Evaluation of Glycosylated FlpA and SodB as Subunit Vaccines Against Campylobacter jejuni Colonisation in Chickens

 , and
, and 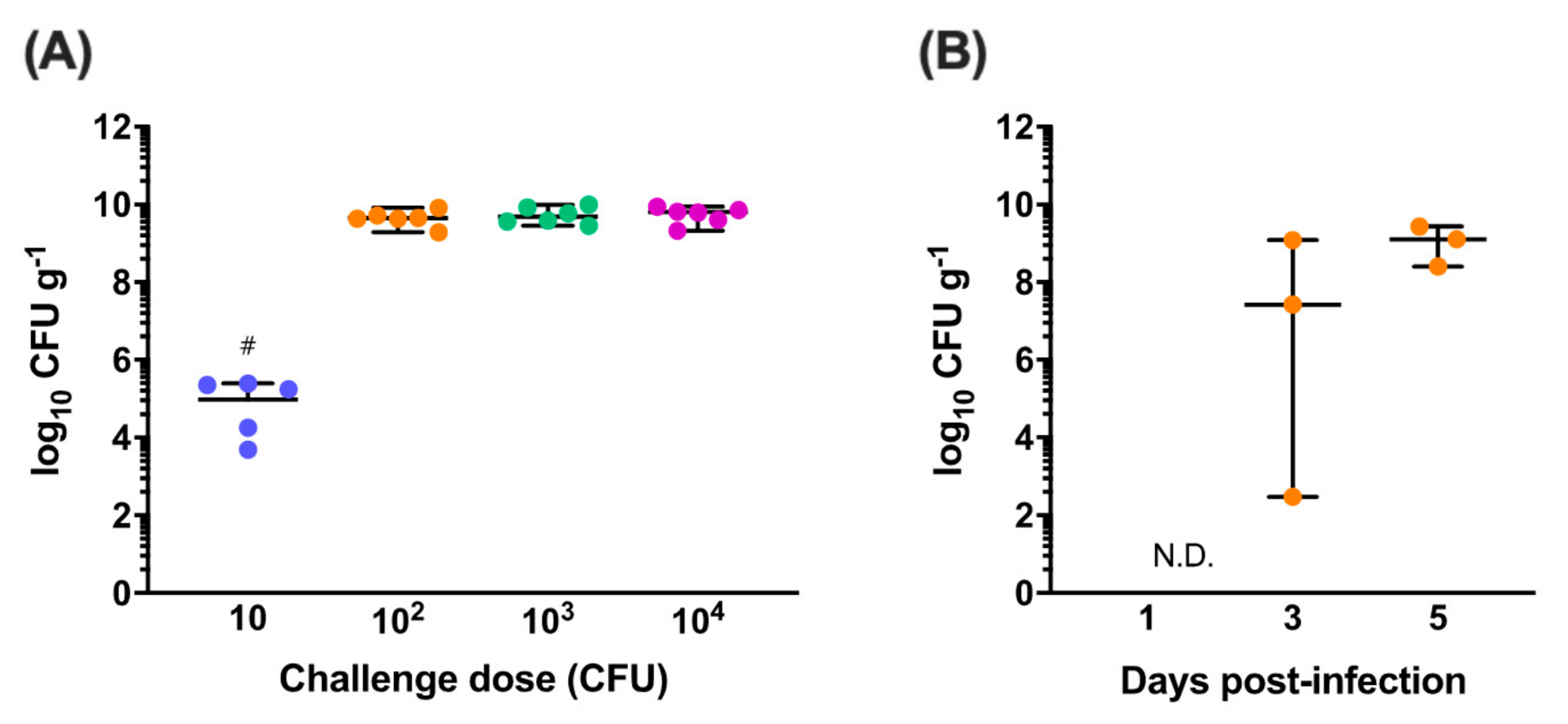{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
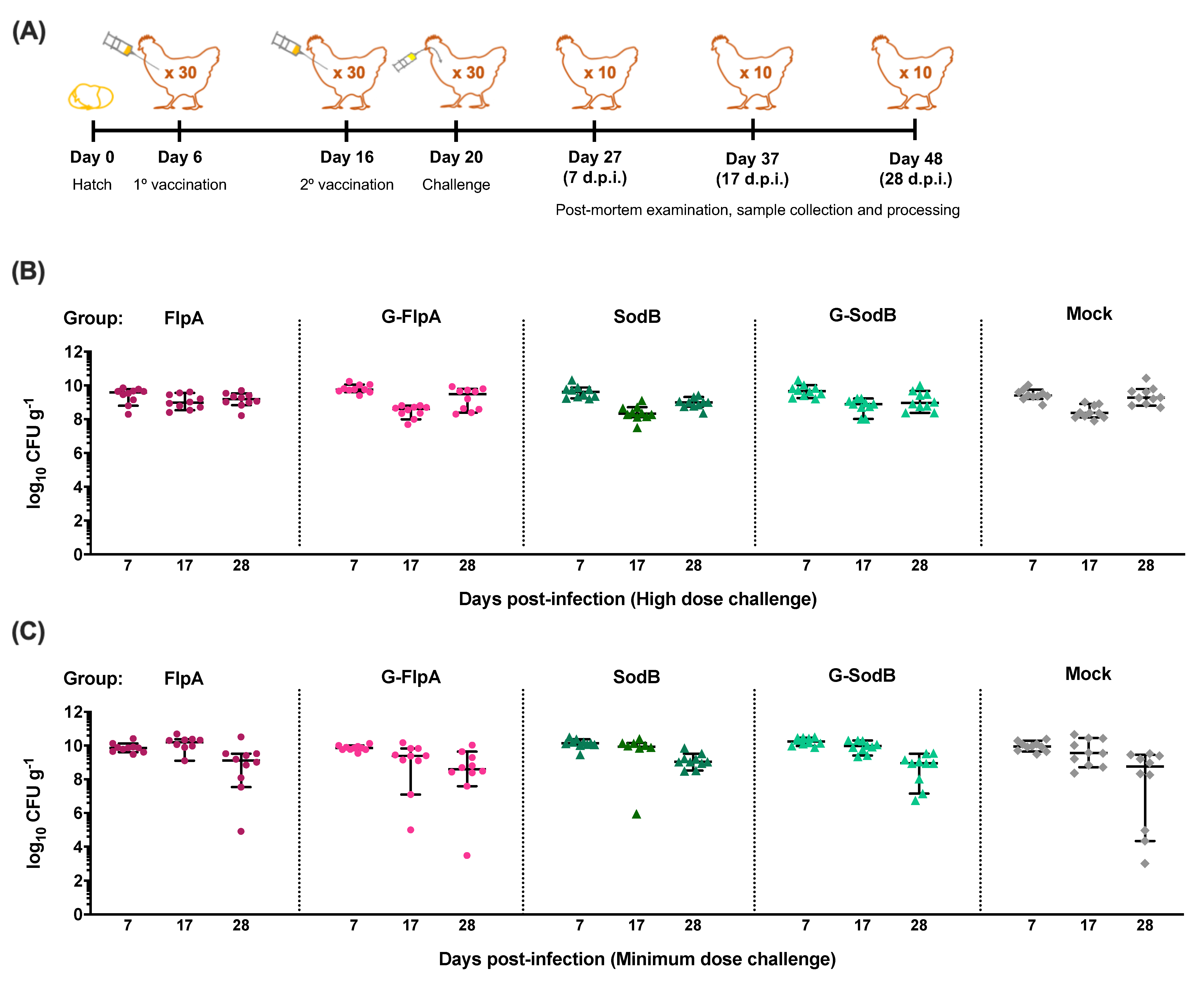
2.2. Animal Experiments with Layer Chickens
2.3. Dose Titration and Colonisation Dynamics of C. jejuni M1
2.4. Preparation of FlpA, SodB and Their Glycosylated Variants
2.5. Trials to Evaluate the Efficacy of FlpA, SodB and Their Glycosylated Variants as C. jejuni Vaccines
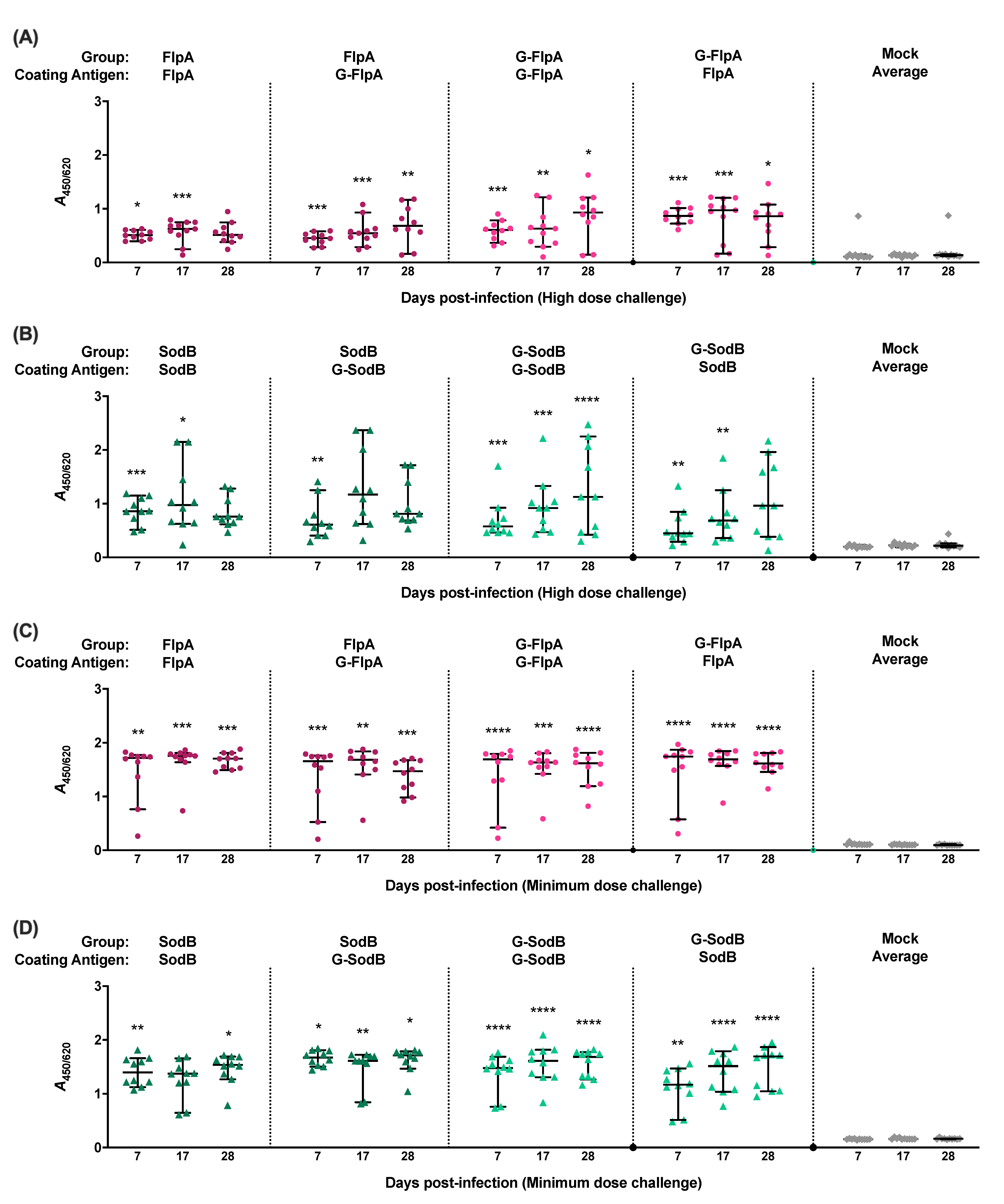
2.6. Analysis of Humoral Immune Responses Following Vaccination
2.7. Statistical Analysis
3. Results
3.1. Determining the Minimum Dose of C. jejuni M1 Required for Reliable Intestinal Colonisation
3.2. Effect of Vaccination on C. jejuni Colonisation
3.3. Immune Responses to Vaccination
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Havelaar, A.H.; Kirk, M.D.; Torgerson, P.R.; Gibb, H.J.; Hald, T.; Lake, R.J.; Praet, N.; Bellinger, D.C.; Silva, N.R.d.; Gargouri, N.; et al. World Health Organization foodborne disease burden epidemiology reference group world health organization global estimates and regional comparisons of the burden of foodborne disease in 2010. PLoS Med. 2015, 12, e1001923. [Google Scholar] [CrossRef] [PubMed]
- Tam, C.C.; Higgins, C.D.; Neal, K.R.; Rodrigues, L.C.; Millership, S.E.; O’Brien, S.J. Campylobacter Case-Control Study Group Chicken consumption and use of acid-suppressing medications as risk factors for Campylobacter enteritis, England. Emerg. Infect. Dis. 2009, 15, 1402–1408. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Panel on Biological Hazards (BIOHAZ) Scientific Opinion on Campylobacter in broiler meat production: Control options and performance objectives and/or targets at different stages of the food chain. EFSA J. 2011, 9, 2105. [Google Scholar] [CrossRef]
- Public Health England, London, United Kingdom. Zoonoses Report UK 2017. Available online: https://www.gov.uk/government/publications/zoonoses-uk-annual-reports (accessed on 10 September 2020).
