Immunization with Plant-Derived Multimeric H5 Hemagglutinins Protect Chicken against Highly Pathogenic Avian Influenza Virus H5N1

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Construction of Plant Expression Vectors
2.2. Transient Protein Expression in N. Benthamiana Plants
2.3. Total Soluble Protein Extraction
2.4. SDS-PAGE and Western Blotting
2.5. Protein Purification by Immobilized Metal Affinity Chromatography (IMAC)
2.6. Size Exclusion Chromatography
2.7. Mouse Immunization
2.8. Hemagglutination Test and Hemagglutination Inhibition Assay
2.9. Indirect ELISA
2.10. Competitive ELISA
2.11. Chicken Challenge Experiment with the Purified H5-TG Trimer
2.12. Chicken Challenge Experiment with Plant Crude Extracts Containing H5 Antigens
2.13. Statistical Analyses
3. Results
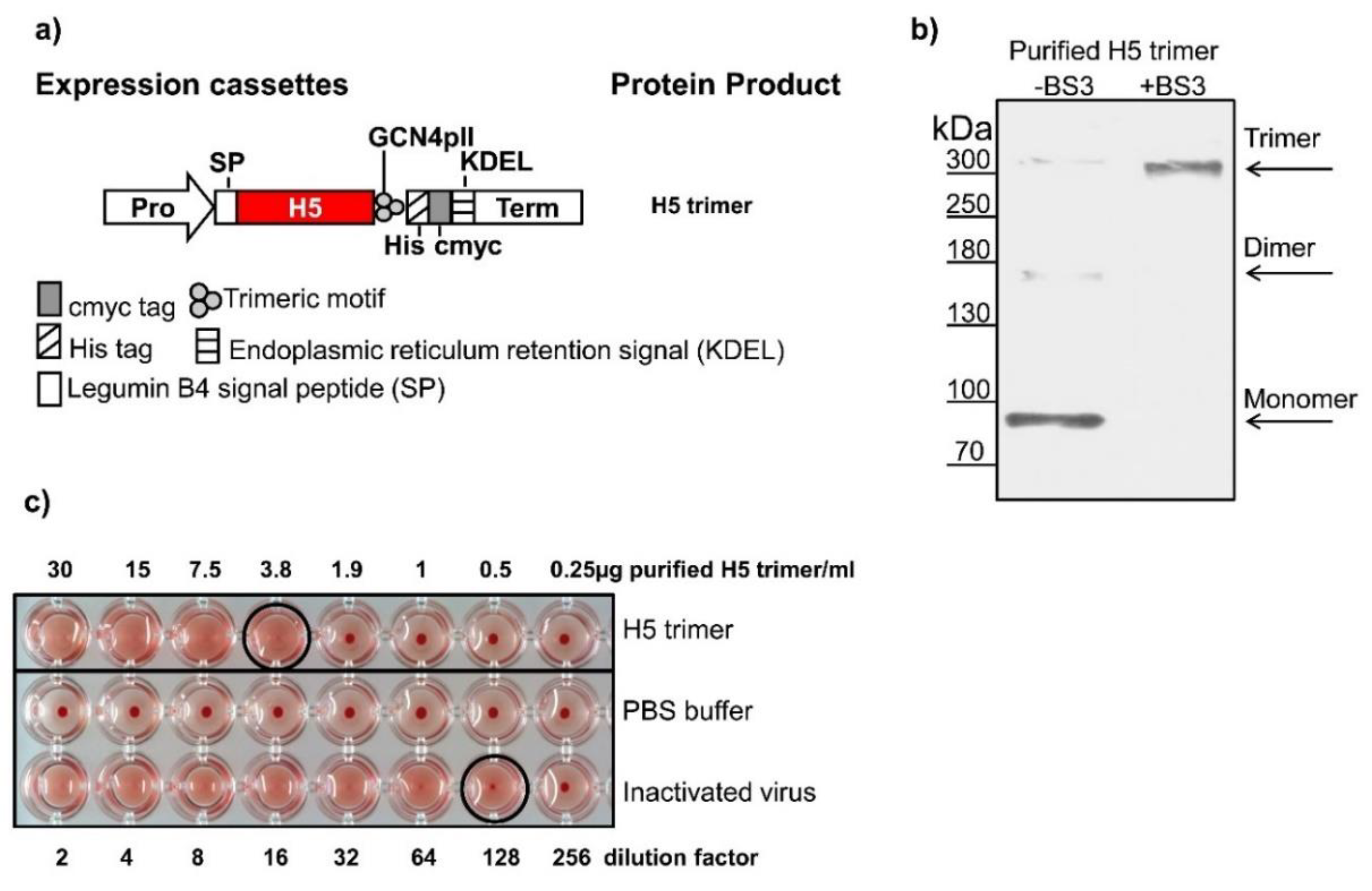
3.1. Trimerization Technology Keeps H5 Trimer and its Activity
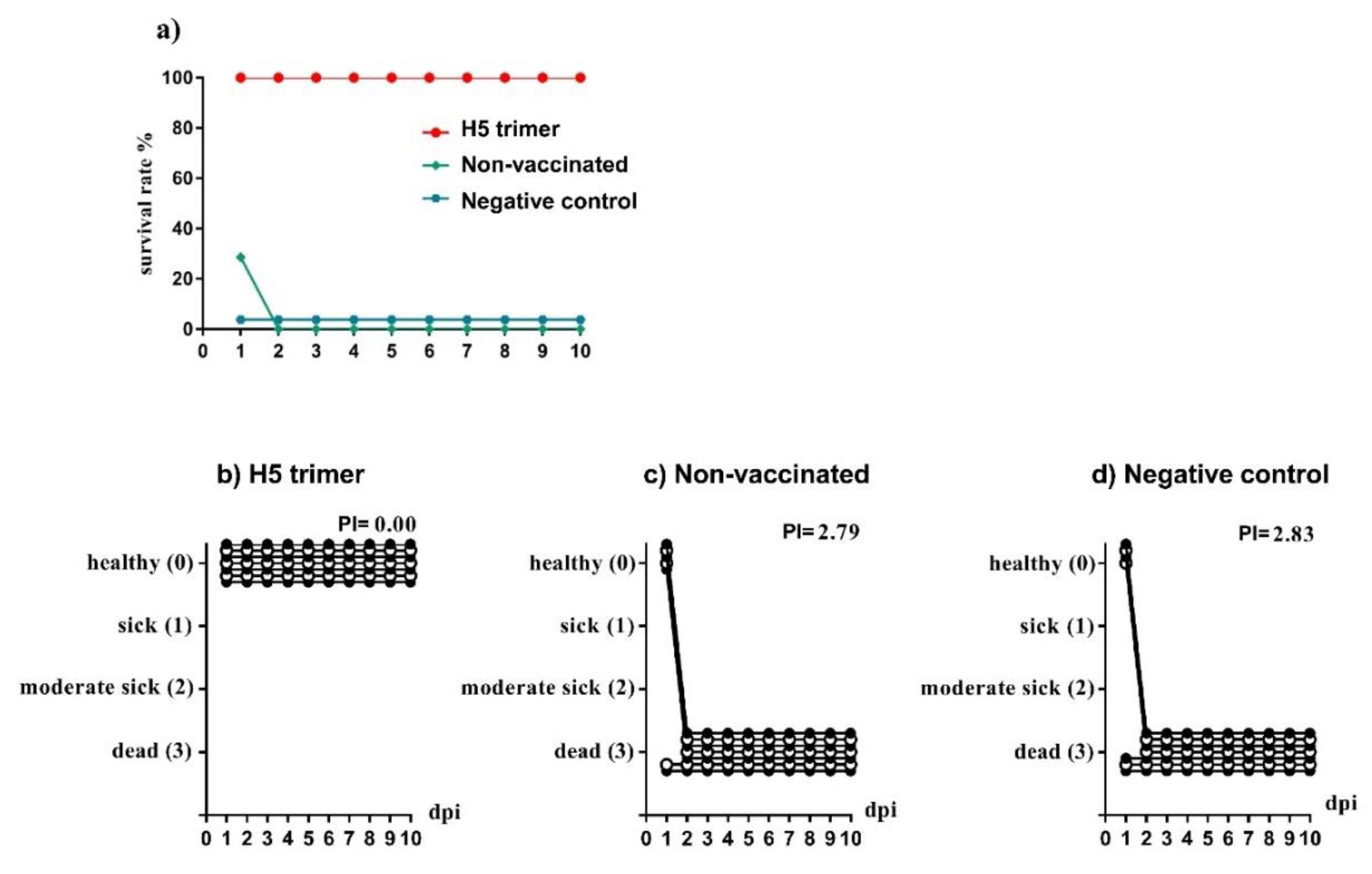
3.2. Plant-Made Purified H5 Trimer Protects Chickens from HPAIV H5N1
3.3. Multimerization of H5-TG Trimer in Plants
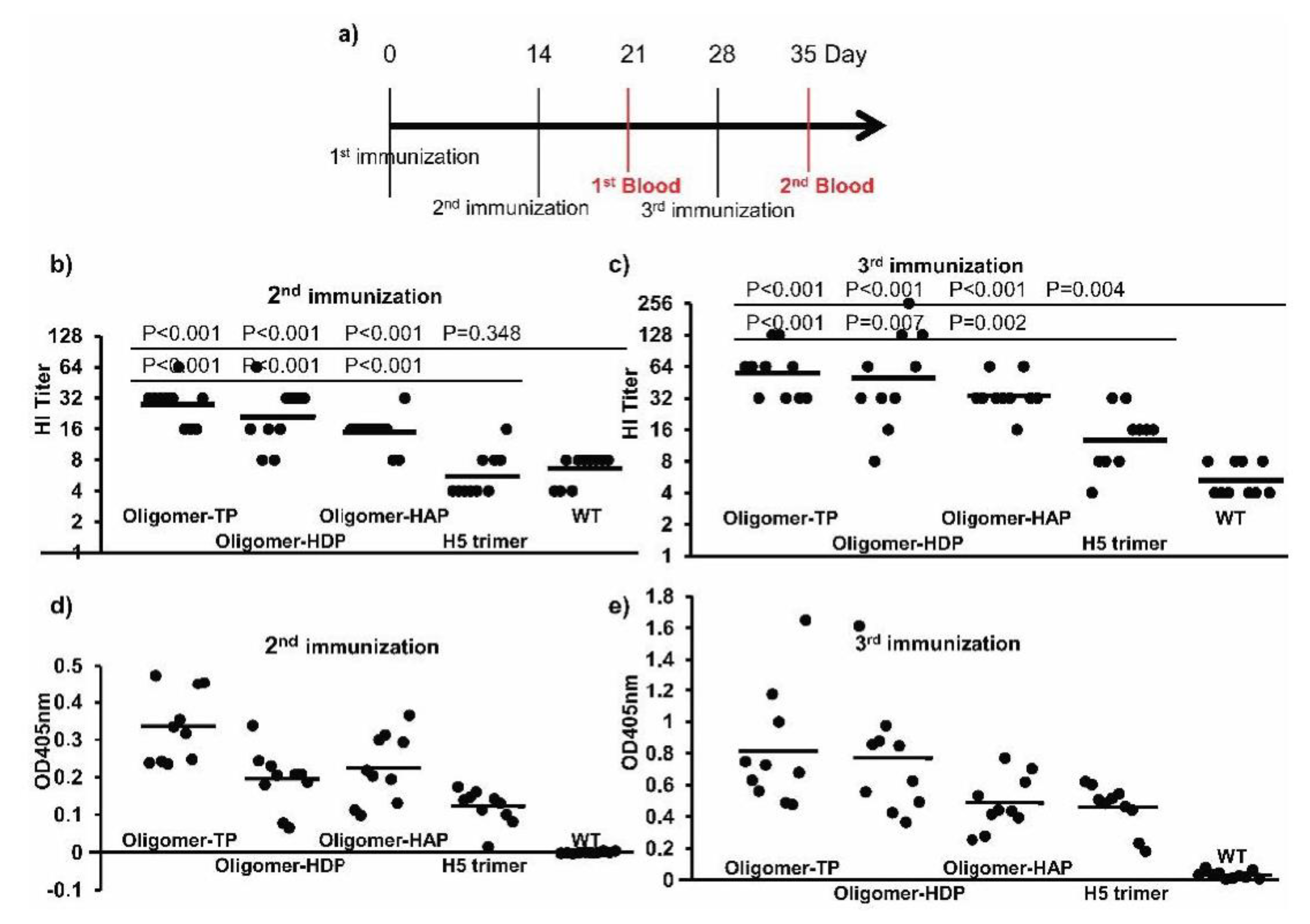
3.4. High Immunogenicity of H5 Oligomers in Plant Crude Extracts in Mice
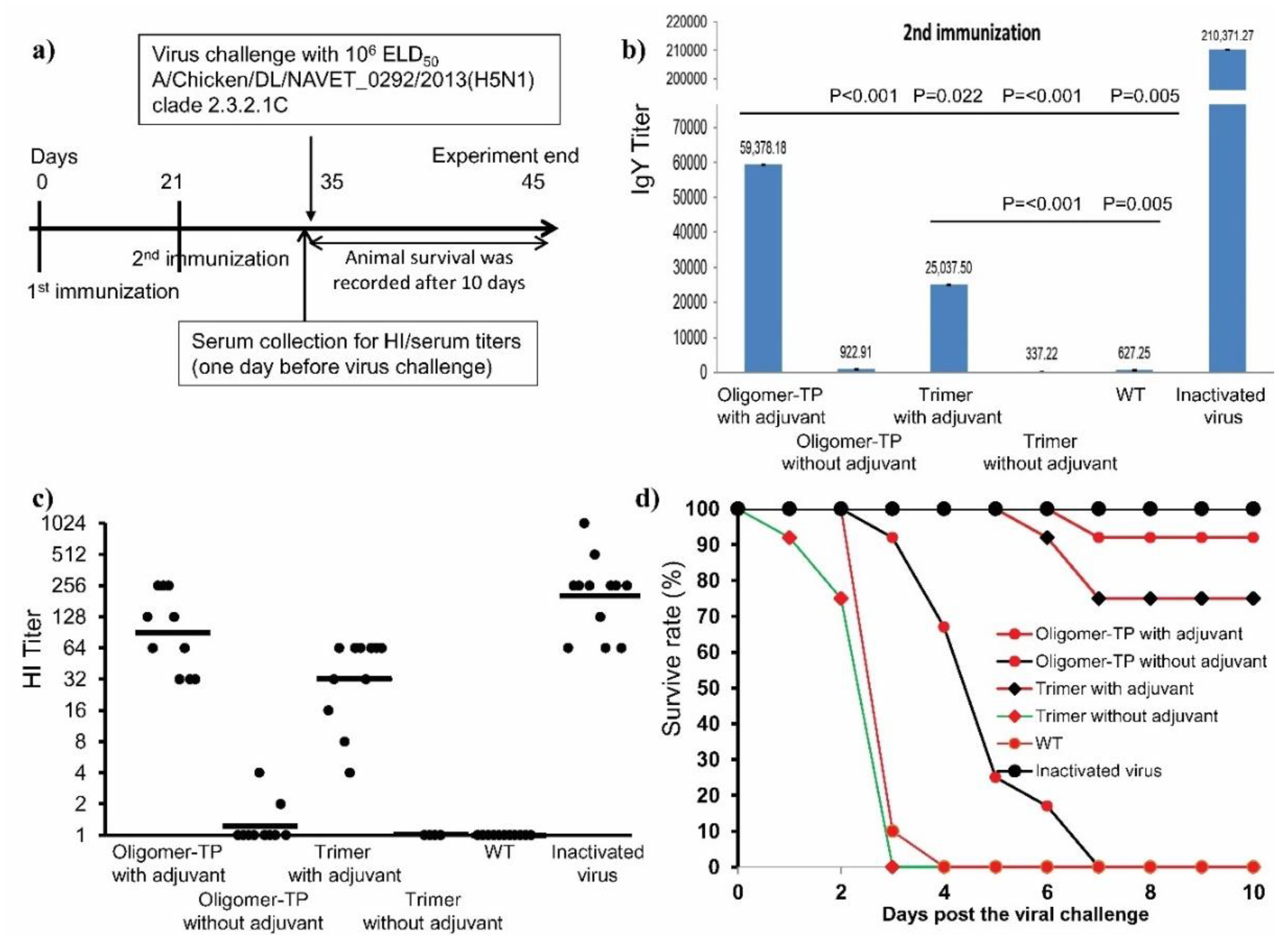
3.5. High Immunogenicity of H5 Oligomers in Plant Crude Extracts in Chickens
