- Orr-Burks, N.; Gulley, S.L.; Gallardo, R.A.; Toro, H.; van Ginkel, F.W. Immunoglobulin A as an early humoral responder after mucosal avian coronavirus vaccination. Avian Dis. 2014, 58, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Awad, F.; Hutton, S.; Forrester, A.; Baylis, M.; Ganapathy, K. Heterologous live infectious bronchitis virus vaccination in day-old commercial broiler chicks: Clinical signs, ciliary health, immune responses and protection against variant infectious bronchitis viruses. Avian Pathol. 2016, 45, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Wang, H.; Yang, T.; Wang, X.; Lu, D.; Li, Y.; Zhang, Y. Priming with a DNA vaccine and boosting with an inactivated vaccine enhance the immune response against infectious bronchitis virus. J. Virol. Methods 2010, 167, 84–89. [Google Scholar] [CrossRef]
- Lopes, P.D.; Okino, C.H.; Fernando, F.S.; Pavani, C.; Casagrande, V.M.; Lopez, R.F.V.; Montassier, M.D.-F.S.; Montassier, H.J. Inactivated infectious bronchitis virus vaccine encapsulated in chitosan nanoparticles induces mucosal immune responses and effective protection against challenge. Vaccine 2018, 36, 2630–2636. [Google Scholar] [CrossRef]
- Santos, R.M.; Fernando, F.S.; Montassier, M.D.-F.S.; Silva, K.-R.; Lopes, P.D.; Pavani, C.; Borzi, M.M.; Okino, C.H.; Montassier, H.J. Memory immune responses and protection of chickens against a nephropathogenic infectious bronchitis virus strain by combining live heterologous and inactivated homologous vaccines. J. Vet. Med. Sci. 2019, 81, 612–619. [Google Scholar] [CrossRef]
- van Ginkel, F.W.; Padgett, J.; Martinez-Romero, G.; Miller, M.S.; Joiner, K.S.; Gulley, S.L. Age dependent immune responses and immune protection after avian coronavirus vaccination. Vaccine 2015, 33, 2655–2661. [Google Scholar] [CrossRef]
- Saiada, F.; Eldemery, F.; Zegpi, R.A.; Gulley, S.L.; Mishra, A.; van Santen, V.L.; Toro, H. Early vaccination of chickens induces suboptimal immunity against infectious bronchitis virus. Avian Dis. 2018, 63, 38–47. [Google Scholar] [CrossRef]
- Zegpi, R.A.; Breedlove, C.; Gulley, S.; Toro, H. Infectious Bronchitis Virus Immune Responses in the Harderian Gland upon Initial Vaccination. Avian Dis. 2019, 64, 92–95. [Google Scholar] [CrossRef]
- Larsen, F.T.; Bed’Hom, B.; Naghizadeh, M.; Kjaerup, R.B.; Zohari, S.; Dalgaard, T.S. Immunoprofiling of peripheral blood from infectious bronchitis virus vaccinated MHC-B chicken lines-monocyte MHC-II expression as a potential correlate of protection. Dev. Comp. Immunol. 2019, 96, 93–102. [Google Scholar] [CrossRef]
- Tian, L.; Wang, H.-N.; Lu, D.; Zhang, Y.-F.; Wang, T.; Kang, R.-M. The immunoreactivity of a chimeric multi-epitope DNA vaccine against IBV in chickens. Biol. Biophys. Res. Comm. 2008, 377, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Liu, T.; Du, Y.; Cui, X.; Yu, Q.; Wang, Z.; Zhang, J.; Li, Y.; Zhu, Q. A novel genotype VII Newcastle disease virus vaccine candidate generated by mutation of the L and F genes confers improved protection in chickens. Vet. Microbiol. 2018, 216, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Brujeni, G.N.; Hassanzadeh, M.; Al-karagoly, H.; Tolouei, T.; Esmailnejad, A. Evaluation of humoral immune responses to enterotropic lentogenic VG/GA vaccine of Newcastle disease in commercial turkey poults (Meleagris gallopavo). Acta Vet. Scan. 2019, 61, 41. [Google Scholar] [CrossRef] [PubMed]
- Shafiei, B.H.; Mosleh, N.; Dudras, H. Assessment of type I interferons, clinical signs and virus shedding in broiler chickens with pre and post challenge Newcastle disease vaccination. Iran. J. Vet. Res. 2020, 21, 84–91. [Google Scholar]
- Sedeik, M.E.; Elbestawy, A.R.; EL-Shall, N.A.; Abd EL-Hack, M.E.; Saadeldin, I.M.; Swelum, A.A. Comparative efficacy of commercial inactivated Newcastle disease virus vaccines against Newcastle disease virus genotype VII in broiler chickens. Poult. Sci. 2019, 98, 2000–2007. [Google Scholar] [CrossRef] [PubMed]
- Rauw, F.; Nguyen, T.G.; Ngabirano, E.; Marche, S.; Lambrecht, B. Specific antibody-mediated immunity in the reproductive tract of laying chickens immunized against Newcastle disease with conventional attenuated and inactivated vaccines. Avian Pathol. 2017, 46, 434–441. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Han, J.; Zhang, Y.; Wei, L.; Yu, S.; Wang, X.; Jin, Z.; Wang, Y.-F. Enhancing mucosal immune response of Newcastle disease virus DNA vaccine using N-2-hydroxypropyl trimethylammonium chloride chitosan and N,O-carboxymethyl chitosan nanoparticles as delivery carrier. Mol. Pharm. 2018, 15, 226–237. [Google Scholar] [CrossRef]