- Tam, C.C.; Rodrigues, L.C.; Viviani, L.; Dodds, J.P.; Evans, M.R.; Hunter, P.R.; Gray, J.J.; Letley, L.H.; Rait, G.; Tompkins, D.S.; et al. IID2 Study Executive Committee Longitudinal study of infectious intestinal disease in the UK (IID2 study): Incidence in the community and presenting to general practice. Gut 2012, 61, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Tam, C.C.; O’Brien, S.J. Economic cost of Campylobacter, Norovirus and Rotavirus disease in the United Kingdom. PLoS ONE 2016, 11, e0138526. [Google Scholar] [CrossRef]
- Food Standards Agency, United Kingdom. A microbiological Survey of Campylobacter Contamination in Fresh Whole UK-Produced Chilled Chickens at Retail Sale (Y2/3/4). Available online: https://www.food.gov.uk/print/pdf/node/680 (accessed on 10 September 2020).
- Nachamkin, I.; Allos, B.M.; Ho, T. Campylobacter species and Guillain-Barré syndrome. Clin. Microbiol. Rev. 1998, 11, 555–567. [Google Scholar] [CrossRef]
- McCarthy, N.; Giesecke, J. Incidence of Guillain-Barré syndrome following infection with Campylobacter jejuni. Am. J. Epidemiol. 2001, 153, 610–614. [Google Scholar] [CrossRef]
- Nachamkin, I. Chronic effects of Campylobacter infection. Microbes Infect. 2002, 4, 399–403. [Google Scholar] [CrossRef]
- Pope, J.E.; Krizova, A.; Garg, A.X.; Thiessen-Philbrook, H.; Ouimet, J.M. Campylobacter reactive arthritis: A systematic review. Semin. Arthritis Rheum. 2007, 37, 48–55. [Google Scholar] [CrossRef]
- Beery, J.T.; Hugdahl, M.B.; Doyle, M.P. Colonization of gastrointestinal tracts of chicks by Campylobacter jejuni. Appl. Environ. Microbiol. 1988, 54, 2365–2370. [Google Scholar] [CrossRef]
- Sahin, O.; Morishita, T.Y.; Zhang, Q. Campylobacter colonization in poultry: Sources of infection and modes of transmission. Anim. Health Res. Rev. 2002, 3, 95–105. [Google Scholar] [CrossRef]