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- André, F.E. Vaccinology: Past achievements, present roadblocks and future promises. Vaccine 2003, 21, 593–595. [Google Scholar] [CrossRef]
- Shams, H. Recent developments in veterinary vaccinology. Vet. J. 2005, 170, 289–299. [Google Scholar] [CrossRef]
- Woolhouse, M.E.; Gowtage-Sequeria, S. Host range and emerging and reemerging pathogens. Emerg. Infect. Dis. 2005, 11, 1842–1847. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization (FAO) of the United Nations. A Global Strategy for the Progressive Control of Highly Pathogenic Avian Influenza (2005). Available online: https://pdfs.semanticscholar.org/366f/ac12e92129b3833b0bc5979bac944e291aea.pdf (accessed on 12 July 2020).
- European Centre for Disease Prevention and Control (ECDPC). Rapid Risk Assessment: Outbreaks of Highly Pathogenic Avian Influenza A (H5N8) in Europe, 18 November 2016. Available online: https://www.ecdc.europa.eu/en/publications-data/rapid-risk-assessment-outbreaks-highly-pathogenic-avian-influenza-ah5n8-europe-18 (accessed on 12 July 2020).
- Nelson, M.I.; Holmes, E.C. The evolution of epidemic influenza. Nat. Rev. Genet. 2007, 8, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Ling, H.Y.; Pelosi, A.; Walmsley, A.M. Current status of plant-made vaccines for veterinary purposes. Expert Rev. Vaccines 2010, 9, 971–982. [Google Scholar] [CrossRef]