- Zhao, K.; Rong, G.; Hao, Y.; Yu, L.; Kang, H.; Wang, X.; Wang, X.; Jin, Z.; Ren, Z.; Li, Z. IgA response and protection following nasal vaccination of chickens with Newcastle disease virus DNA vaccine nanoencapsulated with Ag@SiO2 hollow nanoparticles. Sci. Rep. 2016, 6, 25720. [Google Scholar] [CrossRef]
- Jin, Z.; Li, D.; Dai, C.; Cheng, G.; Wang, X.; Zhao, K. Response of live Newcastle disease virus encapsulated in N-2-hydroxypropyl dimethylethyl ammonium chloride chitosan nanoparticles. Carbohydr. Polym. 2017, 171, 267–280. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Li, S.; Li, W.; Yu, L.; Duan, X.; Han, J.; Wang, X.; Jin, Z. Quaternized chitosan nanoparticles loaded with the combined attenuated live vaccine against Newcastle disease and infectious bronchitis elicit immune response in chicken after intranasal administration. Drug Deliv. 2017, 24, 1574–1586. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Zhai, X.; Lai, Y.; Zuo, L.; Zhang, Y.; Mei, X.; Xiang, R.; Kang, Z.; Zhou, L.; Wang, H. Construction and immunogenicity of novel chimeric virus-like particles bearing antigens of infectious bronchitis virus and Newcastle disease virus. Viruses 2019, 11, 254. [Google Scholar] [CrossRef] [PubMed]
- Benyeda, Z.S.; Szeredi, L.; Mato, T.; Suveges, T.; Balka, G.Y.; Abonyi-Toth, Z.S.; Rusvai, M.; Palya, V. Comparative histopathology and immunohistochemistry of QX-like, Massachusetts and 793/B serotypes of infectious bronchitis virus infection in chickens. J. Comp. Path. 2010, 143, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Bezuidenhout, A.; Mondal, S.P.; Buckles, E.L. Histopathological and immunohistochemical study of air sac lesions induced by two strains of infectious bronchitis virus. J. Comp. Path. 2011, 145, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Jackwood, M.W.; Jordan, B.J.; Roh, H.-J.; Hilt, D.A.; Williams, S.M. Evaluating protection against infectious bronchitis virus by clinical signs, ciliostasis, challenge virus detection, and histopathology. Avian Dis. 2015, 59, 368–374. [Google Scholar] [CrossRef]
- Liang, J.Q.; Fang, S.; Yuan, Q.; Huang, M.; Chen, R.A.; Fung, T.S.; Liu, D.X. N-linked glycosylation of the membrane protein ectodomain regulates infectious bronchitis virus-induced ER stress response, apoptosis and pathogenesis. Virology 2019, 531, 48–56. [Google Scholar] [CrossRef]
- Nii, T.; Isobe, N.; Yoshimura, Y. Effects of avian infectious bronchitis virus antigen on egg shell formation and immunoreaction in hen oviduct. Theriogenology 2014, 81, 1129–1138. [Google Scholar] [CrossRef]
- Okino, C.H.; Santos, I.L.; Fernando, F.S.; Alessi, A.C.; Wang, X.; Montassier, H.J. Inflammatory and cell-mediated immune responses in the respiratory tract of chickens to infection with avian infectious bronchitis virus. Viral Immunol. 2014, 27, 1–9. [Google Scholar] [CrossRef]
- Han, X.; Tian, Y.; Guan, R.; Gao, W.; Yang, X.; Zhou, L.; Wang, H. Infectious bronchitis virus infection induces apoptosis during replication in chicken macrophage HD11 cells. Viruses 2017, 9, 198. [Google Scholar] [CrossRef]
- Liao, Y.; Fung, T.S.; Huang, M.; Fang, S.G.; Zhong, Y.; Liu, D.X. Upregulation of CHOP/GADD153 during coronavirus infectious bronchitis virus infection modulates apoptosis by restricting activation of the extracellular signal-regulated kinase pathway. J. Virol. 2013, 87, 8124–8134. [Google Scholar] [CrossRef]
- Liao, Y.; Wang, X.; Huang, M.; Tam, J.P.; Liu, D.X. Regulation of the p38 mitogen-activated protein kinase and dual-specificity phosphatases 1 feed-back loop modulates the induction of interleukin 6 and 8 in cells infected with coronavirus infectious bronchitis virus. Virology 2011, 420, 106–116. [Google Scholar] [CrossRef]
- Fung, T.S.; Liu, D.X. Activation of the c-Jun NH2-terminal kinase pathway by coronavirus infectious bronchitis virus promotes apoptosis independently of c-Jun. Cell Death Dis. 2017, 8, 3215. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Zhan, Y.; Yang, S.; Ren, S.; Qiu, X.; Rehamn, Z.U.; Tan, L.; Sun, Y.; Meng, C.; Song, C.; et al. Newcastle disease virus infection triggers HMGB1 release to promote the inflammatory response. Virology 2018, 525, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Guo, K.; Liu, C.; Wang, J.; Tan, D.; Han, X.; Tang, C.; Zhang, Y.; Wang, J. Strong Inflammatory responses and apoptosis in the oviducts of egg-laying hens caused by genotype VIId Newcastle disease virus. BMC Vet. Res. 2016, 12, 155. [Google Scholar] [CrossRef] [PubMed]