- Awad, W.A.; Molnár, A.; Aschenbach, J.R.; Ghareeb, K.; Khayal, B.; Hess, C.; Liebhart, D.; Dublecz, K.; Hess, M. Campylobacter infection in chickens modulates the intestinal epithelial barrier function. Innate Immun. 2014, 21, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, S.; Chaloner, G.; Kemmett, K.; Davidson, N.; Williams, N.; Kipar, A.; Humphrey, T.; Wigley, P. Campylobacter jejuni is not merely a commensal in commercial broiler chickens and affects bird welfare. mBio 2014, 5, e01364-14. [Google Scholar] [CrossRef] [PubMed]
- Rosenquist, H.; Nielsen, N.L.; Sommer, H.M.; Nørrung, B.; Christensen, B.B. Quantitative risk assessment of human campylobacteriosis associated with thermophilic Campylobacter species in chickens. Int. J. Food Microbiol. 2003, 83, 87–103. [Google Scholar] [CrossRef]
- Khoury, C.A.; Meinersmann, R.J. A genetic hybrid of the Campylobacter jejuni flaA gene with LT-B of Escherichia coli and assessment of the efficacy of the hybrid protein as an oral chicken vaccine. Avian Dis. 1995, 39, 812–820. [Google Scholar] [CrossRef]
- Neal-McKinney, J.M.; Samuelson, D.R.; Eucker, T.P.; Nissen, M.S.; Crespo, R.; Konkel, M.E. Reducing Campylobacter jejuni colonization of poultry via vaccination. PLoS ONE 2014, 9, e114254. [Google Scholar] [CrossRef]
- Chintoan-Uta, C.; Cassady-Cain, R.L.; Stevens, M.P. Evaluation of flagellum-related proteins FliD and FspA as subunit vaccines against Campylobacter jejuni colonisation in chickens. Vaccine 2016, 34, 1739–1743. [Google Scholar] [CrossRef]
- Wyszyńska, A.; Raczko, A.; Lis, M.; Jagusztyn-Krynicka, E.K. Oral immunization of chickens with avirulent Salmonella vaccine strain carrying C. jejuni 72Dz/92 cjaA gene elicits specific humoral immune response associated with protection against challenge with wild-type Campylobacter. Vaccine 2004, 22, 1379–1389. [Google Scholar] [CrossRef]
- Buckley, A.M.; Wang, J.; Hudson, D.L.; Grant, A.J.; Jones, M.A.; Maskell, D.J.; Stevens, M.P. Evaluation of live-attenuated Salmonella vaccines expressing Campylobacter antigens for control of C. jejuni in poultry. Vaccine 2010, 28, 1094–1105. [Google Scholar] [CrossRef]
- Layton, S.L.; Morgan, M.J.; Cole, K.; Kwon, Y.M.; Donoghue, D.J.; Hargis, B.M.; Pumford, N.R. Evaluation of Salmonella-vectored Campylobacter peptide epitopes for reduction of Campylobacter jejuni in broiler chickens. Clin. Vaccine Immunol. 2011, 18, 449–454. [Google Scholar] [CrossRef]