- Cox, R.J.; Brokstad, K.A.; Ogra, P. Influenza virus: Immunity and vaccination strategies. Comparison of the immune response to inactivated and live, attenuated influenza vaccines. Scan. J. Immunol. 2004, 59, 1–15. [Google Scholar] [CrossRef]
- Capua, I.; Marangon, S. Control of avian influenza in poultry. Emerg. Infect. Dis. 2006, 12, 1319–1324. [Google Scholar] [CrossRef]
- Topp, E.; Irwin, R.; McAllister, T.; Lessard, M.; Joensuu, J.J.; Kolotilin, I.; Conrad, U.; Stöger, E.; Mor, T.; Warzecha, H.; et al. The case for plant-made veterinary immunotherapeutics. Biotechnol. Adv. 2016, 34, 597–604. [Google Scholar] [CrossRef] [PubMed]
- Treanor, J.J.; Wilkinson, B.E.; Masseoud, F.; Hu-Primmer, J.; Battaglia, R.; O’Brien, D.; Wolff, M.; Rabinovich, G.; Blackwelder, W.; Katz, J.M. Safety and immunogenicity of a recombinant hemagglutinin vaccine for H5 influenza in humans. Vaccine 2011, 19, 1732–1737. [Google Scholar] [CrossRef]
- Wei, C.J.; Xu, L.; Kong, W.P.; Shi, W.; Canis, K.; Stevens, J.; Yang, Z.Y.; Dell, A.; Haslam, S.M.; Wilson, I.A.; et al. Comparative Efficacy of Neutralizing Antibodies Elicited by Recombinant Hemagglutinin Proteins from Avian H5N1 Influenza Virus. J. Virol. 2008, 82, 6200–6208. [Google Scholar] [CrossRef] [PubMed]