- Subbaiah, K.C.V.; Valluru, L.; Rajendra, W.; Ramamurthy, C.; Thirunavukkarusu, C.; Subramanyam, R. Newcastle disease virus (NDV) induces protein oxidation and nitration in brain and liver of chicken: Ameliorative effect of vitamin E. Int. J. Biochem. Cell Biol. 2015, 64, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Kristeen-Teo, Y.W.; Yeap, S.K.; Tan, S.W.; Omar, A.R.; Ideris, A.; Tan, S.G.; Alitheen, N.B. The effect of different velogenic NDV infections on the chicken bursa of fabricus. BMC Vet. Res. 2017, 13, 151. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhu, J.; Li, D.; Wang, Y.; Zhan, Y.; Tan, L.; Qiu, X.; Sun, Y.; Song, C.; Meng, C.; et al. Newcastle disease virus infection induces activation of the NLRP3 inflammasome. Virology 2016, 496, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xie, P.; Sun, M.; Xiang, B.; Kang, Y.; Gao, P.; Zhu, W.; Ning, Z.; Ren, T. S1PR1 expression correlates with inflammatory responses to Newcastle disease virus infection. Infect. Genet. Evol. 2016, 37, 37–42. [Google Scholar] [CrossRef]
- Li, Y.; Jiang, W.; Niu, Q.; Sun, Y.; Meng, C.; Tan, L.; Song, C.; Qiu, X.; Liao, Y.; Ding, C. elF2α-CHOP-BCL-2/JNK and IRE1α-XBP1/JNK signaling promote apoptosis and inflammation and support the proliferation of Newcastle disease virus. Cell Death Dis. 2019, 10, 891. [Google Scholar] [CrossRef]
- Li, X.; Jia, Y.; Liu, H.; Wang, X.; Chu, Z.; Hu, R.; Ren, J.; Xiao, S.; Zhang, S.; Wang, X.; et al. High level expression of ISG12(1) promotes cell apoptosis via the mitochondrial-dependent pathway and so as to hinder Newcastle disease virus replication. Vet. Microbiol. 2019, 228, 147–156. [Google Scholar] [CrossRef]
- Hu, Z.; Gu, H.; Hu, J.; Hu, S.; Wang, X.; Liu, X.; Jiao, X.; Liu, X. Quantitative proteomics identify an association between extracellular matrix degradation and immunopathology of genotype VII Newcastle disease virus in the spleen in chickens. J. Proteom. 2018, 181, 201–212. [Google Scholar] [CrossRef]
- Lim, T.-H.; Youn, H.-N.; Yuk, S.-S.; Kwon, J.-H.; Hong, W.-T.; Gwon, G.-B.; Lee, J.-A.; Lee, J.-B.; Lee, S.-W.; Song, C.-S. Successful cross-protective efficacy induced by heat-adapted live attenuated nephropathogenic infectious bronchitis virus derived from a natural recombinant strain. Vaccine 2015, 33, 7370–7374. [Google Scholar] [CrossRef] [PubMed]
- Jackwood, M.W.; Lee, D.-H. Different evolutionary trajectories of vaccine-controlled and non-controlled avian infectious bronchitis viruses in commercial poultry. PLoS ONE 2017, 12, e0176709. [Google Scholar] [CrossRef] [PubMed]
- Ellis, S.; Keep, S.; Britton, P.; de Wit, S.; Bickerton, E.; Vervelde, L. Recombinant infectious bronchitis viruses expressing chimeric spike glycoproteins induce partial protective immunity against homologous challenge despite limited replication in vivo. J. Virol. 2018, 92, e01473-18. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Zhao, J.; Xie, D.; Huang, X.; Cheng, J.; Guo, Y.; Liu, C.; Ma, Z.; Yang, H.; Zhang, G. Attenuation, safety, and efficacy of a QX-like infectious bronchitis virus serotype vaccine. Vaccine 2018, 36, 1880–1886. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; He, X.; Du, L.-J.; Liu, Y.-Y.; You, G.-J.; Li, S.-Y.; Liu, P.; Cao, S.-J.; Han, X.-F.; Huang, Y. Preparation and protective efficacy of a chicken embryo kidney cell-attenuation GI-19/QX-like avian infectious bronchitis virus vaccine. Vaccine 2018, 36, 4087–4094. [Google Scholar] [CrossRef]
- Leyson, C.M.; Hilt, D.A.; Jordan, B.J.; Jackwood, M.W. Minimum infectious dose determination of the Arkansas Delmarva poultry industry infectious bronchitis virus vaccine delivered by hatchery spray cabinet. Avian Dis. 2017, 61, 123–127. [Google Scholar] [CrossRef]
- Albanese, G.A.; Lee, D.-H.; Cheng, I.-H.N.; Hilt, D.A.; Jackwood, M.W.; Jordan, B.J. Biological and molecular characterization of ARKGA: A novel Arkansas serotype vaccine that is highly attenuated, efficacious and protective against homologous challenge. Vaccine 2018, 36, 6077–6086. [Google Scholar] [CrossRef]
- Zhang, Y.; Huang, S.; Zeng, Y.; Xue, C.; Cao, Y. Rapid development and evaluation of a live attenuated QX-like infectious bronchitis virus vaccine. Vaccine 2018, 36, 4245–4254. [Google Scholar] [CrossRef]
- Feng, K.; Xue, Y.; Wang, J.; Chen, W.; Chen, F.; Bi, Y.; Xie, Q. Development and efficacy of a novel live-attenuated nephropathogenic infectious bronchitis virus in China. Vaccine 2015, 33, 1113–1120. [Google Scholar] [CrossRef]
- Lee, H.J.; Youn, H.N.; Kwon, J.S.; Lee, Y.J.; Kim, J.H.; Lee, J.B.; Park, S.Y.; Choi, I.S.; Song, C.S. Characterization of a novel live attenuated infectious bronchitis virus vaccine candidate derived from a Korean nephropathogenic strain. Vaccine 2010, 28, 2887–2894. [Google Scholar] [CrossRef]