- Clark, J.D.; Oakes, R.D.; Redhead, K.; Crouch, C.F.; Francis, M.J.; Tomley, F.M.; Blake, D.P. Eimeria species parasites as novel vaccine delivery vectors: Anti-Campylobacter jejuni protective immunity induced by Eimeria tenella-delivered CjaA. Vaccine 2012, 30, 2683–2688. [Google Scholar] [CrossRef] [PubMed]
- Łaniewski, P.; Kuczkowski, M.; Chrząstek, K.; Woźniak, A.; Wyszyńska, A.; Wieliczko, A.; Jagusztyn-Krynicka, E.K. Evaluation of the immunogenicity of Campylobacter jejuni CjaA protein delivered by Salmonella enterica sv. Typhimurium strain with regulated delayed attenuation in chickens. World J. Microbiol. Biotechnol. 2014, 30, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Chintoan-Uta, C.; Cassady-Cain, R.L.; Al-Haideri, H.; Watson, E.; Kelly, D.J.; Smith, D.G.E.; Sparks, N.H.C.; Kaiser, P.; Stevens, M.P. Superoxide dismutase SodB is a protective antigen against Campylobacter jejuni colonisation in chickens. Vaccine 2015, 33, 6206–6211. [Google Scholar] [CrossRef][Green Version]
- Guerry, P.; Poly, F.; Riddle, M.; Maue, A.C.; Chen, Y.-H.; Monteiro, M.A. Campylobacter polysaccharide capsules: Virulence and vaccines. Front. Cell Infect. Microbiol. 2012, 2, 7. [Google Scholar] [CrossRef]
- Bertolo, L.; Ewing, C.P.; Maue, A.; Poly, F.; Guerry, P.; Monteiro, M.A. The design of a capsule polysaccharide conjugate vaccine against Campylobacter jejuni serotype HS15. Carbohydr. Res. 2013, 366, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Hodgins, D.C.; Barjesteh, N.; Paul, M.S.; Ma, Z.; Monteiro, M.A.; Sharif, S. Evaluation of a polysaccharide conjugate vaccine to reduce colonization by Campylobacter jejuni in broiler chickens. BMC Res. Notes 2015, 8, 204–208. [Google Scholar] [CrossRef] [PubMed]
- Nothaft, H.; Davis, B.; Lock, Y.Y.; Perez-Munoz, M.E.; Vinogradov, E.; Walter, J.; Coros, C.; Szymanski, C.M. Engineering the Campylobacter jejuni N-glycan to create an effective chicken vaccine. Sci. Rep. 2016, 6, 26511. [Google Scholar] [CrossRef]