- Khurana, S.; Verma, S.; Verma, N.; Crevar, C.J.; Carter, D.M.; Manischewitz, J.; King, L.R.; Ross, T.M.; Golding, H. Bacterial HA1 vaccine against pandemic H5N1 influenza virus: Evidence of oligomerization, hemagglutination, and cross-protective immunity in ferrets. J. Virol. 2011, 85, 1246–1256. [Google Scholar] [CrossRef] [PubMed]
- Phan, H.T.; Ho, T.T.; Chu, H.H.; Vu, T.H.; Gresch, U.; Conrad, U. Neutralizing immune responses induced by oligomeric H5N1-hemagglutinins from plants. Vet. Res. 2017, 48, 53. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Phan, H.T.; Gresch, U.; Conrad, U. In vitro-Formulated Oligomers of Strep-Tagged Avian Influenza Haemagglutinin Produced in Plants Cause Neutralizing Immune Responses. Front. Bioeng. Biotechnol. 2018, 6, 115. [Google Scholar] [CrossRef] [PubMed]
- Phan, H.T.; Pohl, J.; Floss, D.M.; Rabenstein, F.; Veits, J.; Le, B.T.; Chu, H.H.; Hause, G.; Mettenleiter, T.; Conrad, U. ELPylated haemagglutinins produced in tobacco plants induce potentially neutralizing antibodies against H5N1 viruses in mice. Plant Biotechnol. J. 2013, 11, 582–593. [Google Scholar] [CrossRef]
- Wandelt, C.I.; Khan, M.R.; Craig, S.; Schroeder, H.E.; Spencer, D.; Higgins, T.J. Vicilin with carboxy-terminal KDEL is retained in the endoplasmic reticulum and accumulates to high levels in the leaves of transgenic plants. Plant J. 1992, 2, 181–192. [Google Scholar]