- Erfanmanesh, A.; Ghalyanchilangeroudi, A.; Nikaein, D.; Hosseini, H.; Mohajerfar, T. Evaluation of inactivated vaccine of the variant 2 (IS-1494/GI-23) genotype of avian infectious bronchitis. Comp. Immunol. Microbiol. Infect. Dis. 2020, 71, 101497. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Kilany, W.H.; El-Abideen, M.A.Z.; Sayed, M.E.; Elkady, M. Safety and efficacy of attenuated classic and variant 2 infectious bronchitis virus candidate vaccines. Poult. Sci. 2018, 97, 4238–4244. [Google Scholar] [CrossRef] [PubMed]
- Elhady, M.A.; Ali, A.; Kilany, W.H.; Elfeil, W.K.; Ibrahim, H.; Nabil, A.; Samir, A.; El Sayed, M. Field efficacy of an attenuated infectious bronchitis variant 2 vaccine in commercial broiler chickens. Vet. Sci. 2018, 5, 49. [Google Scholar] [CrossRef] [PubMed]
- Jackwood, M.W.; Hilt, D.A.; Sellers, H.S.; Williams, S.M.; Lasher, H.N. Rapid heat-treatment attenuation of infectious bronchitis virus. Avian Pathol. 2010, 39, 227–233. [Google Scholar] [CrossRef]
- Bijlenga, G.; Cook, J.K.; Gelb, J.J.; Wit, J.J. Development and use of the H strain of avian infectious bronchitis virus from Netherlands as a vaccine: A review. Avian pathol. 2004, 33, 550–557. [Google Scholar] [CrossRef]
- Han, Z.; Zhang, T.; Xu, Q.; Gao, M.; Chen, Y.; Wang, Q.; Zhao, Y.; Shao, Y.; Li, H.; Kong, X.; et al. Altered pathogenicity of a tl/CH/LDT3/03 genotype infectious bronchitis coronavirus due to a natural recombination in the 5’-17kb region of the genome. Virus Res. 2015, 213, 140–148. [Google Scholar] [CrossRef]
- Zhao, Y.; Xie, D.; Zhang, K.; Cheng, J.; Xu, G.; Zhang, G. Pathogenicity of a GI-22 genotype infectious bronchitis virus isolated in China and protection against it afforded by GI-19 vaccine. Virus Res. 2019, 267, 59–66. [Google Scholar] [CrossRef]
- Jiang, Y.; Cheng, X.; Zhao, X.; Yu, Y.; Gao, M.; Zhou, S. Recombinant infectious bronchitis coronavirus H120 with the spike protein S1 gene of the nephropathogenic IBYZ strain remains attenuated but induces protective immunity. Vaccine 2020, 38, 3157–3168. [Google Scholar] [CrossRef]
- Armesto, M.; Evans, S.; Cavanagh, D.; Abu-Median, A.-B.; Keep, S.; Britton, P. A recombinant avian infectious bronchitis expressing a heterologous spike gene belonging to the 4/91 serotype. PLoS ONE 2011, 6, e24352. [Google Scholar] [CrossRef]
- Yang, H.-M.; Zhao, J.; Xue, J.; Yang, Y.-L.; Zhang, G.-Z. Antigenic variation of LaSota and genotype VII Newcastle disease virus (NDV) and their efficacy against challenge with velogenic NDV. Vaccine 2017, 35, 27–32. [Google Scholar] [CrossRef]
- Huang, Y.P.; Wang, C.H. Development of attenuated vaccines from Taiwanese infectious bronchitis virus strain. Vaccine 2006, 24, 785–791. [Google Scholar] [CrossRef] [PubMed]
- Manoharan, V.K.; Varghese, B.P.; Paldurai, A.; Samal, S.K. Effect of fusion protein cleavage site sequence on generation of a genotype VII Newcastle disease virus vaccine. PLoS ONE 2018, 13, e0197253. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Xiang, B.; Li, Y.; Xie, P.; Gao, S.; Kang, Y.; Gao, P.; Li, Y.; Wang, Z.; Liang, J.; et al. Generation and evaluation of a genetically attenuated Newcastle disease virus rGM-VIIm as a genotype-matched vaccine. Virus Genes 2017, 53, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Ji, H.; Peng, L.; Gao, X.; Jiang, S. Development of PLGA-PEG-PLGA hydrogel delivery system for enhanced immunoreaction and efficacy of Newcastle disease virus DNA vaccine. Molecules 2020, 25, 2505. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kye, S.J.; Lee, H.J.; Gaikwad, S.; Lee, H.S.; Jung, S.C.; Choi, K.S. Development of a highly immunogenic Newcastle disease virus chicken vaccine strain of duck origin. Poult. Sci. 2016, 95, 790–797. [Google Scholar] [CrossRef]
- Mebrahtu, K.; Teshale, S.; Esatu, W.; Habte, T.; Gelaye, E. Evaluation of spray and oral delivery of Newcastle disease I2 vaccine in chicken reared by small holder farmers in central Ethiopia. BMC Vet. Res. 2018, 14, 48. [Google Scholar] [CrossRef]
- Perozo, F.; Villegas, P.; Dolz, R.; Afonso, C.L.; Purvis, L.B. The VG/GA strain of Newcastle disease virus: Mucosal immunity, protection against lethal challenge and molecular analysis. Avian Pathol. 2008, 37, 237–245. [Google Scholar] [CrossRef]
- Li, S.-Y.; You, G.-J.; Du, J.-T.; Xia, J.; Wen, Y.-P.; Huang, X.-B.; Zhao, Q.; Han, X.-F.; Yan, Q.-G.; Wu, R.; et al. A class I lentogenic Newcastle disease virus strain confers effective protection against the prevalent strains. Biologicals 2020, 63, 74–80. [Google Scholar] [CrossRef]
- Hu, Z.; Hu, S.; Meng, C.; Wang, X.; Zhu, J.; Liu, X. Generation of a genotype VII Newcastle disease virus vaccine candidate with high yield in embryonated chicken eggs. Avian Dis. 2011, 55, 391–397. [Google Scholar] [CrossRef]