- Price, N.L.; Goyette-Desjardins, G.; Nothaft, H.; Valguarnera, E.; Szymanski, C.M.; Segura, M.; Feldman, M.F. Glycoengineered outer membrane vesicles: A novel platform for bacterial vaccines. Sci. Rep. 2016, 6, 24931. [Google Scholar] [CrossRef]
- Nothaft, H.; Perez-Muñoz, M.E.; Gouveia, G.J.; Duar, R.M.; Wanford, J.J.; Lango-Scholey, L.; Panagos, C.G.; Srithayakumar, V.; Plastow, G.S.; Coros, C.; et al. Co-administration of the Campylobacter jejuni N-glycan based vaccine with probiotics improves vaccine performance in broiler chickens. Appl. Environ. Microbiol. 2017, 83, e01523-17. [Google Scholar] [CrossRef]
- Kowarik, M.; Young, N.M.; Numao, S.; Schulz, B.L.; Hug, I.; Callewaert, N.; Mills, D.C.; Watson, D.C.; Hernandez, M.; Kelly, J.F.; et al. Definition of the bacterial N-glycosylation site consensus sequence. EMBO J. 2006, 25, 1957–1966. [Google Scholar] [CrossRef]
- Karlyshev, A.V.; Everest, P.; Linton, D.; Cawthraw, S.; Newell, D.G.; Wren, B.W. The Campylobacter jejuni general glycosylation system is important for attachment to human epithelial cells and in the colonization of chicks. Microbiology 2004, 150, 1957–1964. [Google Scholar] [CrossRef]
- Jones, M.A.; Marston, K.L.; Woodall, C.A.; Maskell, D.J.; Linton, D.; Karlyshev, A.V.; Dorrell, N.; Wren, B.W.; Barrow, P.A. Adaptation of Campylobacter jejuni NCTC11168 to high-level colonization of the avian gastrointestinal tract. Infect. Immun. 2004, 72, 3769–3776. [Google Scholar] [CrossRef] [PubMed]
- Kelly, J.; Jarrell, H.; Millar, L.; Tessier, L.; Fiori, L.M.; Lau, P.C.; Allan, B.; Szymanski, C.M. Biosynthesis of the N-linked glycan in Campylobacter jejuni and addition onto protein through block transfer. J. Bacteriol. 2006, 188, 2427–2434. [Google Scholar] [CrossRef] [PubMed]
- Abouelhadid, S.; North, S.J.; Hitchen, P.; Vohra, P.; Chintoan-Uta, C.; Stevens, M.; Dell, A.; Cuccui, J.; Wren, B.W. Quantitative analyses reveal novel roles for N-glycosylation in a major enteric bacterial pathogen. mBio 2019, 10, e00297-19. [Google Scholar] [CrossRef] [PubMed]