- Müller, R.; Gräwert, M.A.; Kern, T.; Madl, T.; Peschek, J.; Sattler, M.; Groll, M.; Buchner, J. High-resolution structures of the IgM Fc domains reveal principles of its hexamer formation. Proc. Natl. Acad. Sci. USA 2013, 110, 10183–10188. [Google Scholar] [CrossRef]
- Kahsai, M.A.; Vogler, B.; Clark, A.T.; Edmondson, S.P.; Shriver, J.W. Solution structure, stability, and flexibility of Sso10a: A hyperthermophile coiled-coil DNA-binding protein. Biochemistry 2005, 44, 2822–2832. [Google Scholar] [CrossRef]
- Lu, Q.; Ye, F.; Wei, Z.; Wen, Z.; Zhang, M. Antiparallel coiled-coil–mediated dimerization of myosin X. Proc. Natl. Acad. Sci. USA 2012, 109, 17388–17393. [Google Scholar] [CrossRef] [PubMed]
- Hwang, E.; Cheong, H.K.; Mushtaq, A.U.; Kim, H.Y.; Yeo, K.J.; Kim, E.; Lee, W.C.; Hwang, K.Y.; Cheong, C.; Jeon, Y.H. Structural basis of the heterodimerization of the MST and RASSF SARAH domains in the Hippo signalling pathway. Acta Crystallogr. Sect. D Biol. Crystallogr. 2014, 70, 1944–1953. [Google Scholar] [CrossRef]
- Meir, A.; Bayer, E.A.; Livnah, O. Structural adaptation of a thermostable biotin-binding protein in a psychrophilic environment. J. Biol. Chem. 2012, 287, 17951–17962. [Google Scholar] [CrossRef]