- Xiao, S.; Nayak, B.; Samuel, A.; Paldurai, A.; Kanabagattebasavarajappa, M.; Prajitno, T.Y.; Bharoto, E.E.; Collins, P.L.; Samal, S.K. Generation by reverse genetics of an effective, stable, live-attenuated Newcastle disease virus vaccine based on a currently circulating, highly virulent Indonesian strain. PLoS ONE 2012, 7, e52751. [Google Scholar] [CrossRef]
- Liu, J.; Zhu, J.; Xu, H.; Li, J.; Hu, Z.; Hu, S.; Wang, X.; Liu, X. Effects of the HN antigenic difference between the vaccine strain and the challenge strain of Newcastle disease virus on virus shedding and transmission. Viruses 2017, 9, 225. [Google Scholar] [CrossRef] [PubMed]
- Roohani, K.; Tan, S.W.; Yeap, S.K.; Ideris, A.; Bejo, M.H.; Omar, A.R. Characterisation of genotype VII Newcastle disease virus (NDV) isolated from NDV vaccinated chickens, and the efficacy of LaSota and recombinant genotype VII vaccines against challenge with velogenic NDV. J. Vet. Sci. 2015, 16, 447–457. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.-M.; Cheng, J.-L.; Yu, X.-H.; Qin, Z.-M.; Tian, F.-L.; Zhang, G.-Z. Generation by reverse genetics of an effective attenuated Newcastle disease virus vaccine based on a prevalent highly virulent Chinese strain. Biotechnol. Lett. 2015, 37, 1287–1296. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo-Lara, R.; Chumbe, A.; Calderon, K.; Fernandez-Diaz, M.; Vakharia, V.N. Genotype-matched Newcastle disease virus vaccine confers improved protection against genotype XII challenge: The importance of cytoplasmic tails in viral replication and vaccine design. PLoS ONE 2019, 14, e0209539. [Google Scholar] [CrossRef]
- Bello, M.B.; Mahamud, S.N.A.; Yusoff, K.; Ideris, A.; Hair-Bejo, M.; Peeters, B.P.H.; Omar, A.R. Development of an effective and stable genotype-matched live attenuated Newcastle disease virus vaccine based on a novel naturally recombinant Malaysian isolate using reverse genetics. Vaccines 2020, 8, 270. [Google Scholar] [CrossRef]
- Yadav, K.; Pathak, D.C.; Saikia, D.P.; Debnath, A.; Ramakrishnan, S.; Dey, S.; Chellappa, M.M. Generation and evaluation of a recombinant Newcastle disease virus strain R2B with an altered fusion protein cleavage site as a vaccine candidate. Microb. Pathog. 2018, 118, 230–237. [Google Scholar] [CrossRef]
- Bande, F.; Arshad, S.S.; Bejo, M.H.; Kdkhodaei, S.; Omar, A.R. Prediction and insilico identification of novel B-cells and T-cells epitopes in the S1-spike glycoprotein of M41 and CR88 (793/B) infectious bronchitis virus serotypes for application in peptide vaccines. Adv. Bioinform. 2016, 2016, 1–5. [Google Scholar] [CrossRef]
- Li, J.; Helal, Z.H.; Karch, C.P.; Mishra, N.; Girshick, T.; Garmendia, A.; Burkhard, P.; Khan, M.I. A self-adjuvanted nanoparticle-based vaccine against infectious bronchitis virus. PLoS ONE 2018, 13, e0203771. [Google Scholar] [CrossRef]
- Wu, Q.; Lin, Z.; Qian, K.; Shao, H.; Ye, J.; Qin, A. Peptides with 16R in S2 protein showed broad reactions with sera against different types of infectious bronchitis viruses. Vet. Microbiol. 2019, 236, 108391. [Google Scholar] [CrossRef]
- Zhao, K.; Zhang, Y.; Zhang, X.; Li, W.; Shi, C.; Guo, C.; Dai, C.; Chen, Q.; Jin, Z.; Zhao, Y.; et al. Preparation and efficacy of Newcastle disease virus DNA vaccine encapsulated in chitosan nanoparticles. Int. J. Nanomed. 2014, 4, 389–402. [Google Scholar] [CrossRef]
- Zhao, K.; Zhang, Y.; Zhang, X.; Shi, C.; Wang, X.; Wang, X.; Jin, Z.; Cui, S. Chitosan-coated poly (lactic-co-glycolytic) acid nanoparticle as an efficient delivery system for Newcastle disease virus DNA vaccine. Int. J. Nanomed. 2014, 9, 4609–4619. [Google Scholar] [CrossRef] [PubMed]
- Kang, X.; Wang, J.; Jiao, Y.; Tang, P.; Song, L.; Xiong, D.; Yin, Y.; Pan, Z.; Jiao, X. Expression of recombinant Newcastle disease virus F protein in Pichia pastoris and its immunogenicity using flagellin as the adjuvant. Protein Expr. Purif. 2016, 128, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Khulape, S.A.; Maity, H.K.; Pathak, D.C.; Ramamurthy, N.; Ramakrishnan, S.; Chellappa, M.M.; Dey, S. Evaluation of a fusion gene-based DNA prime-protein boost vaccination strategy against Newcastle disease virus. Trop. Anim. Health Prod. 2019, 51, 2529–2538. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.; Zhang, E.; Li, Q.; Xu, Q.; Ou, J.; Yin, H.; Li, K.; Wang, L.; Zhao, X.; Niu, X.; et al. A Plant-produced recombinant fusion protein-based Newcastle disease subunit vaccine and rapid differential diagnosis platform. Vaccines 2020, 8, 122. [Google Scholar] [CrossRef] [PubMed]
- Shahriari, A.G.; Bagheri, A.; Afsharifar, A.; Habibi-Pirkooshi, M. Induction of immune response in animal model using recombinant anti-NDV vaccine. Iran. J. Biotechnol. 2019, 17, e2215. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Sun, J.; Qi, T.; Zhao, W.; Han, Z.; Yang, X.; Liu, S. Recombinant Newcastle disease virus expressing the infectious bronchitis virus S1 gene protects chickens against Newcastle disease virus and infectious bronchitis virus challenge. Vaccine 2017, 35, 2435–2442. [Google Scholar] [CrossRef]