- Cain, J.A.; Dale, A.L.; Niewold, P.; Klare, W.P.; Man, L.; White, M.Y.; Scott, N.E.; Cordwell, S.J. Proteomics reveals multiple phenotypes associated with n-linked glycosylation in Campylobacter jejuni. Mol. Cell Proteom. 2019, 18, 715–734. [Google Scholar] [CrossRef] [PubMed]
- Wacker, M.; Linton, D.; Hitchen, P.G.; Nita-Lazar, M.; Haslam, S.M.; North, S.J.; Panico, M.; Morris, H.R.; Dell, A.; Wren, B.W.; et al. N-linked glycosylation in Campylobacter jejuni and its functional transfer into E. coli. Science 2002, 298, 1790–1793. [Google Scholar] [CrossRef] [PubMed]
- Terra, V.S.; Mills, D.C.; Yates, L.E.; Abouelhadid, S.; Cuccui, J.; Wren, B.W. Recent developments in bacterial protein glycan coupling technology and glycoconjugate vaccine design. J. Med. Microbiol. 2012, 61, 919–926. [Google Scholar] [CrossRef]
- Cuccui, J.; Thomas, R.M.; Moule, M.G.; D’Elia, R.V.; Laws, T.R.; Mills, D.C.; Williamson, D.; Atkins, T.P.; Prior, J.L.; Wren, B.W. Exploitation of bacterial N-linked glycosylation to develop a novel recombinant glycoconjugate vaccine against Francisella tularensis. Open Biol. 2013, 3, 130002. [Google Scholar] [CrossRef][Green Version]
- Wacker, M.; Wang, L.; Kowarik, M.; Dowd, M.; Lipowsky, G.; Faridmoayer, A.; Shields, K.; Park, S.; Alaimo, C.; Kelley, K.A.; et al. Prevention of Staphylococcus aureus infections by glycoprotein vaccines synthesized in Escherichia coli. J. Infect. Dis. 2014, 209, 1551–1561. [Google Scholar] [CrossRef]
- Riddle, M.S.; Kaminski, R.W.; Di Paolo, C.; Porter, C.K.; Gutierrez, R.L.; Clarkson, K.A.; Weerts, H.E.; Duplessis, C.; Castellano, A.; Alaimo, C.; et al. Safety and immunogenicity of a candidate bioconjugate vaccine against Shigella flexneri 2a administered to healthy adults: A single-blind, randomized Phase I Study. Clin. Vaccine Immunol. 2016, 23, 908–917. [Google Scholar] [CrossRef]
- Reglinski, M.; Ercoli, G.; Plumptre, C.; Kay, E.; Petersen, F.C.; Paton, J.C.; Wren, B.W.; Brown, J.S. A recombinant conjugated pneumococcal vaccine that protects against murine infections with a similar efficacy to Prevnar-13. Npj Vaccines 2018, 3, 53. [Google Scholar] [CrossRef] [PubMed]