- Avraham, O.; Meir, A.; Fish, A.; Bayer, E.A.; Livnah, O. Hoefavidin: A dimeric bacterial avidin with a C-terminal binding tail. J. Struct. Biol. 2015, 191, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Phan, H.; Conrad, U. Plant-Based Vaccine Antigen Production. In Vaccine Technologies for Veterinary Viral Diseases; Brun, A., Ed.; Springer: New York, NY, USA, 2016; Volume 1349, pp. 35–47. [Google Scholar]
- Conrad, U.; Plagmann, I.; Malchow, S.; Sack, M.; Floss, D.M.; Kruglov, A.A.; Nedospasov, S.A.; Rose-John, S.; Scheller, J. ELPylated anti-human TNF therapeutic single-domain antibodies for prevention of lethal septic shock. Plant Biotechnol. J. 2011, 9, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Gahrtz, M.; Conrad, U. Immunomodulation of plant function by in vitro selected single-chain Fv intrabodies. Methods Mol. Biol. 2009, 483, 289–312. [Google Scholar] [PubMed]
- Benoit, A.; Beran, J.; Devaster, J.M.; Esen, M.; Launay, O.; Leroux-Roels, G.; McElhaney, J.E.; Oostvogels, L.; van Essen, G.A.; Gaglani, M.; et al. Hemagglutination Inhibition Antibody Titers as a Correlate of Protection Against Seasonal A/H3N2 Influenza Disease. Open Forum Infect. Dis. 2015, 2, ofv067. [Google Scholar] [CrossRef]
- Hsu, J.P.; Zhao, X.; Mark, I.; Chen, C.; Cook, A.R.; Lee, V.; Lim, W.Y.; Tan, L.; Barr, I.G.; Jiang, L.; et al. Rate of decline of antibody titers to pandemic influenza A (H1N1-2009) by hemagglutination inhibition and virus microneutralization assays in a cohort of seroconverting adults in Singapore. BMC Infect. Dis. 2014, 14, 414. [Google Scholar] [CrossRef]