- Shirvani, E.; Paldurai, A.; Manoharan, V.K.; Varghese, B.P.; Samal, S.K. A recombinant Newcastle disease virus (NDV) expressing S protein of infectious bronchitis virus (IBV) protects chickens against IBV and NDV. Sci. Rep. 2018, 8, 11951. [Google Scholar] [CrossRef]
- Abozeid, H.H.; Paldurai, A.; Varghese, B.P.; Khattar, S.K.; Afifi, M.A.; Zouelfakkar, S.; EL-Deeb, A.H.; EL-Kady, M.F.; Samal, S.K. Development of a recombinant Newcastle disease virus-vectored vaccine for infectious bronchitis virus variant strains circulating in Egypt. Vet. Res. 2019, 50, 12. [Google Scholar] [CrossRef]
- Tan, L.; Wen, G.; Qiu, X.; Yuan, Y.; Meng, C.; Sun, Y.; Liao, Y.; Song, C.; Liu, W.; Shi, Y.; et al. A recombinant LaSota vaccine strain expressing multiple epitopes of infectious bronchitis virus (IBV) protects specific pathogen-free (SPF) chickens against IBV and NDV challenges. Vaccines 2019, 7, 170. [Google Scholar] [CrossRef]
- Yang, X.; Zhou, Y.; Li, J.; Fu, L.; Ji, G.; Zeng, F.; Zhou, L.; Gao, W.; Wang, H. Recombinant infectious bronchitis virus (IBV) H120 vaccine strain expressing the hemagglutinin-neuraminidase (HN) protein of Newcastle disease virus (NDV) protects chicken against IBV and NDV challenge. Arch. Virol. 2016, 161, 1209–1216. [Google Scholar] [CrossRef]
- Purchase, H.G.; Okazaki, W.; Burmester, B.R. Long-term field trials with herpesvirus of turkeys vaccine against Marek’s disease. Avian Dis. 1972, 16, 57–71. [Google Scholar] [CrossRef] [PubMed]
- Ingrao, F.; Rauw, F.; van de Berg, T.; Lambrecht, B. Characterization of two recombinant HVT-IBD vaccines by VP2 insert detection of specific pathogen-free chickens. Avian Pathol. 2017, 46, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Le Gros, F.X.; Dancer, A.; Giacomini, C.; Pizzoni, L.; Bublot, M.; Graziani, M.; Prandini, F. Field efficacy trial of a novel HVT-IBD vector vaccine for 1-day-old broilers. Vaccine 2009, 27, 592–596. [Google Scholar] [CrossRef] [PubMed]
- Lamiere, S.; Fernández, R.; Pritchard, N.; Cruz-Coy, J.; Rojo, F.; Wong, S.Y.; Saint-Gerand, A.L.; Gauthier, J.C.; Perozo, F. Concomitant turkey herpesvirus-infectious bursal disease vector vaccine and oil-adjuvanted inactivated Newcastle disease vaccine administration; consequences for vaccine intake and protection. Avian Dis. 2011, 55, 642–649. [Google Scholar] [CrossRef]
- Palya, V.; Kiss, I.; Tatár-Kis, T.; Mató, T.; Felföldi, B.; Gardin, Y. Advancement in vaccination against Newcastle disease: Recombinant HVT NDV provides high clinical protection and reduces challenge virus shedding with the absence of vaccine reactions. Avian Dis. 2012, 56, 282–287. [Google Scholar] [CrossRef] [PubMed]
- Rauw, F.; Gardin, Y.; Palya, V.; van der Berg, T.; Lambrecht, B. The combination of attenuated Newcastle disease (ND) vaccine with rHVT-ND vaccine at 1 day old is more protective against ND virus challenge than when combined with inactivated ND vaccine. Avian Pathol. 2014, 43, 26–36. [Google Scholar] [CrossRef]


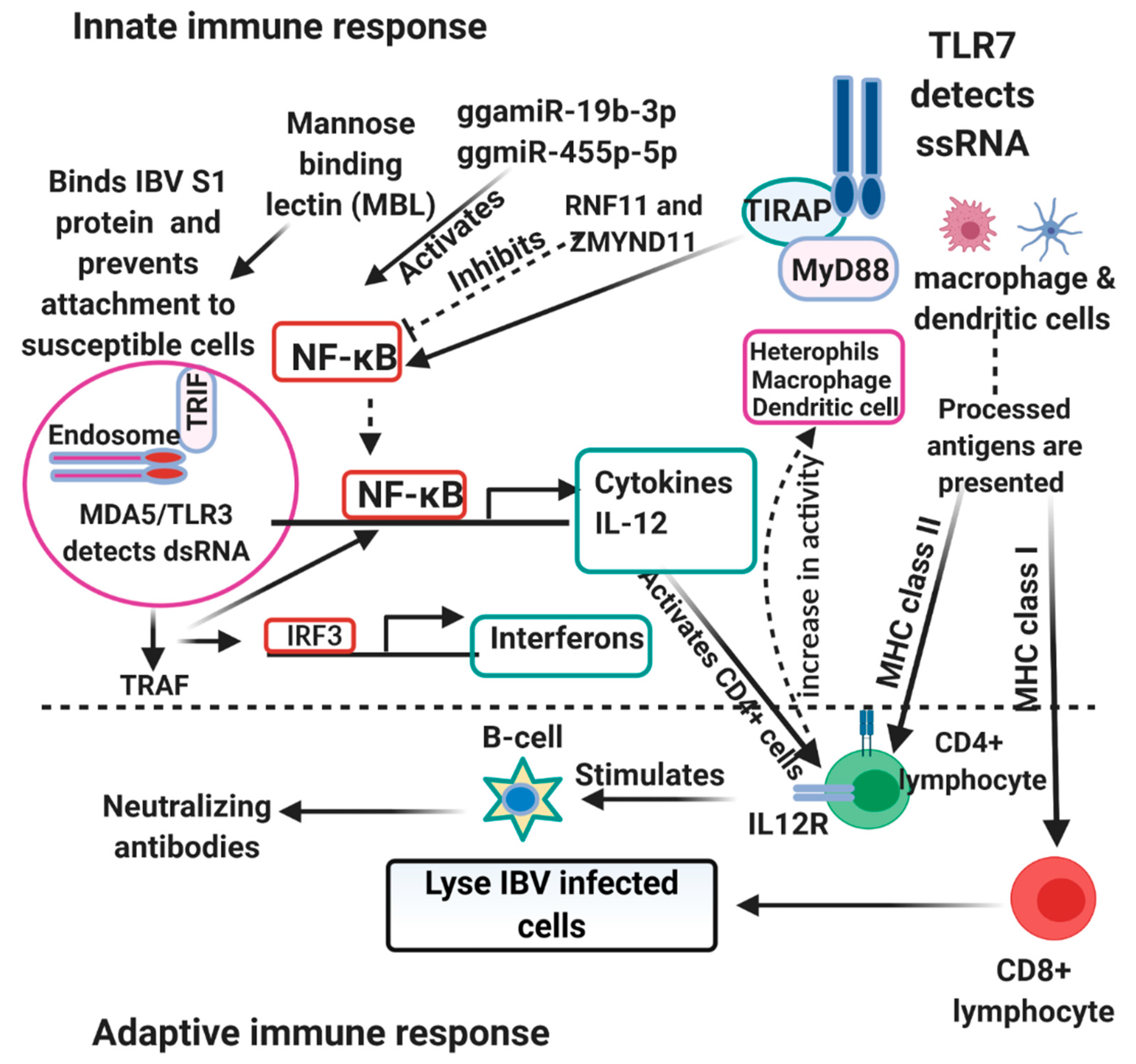{kind=link}
| S/N | Vaccine Name | Type of Vaccine | Formulation | Parent Strain | Challenge Strain | Observed Response |
|---|---|---|---|---|---|---|
| 1 | SZ130 [127] | Live attenuated vaccine | Continuous passage in chicken embryo for 130 generations | Attenuated QX-like IBV strain SZ130 | QX-like IBV strain SD | Reduces the proliferation efficiency of the challenge strain |
| 2 | IBV-CS [88] | Inactivated vaccine | Ionic gelation method (IBV, BR-1 genotype strain encapsulated in chitosan nanoparticles) | IBV, BR-1 genotype strain | IBVPR-05 strain | IgA- and IgY-mediated mucosal immune responses and T-cell-mediated immunity (CMI) responses |
| 3 | SczyC100 [128] | Live attenuated vaccine | Continuous passage in chicken embryo kidney cells for 100 times | Attenuated GI-19/QX-like field isolate Sczy3 | GI-19/QX and GI-7/TWI type virulent strains | Reduced morbidity, mortality, and tracheal viral loads caused by the challenge strains |
| 4 | ArkDPI [129] | Live attenuated vaccine | Passaged 50 times in embryonated eggs and further passaged by individual vaccine companies to generate different commercial ArkDPI-derived Ark serotype IBV vaccines | Attenuated Ark-type IBV | Ark IBV serotype | Induced protection against homologous challenge |
| 5 | ArkGA [130] | Live attenuated vaccine | Continuously passaged 60 times in 9-to-11 day-old embryonated chicken eggs | Ark99 vaccine no longer commercially used | Ark IBV serotype | Induces efficacious immune response against the challenge strain |