- Friis, C.; Wassenaar, T.M.; Javed, M.A.; Snipen, L.; Lagesen, K.; Hallin, P.F.; Newell, D.G.; Toszeghy, M.; Ridley, A.; Manning, G.; et al. Genomic characterization of Campylobacter jejuni strain M1. PLoS ONE 2010, 5, e12253. [Google Scholar] [CrossRef] [PubMed]
- Coward, C.; van Diemen, P.M.; Conlan, A.J.K.; Gog, J.R.; Stevens, M.P.; Jones, M.A.; Maskell, D.J. Competing isogenic Campylobacter strains exhibit variable population structures in vivo. Appl. Environ. Microbiol. 2008, 74, 3857–3867. [Google Scholar] [CrossRef] [PubMed]
- Conlan, A.J.K.; Line, J.E.; Hiett, K.; Coward, C.; van Diemen, P.M.; Stevens, M.P.; Jones, M.A.; Gog, J.R.; Maskell, D.J. Transmission and dose-response experiments for social animals: A reappraisal of the colonization biology of Campylobacter jejuni in chickens. J. R. Soc. Interface 2011, 8, 1720–1735. [Google Scholar] [CrossRef]
- Lacharme-Lora, L.; Chaloner, G.; Gilroy, R.; Humphrey, S.; Gibbs, K.; Jopson, S.; Wright, E.; Reid, W.; Ketley, J.; Humphrey, T.; et al. B lymphocytes play a limited role in clearance of Campylobacter jejuni from the chicken intestinal tract. Sci. Rep. 2017, 7, 45090. [Google Scholar] [CrossRef]
- Gilroy, R.; Chaloner, G.; Wedley, A.; Lacharme-Lora, L.; Jopson, S.; Wigley, P. Campylobacter jejuni transmission and colonisation in broiler chickens is inhibited by Faecal Microbiota Transplantation. bioRxiv 2018, 476119. [Google Scholar] [CrossRef]
- Pillai, S.; Dermody, K.; Metcalf, B. Immunogenicity of genetically engineered glutathione S-transferase fusion proteins containing a T-cell epitope from diphtheria toxin. Infect. Immun. 1995, 63, 1535–1540. [Google Scholar] [CrossRef]
- Rao, K.V.N.; He, Y.-X.; Kalyanasundaram, R. Expression of a 28-kilodalton glutathione S-transferase antigen of Schistosoma mansoni on the surface of filamentous phages and evaluation of its vaccine potential. Clin. Diagn. Lab. Immunol. 2003, 10, 536–541. [Google Scholar] [CrossRef]