- Le, T.H.; Nguyen, N.T. Evolutionary dynamics of highly pathogenic avian influenza A/H5N1 HA clades and vaccine implementation in Vietnam. Clin. Exp. Vaccine Res. 2014, 3, 117–127. [Google Scholar] [CrossRef]
- Kanekiyo, M.; Wei, C.J.; Yassine, H.M.; McTamney, P.M.; Boyington, J.C.; Whittle, J.R.; Rao, S.S.; Kong, W.P.; Wang, L.; Nabel, G.J. Self-assembling influenza nanoparticle vaccines elicit broadly neutralizing H1N1 antibodies. Nature 2013, 499, 102–106. [Google Scholar] [CrossRef]
- Han, J.; Zhao, D.; Li, D.; Wang, X.; Jin, Z.; Zhao, K. Polymer-Based Nanomaterials and Applications for Vaccines and Drugs. Polymers 2018, 10, 31. [Google Scholar] [CrossRef]
- Kanekiyo, M.; Bu, W.; Joyce, M.G.; Meng, G.; Whittle, J.R.; Baxa, U.; Yamamoto, T.; Narpala, S.; Todd, J.P.; Rao, S.S.; et al. Rational Design of an Epstein-Barr Virus Vaccine Targeting the Receptor-Binding Site. Cell 2015, 162, 1090–1100. [Google Scholar] [CrossRef]
- Schmidt, T. Peptides with Sequentially Arranged Streptavidin Binding Modules. U.S. Patent No. 8,735,540, 27 May 2014. [Google Scholar]
- US Patent for IBA´s One-STrEP-tag Granted. Available online: https://www.iba-lifesciences.com/newsdetails/items/us-patent-for-ibas-one-strep-tag-granted.html (accessed on 6 October 2020).