| 6 | ZYYR-2014 [131] | Live attenuated vaccine | Limiting dilution passage attenuation in embryonated chicken eggs (five passages) | Attenuated QX-like IBV field strain ZYY-2014 | QX-like (HSJ-2016) and Mass (M41) types. | Provided full protection against the IBV challenge strain |
| 7 | K40/09 [124] | Live attenuated vaccine | Continuous passage in embryonated eggs for 50 times at sub-optimal higher temperature (56 °C) | Attenuated K40/09 strain | KM91-like and QX-like subgroup | Elicited high titers of neutralizing antibodies against the challenge strains. |
| 8 | YX10p90 [132] | Live attenuated vaccine | Continuous passage in fertilized chicken eggs for 90 times | Attenuated IB, QX-like YX10 strain | YXp5, CHI, CHIII, and CHV genotypes | Showed 100% protection against YX10p5, effective protection against CHI genotype strains, and partial protection against CHIII and CHV genotype strain |
| 9 | K2p170 [133] | Live attenuated vaccine | Continuous passage in embryonated chicken eggs for 170 times | Attenuated K2/01 strain | KM91-like and QX-like subgroup | Produced neutralizing antibodies against challenge strains and provided almost complete protection |
| 10 | Variant 2 [134] | Inactivated vaccine | Inactivated after 10th passage in embryo with formaldehyde, passaged 3 times after inactivation and emulsified | Variant 2 (IS-1494/GI-23) genotype | IBV variant 2 viruses | Elicited neutralizing antibodies against the challenge strain |
| 11 | att-IBM41 [135] | Live attenuated vaccine | Continuous passage in embryonated chicken eggs for 100 times | Attenuated IBM41 strain | IBM41 strain | Reduced viral shedding |
| 12 | att-IB2 [135] | Live attenuated vaccine | Continuous passage in embryonated chicken eggs for 100 times | Attenuated IB2 strain | IBM41 strain | Reduced viral shedding |
| 13 | ME VAC IB-VAR2 [135,136] | Live attenuated vaccine | Continuous passage in embryonated chicken eggs for 110 times | Attenuated variant 2 IBV strain Eg/1212B/2012 | Variant 2 strains (Egy/VarII | Reduced viral shedding, Produced neutralizing antibodies against the challenge strain |
| 14 | GA08/GA08HSp16/08 [137] | Live attenuated vaccine | Continuous heat treatment (56 °C) passage in embryonated chicken eggs for 8 times, followed by final passage for 4 times without heat treatment | Attenuated GA08/pass4/08 strain | GA08/pass4/08 strain | Induced protective immune response against challenge strain |
| 15 | IB H120 [138] | Live attenuated vaccine | Passage in embryonated chicken eggs for 120 times | Attenuated mass type IB H strain | Recommended for IBV Massachusetts type | Stimulates both humoral and cellular immune responses to challenge strains |
| 16 | LDT3-A [139] | Live attenuated vaccine | Continuous passaging in chicken embryos for 120 times | Attenuated tl/CH/LDT3/03 strain | tl/CH/LDT3/03 strain | Provided protection against challenge strain |
| 17 | GI-19 (SZ200) [140] | Live attenuated vaccine | Continuous passage in embryonated chicken eggs for 200 times. | Attenuated GI-19 vaccine strain SZ200 | GI-19 and GI-22 genotype strains | Provided protection against the challenge strain |
| 18 | rH120-S1/YZ [141] | Live attenuated vaccine | Reverse genetics, S1 gene of H120 vaccine strain replaced with that of ck/CH/IBYZ/2011 | H120 vaccine strain | rIBYZ a QX-like strain | stimulated both humoral and mucosal immunity to the vaccinated birds |
| 19 | BeauR-4/91(S) [142] | Live attenuated vaccine | Reverse genetics, the ectodomain region of Beaudette S glycoprotein replaced by that of IBV 4/91 | Beau-R strain | IBV 4/91 and M41-CK strain | Provided protection against challenge with 4/91 but less protection against IBV M41 |
| 20 | IB TW [143] | Live attenuated vaccine | Passage in specific pathogen-free (SPF) embryonated eggs 74 times | IBV strain 2575/98 a Taiwan Group I strain (TW I) | IBV strain 2993/02 (TW I) | Provided 90% protection against challenge with pathogenic field IBV 2993/02 and stimulated neutralization antibodies with neutralization index greater than 4.4 |
| S/N | Vaccine Name | Type of Vaccine | Formulation | Parent Strain | Challenge Strain | Observed Response |
|---|---|---|---|---|---|---|
| 1 | NDRL0901 [148] | Live attenuated vaccine | Passaged six (6) times in specific pathogen-free (SPF) chickens | DK1307 strain of duck origin | Kr005 strain | Showed >80% protection from mortality irrespective of administration route. |
| 2 | ND I2 [149] | Thermostable Live vaccine | Passaged two (2) times in the allantoid cavity of embryonated chicken egg. | NDV I2 strain | Local NDV strain | Induces antibody response and protective immunity against challenge strain |