- Taylor, D.N.; Trofa, A.C.; Sadoff, J.; Chu, C.; Bryla, D.; Shiloach, J.; Cohen, D.; Ashkenazi, S.; Lerman, Y.; Egan, W. Synthesis, characterization, and clinical evaluation of conjugate vaccines composed of the O-specific polysaccharides of Shigella dysenteriae type 1, Shigella flexneri type 2a, and Shigella sonnei (Plesiomonas shigelloides) bound to bacterial toxoids. Infect. Immun. 1993, 61, 3678–3687. [Google Scholar] [CrossRef]
- Ihssen, J.; Kowarik, M.; Dilettoso, S.; Tanner, C.; Wacker, M.; Thöny-Meyer, L. Production of glycoprotein vaccines in Escherichia coli. Microb. Cell Fact. 2010, 9, 13–61. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vohra, P.; Chintoan-Uta, C.; Terra, V.S.; Bremner, A.; Cuccui, J.; Wren, B.W.; Vervelde, L.; Stevens, M.P., on behalf of the Glycoengineering of Veterinary Vaccines Consortium. Evaluation of Glycosylated FlpA and SodB as Subunit Vaccines Against Campylobacter jejuni Colonisation in Chickens. Vaccines 2020, 8, 520. https://doi.org/10.3390/vaccines8030520
Vohra P, Chintoan-Uta C, Terra VS, Bremner A, Cuccui J, Wren BW, Vervelde L, Stevens MP on behalf of the Glycoengineering of Veterinary Vaccines Consortium. Evaluation of Glycosylated FlpA and SodB as Subunit Vaccines Against Campylobacter jejuni Colonisation in Chickens. Vaccines. 2020; 8(3):520. https://doi.org/10.3390/vaccines8030520
Chicago/Turabian StyleVohra, Prerna, Cosmin Chintoan-Uta, Vanessa S. Terra, Abi Bremner, Jon Cuccui, Brendan W. Wren, Lonneke Vervelde, and Mark P. Stevens on behalf of the Glycoengineering of Veterinary Vaccines Consortium. 2020. "Evaluation of Glycosylated FlpA and SodB as Subunit Vaccines Against Campylobacter jejuni Colonisation in Chickens" Vaccines 8, no. 3: 520. https://doi.org/10.3390/vaccines8030520
APA StyleVohra, P., Chintoan-Uta, C., Terra, V. S., Bremner, A., Cuccui, J., Wren, B. W., Vervelde, L., & Stevens, M. P., on behalf of the Glycoengineering of Veterinary Vaccines Consortium. (2020). Evaluation of Glycosylated FlpA and SodB as Subunit Vaccines Against Campylobacter jejuni Colonisation in Chickens. Vaccines, 8(3), 520. https://doi.org/10.3390/vaccines8030520