- Phan, H.; Hause, B.; Hause, G.; Arcalis, E.; Stoger, E.; Maresch, D.; Altmann, F.; Joensuu, J.; Conrad, U. Influence of Elastin-Like Polypeptide and Hydrophobinon Recombinant Hemagglutinin Accumulations inTransgenic Tobacco Plants. PLoS ONE 2014, 9, e99347. [Google Scholar] [CrossRef]
- Zhang, S.; Sherwood, R.W.; Yang, Y.; Fish, T.; Chen, W.; McCardle, J.A.; Jones, R.M.; Yusibov, V.; May, E.R.; Rose, J.K.; et al. Comparative Characterization of the Glycosylation Profiles of an Influenza Hemagglutinin Produced in Plant and Insect Hosts. Proteomics 2012, 12, 1269–1288. [Google Scholar] [CrossRef] [PubMed]
- De Vries, R.P.; Cornelis, H.S.; de Bruin, E.; Rigter, A.; de Vries, E.; Cornelissen, L.A.; Eggink, D.; Chung, N.P.; Moore, J.P.; Sanders, R.W.; et al. Glycan-Dependent Immunogenicity of Recombinant Soluble Trimeric Hemagglutinin. J. Virol. 2012, 86, 11735–11744. [Google Scholar] [CrossRef] [PubMed]
- Landry, N.; Ward, B.J.; Trépanier, S.; Montomoli, E.; Dargis, M.; Lapini, G.; Vézina, L.P. Preclinical and clinical development of plant-made virus-like Particle vaccine against avian H5N1 Influenza. PLoS ONE 2010, 5, e15559. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, V.; Baztarrica, J.; Rybicki, E.P.; Meyers, A.E.; Wigdorovitz, A. Minimally processed crude leaf extracts of Nicotiana benthamiana containing recombinant foot and mouth disease virus-like particles are immunogenic in mice. Biotechnol. Rep. 2018, 20, e00283. [Google Scholar] [CrossRef]
- Stephan, A.; Hahn-Lobmann, S.; Rosche, F.; Buchholz, M.; Giritch, A.; Gleba, Y. Simple Purification of Nicotiana benthamiana-Produced Recombinant Colicins: High-Yield Recovery of Purified Proteins with Minimum Alkaloid Content Supports the Suitability of the Host for Manufacturing Food Additives. Int. J. Mol. Sci. 2017, 19, 95. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Seroconversion (One Day Old) | Seroconversion * (Before the Viral Challenge) |
|---|---|---|
| Purified H5 trimer | 0/7 | 5/5 |
| Non vaccinated | 0/7 | 0/5 |
| PBS | 0/7 | 0/5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Phan, H.T.; Pham, V.T.; Ho, T.T.; Pham, N.B.; Chu, H.H.; Vu, T.H.; Abdelwhab, E.M.; Scheibner, D.; Mettenleiter, T.C.; Hanh, T.X.; et al. Immunization with Plant-Derived Multimeric H5 Hemagglutinins Protect Chicken against Highly Pathogenic Avian Influenza Virus H5N1. Vaccines 2020, 8, 593. https://doi.org/10.3390/vaccines8040593
Phan HT, Pham VT, Ho TT, Pham NB, Chu HH, Vu TH, Abdelwhab EM, Scheibner D, Mettenleiter TC, Hanh TX, et al. Immunization with Plant-Derived Multimeric H5 Hemagglutinins Protect Chicken against Highly Pathogenic Avian Influenza Virus H5N1. Vaccines. 2020; 8(4):593. https://doi.org/10.3390/vaccines8040593
Chicago/Turabian StylePhan, Hoang Trong, Van Thi Pham, Thuong Thi Ho, Ngoc Bich Pham, Ha Hoang Chu, Trang Huyen Vu, Elsayed M. Abdelwhab, David Scheibner, Thomas C. Mettenleiter, Tran Xuan Hanh, and et al. 2020. "Immunization with Plant-Derived Multimeric H5 Hemagglutinins Protect Chicken against Highly Pathogenic Avian Influenza Virus H5N1" Vaccines 8, no. 4: 593. https://doi.org/10.3390/vaccines8040593
APA StylePhan, H. T., Pham, V. T., Ho, T. T., Pham, N. B., Chu, H. H., Vu, T. H., Abdelwhab, E. M., Scheibner, D., Mettenleiter, T. C., Hanh, T. X., Meister, A., Gresch, U., & Conrad, U. (2020). Immunization with Plant-Derived Multimeric H5 Hemagglutinins Protect Chicken against Highly Pathogenic Avian Influenza Virus H5N1. Vaccines, 8(4), 593. https://doi.org/10.3390/vaccines8040593