| 3 | VG/GA [150] | Live attenuated vaccine | Passaged three times in embryonated chicken egg. | VG/GA strain | NDV GB strain | Provided 95–100% protection against challenge strain |
| 4 | LaSota and PT3 [151] | Live vaccine | Lentogenic live strains | LaSota strain and PT3 strain | NDV SD strain, NDV DY strain | Provided full protection against challenge strains |
| 5 | NDV/A14 [152] | Inactivated vaccine | Amino-acid sequence of the cleavage site of the F0 protein was changed by reverse genetics, passaged 10 times in chicken embryo and inactivated with 0.7% formaldehyde | NDV genotype VII JS3/05 strain | NDV genotype VII JS3/05 strain | Reduced viral shedding and protected the birds against clinical disease |
| 6 | Ban/AF [153] | Live attenuated vaccine | Virulent F protein motif was modified to avirulent motif by reverse genetics and passed 10 times in chicken | NDV strain Ban/010 | NDV strain Ban/010 | Provided full protection against the challenge virus and prevented viral shedding |
| 7 | NDV O/A14 [154] | Inactivated vaccine | HN gene of NDV/A14 vaccine strain replaced with that of JS-14-12-Ch bearing E347K and G362A co-mutation | JS-14-12-Ch HN gene in NDV/A14 | JS-14-12-Ch NDV strain | Provided full protection, reduced viral shedding and viral loads. |
| 8 | aSG10 [155,156] | Live attenuated vaccine | Reverse genetics and the protease cleavage site of the virulent F0 protein was altered to avirulent strain | SG10 strain | SG10 strain | Provided full protection and reduced viral shedding |
| 9 | G7M [95] | Live attenuated vaccine | Reverse genetics and the replacement of F cleavage site sequence typical of velogenic strains with that of LaSota vaccine | G7 strain | Genotype VII NDV (G7 strain) | Provided full protection against the challenged virus strain, induced both humoral and cell-mediated immunity, reduced virus replication and shedding |
| 10 | rLS1-XII-2 [157] | Live attenuated vaccine | Reverse genetics, the ecto- and trans membrane domains of the F and HN protein in the rLS1 backbone were replaced with those of PP2011 | Genotype XII (PP2011) strain | Genotype XII (PP2011) strain | Provided full protection against the challenged strain and reduced virus shedding |
| 11 | rGM-VIIm [146] | Live attenuated vaccine | Reverse genetics, rGM attenuated by changing the F0 polybasic cleavage site to monobasic | Genotype VII GM strain | GM NDV strain | Both inactivated and live rGM-VIIm provided full protection against the challenge strain. However, the live vaccines reduced viral shedding more than the inactivated vaccine. |
| 12 | mIBS025 [158] | Live attenuated vaccine | Reverse genetics, in silico modification of F cleavage site from virulent polybasic to avirulent monobasic motif, modified sequence chemically synthesized and inserted into pOLTV5 transcription vector | Complete genome sequence of NDV IBS025/13 strain | NDV strain IBS002/11 | Induced strong antibody-mediated immunity and reduced viral shedding |
| 13 | rNDV-R2B-FPCS [159] | Live attenuated vaccine | Reverse genetics, changing the F protein cleavage site from polybasic amino acid to dibasic amino acid | rNDV-R2B | Virulent NDV strain (Accession No. KJ769262.1) | Stimulated both humoral and cellular immunity, reduced viral shedding |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ike, A.C.; Ononugbo, C.M.; Obi, O.J.; Onu, C.J.; Olovo, C.V.; Muo, S.O.; Chukwu, O.S.; Reward, E.E.; Omeke, O.P. Towards Improved Use of Vaccination in the Control of Infectious Bronchitis and Newcastle Disease in Poultry: Understanding the Immunological Mechanisms. Vaccines 2021, 9, 20. https://doi.org/10.3390/vaccines9010020
Ike AC, Ononugbo CM, Obi OJ, Onu CJ, Olovo CV, Muo SO, Chukwu OS, Reward EE, Omeke OP. Towards Improved Use of Vaccination in the Control of Infectious Bronchitis and Newcastle Disease in Poultry: Understanding the Immunological Mechanisms. Vaccines. 2021; 9(1):20. https://doi.org/10.3390/vaccines9010020
Chicago/Turabian StyleIke, Anthony C., Chukwuebuka M. Ononugbo, Okechukwu J. Obi, Chisom J. Onu, Chinasa V. Olovo, Sophia O. Muo, Okoro S. Chukwu, Eleazar E. Reward, and Odinakachukwu P. Omeke. 2021. "Towards Improved Use of Vaccination in the Control of Infectious Bronchitis and Newcastle Disease in Poultry: Understanding the Immunological Mechanisms" Vaccines 9, no. 1: 20. https://doi.org/10.3390/vaccines9010020
APA StyleIke, A. C., Ononugbo, C. M., Obi, O. J., Onu, C. J., Olovo, C. V., Muo, S. O., Chukwu, O. S., Reward, E. E., & Omeke, O. P. (2021). Towards Improved Use of Vaccination in the Control of Infectious Bronchitis and Newcastle Disease in Poultry: Understanding the Immunological Mechanisms. Vaccines, 9(1), 20. https://doi.org/10.3390/vaccines9010020