A Replicating Single-Cycle Adenovirus Vaccine Effective against Clostridium difficile

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Single-Cycle Adenoviruses
2.3. Western Blotting
2.4. Animals
2.5. Immunizations and Sample Collection
2.6. Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. Cytotoxicity and Neutralization Assays
2.8. Challenge with Recombinant C. difficile Toxin A in Mice
2.9. Hematology and Clinical Chemistry in Hamsters
2.10. Challenge with C. difficile Spores in Hamsters
2.11. Statistical Analysis
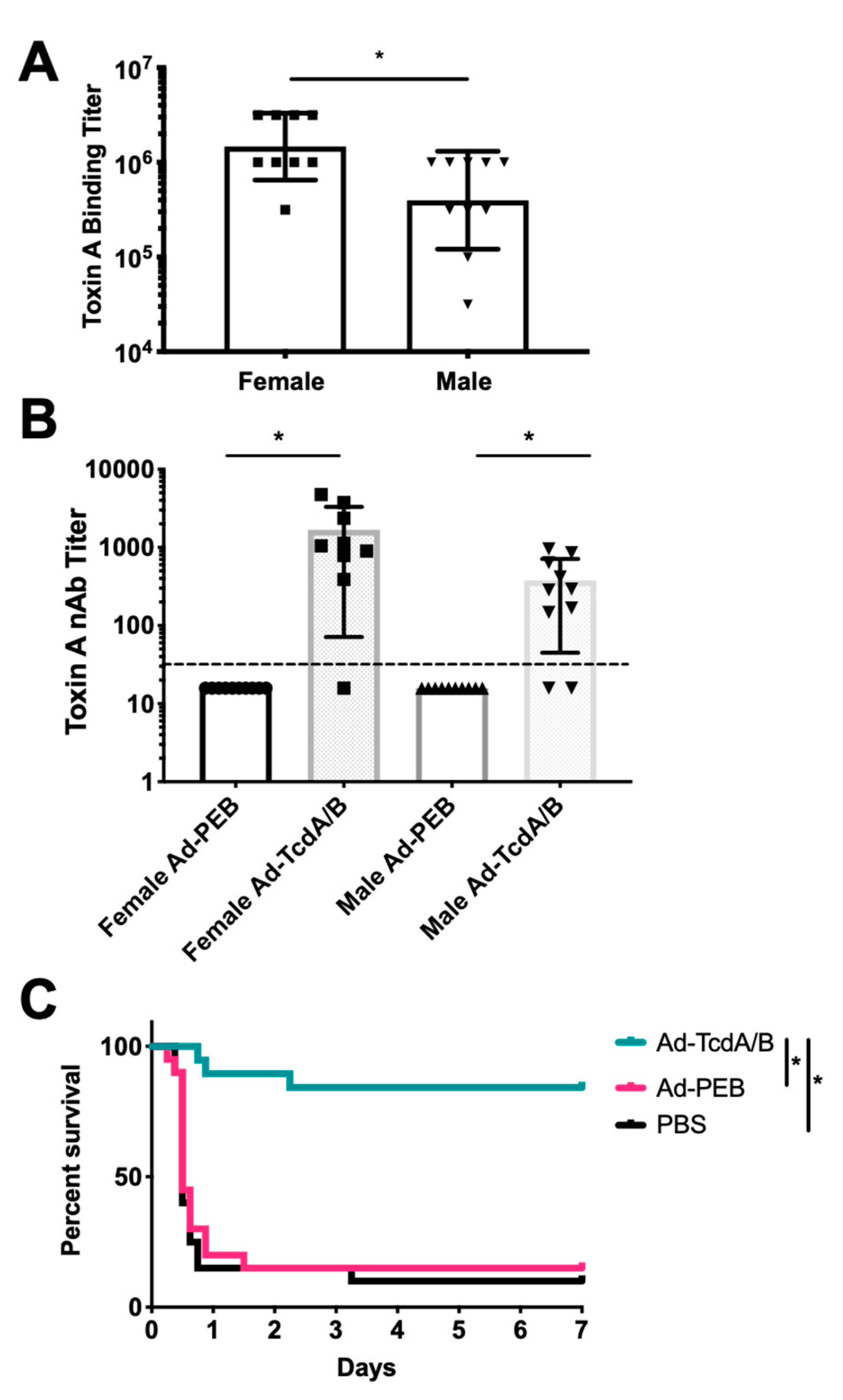
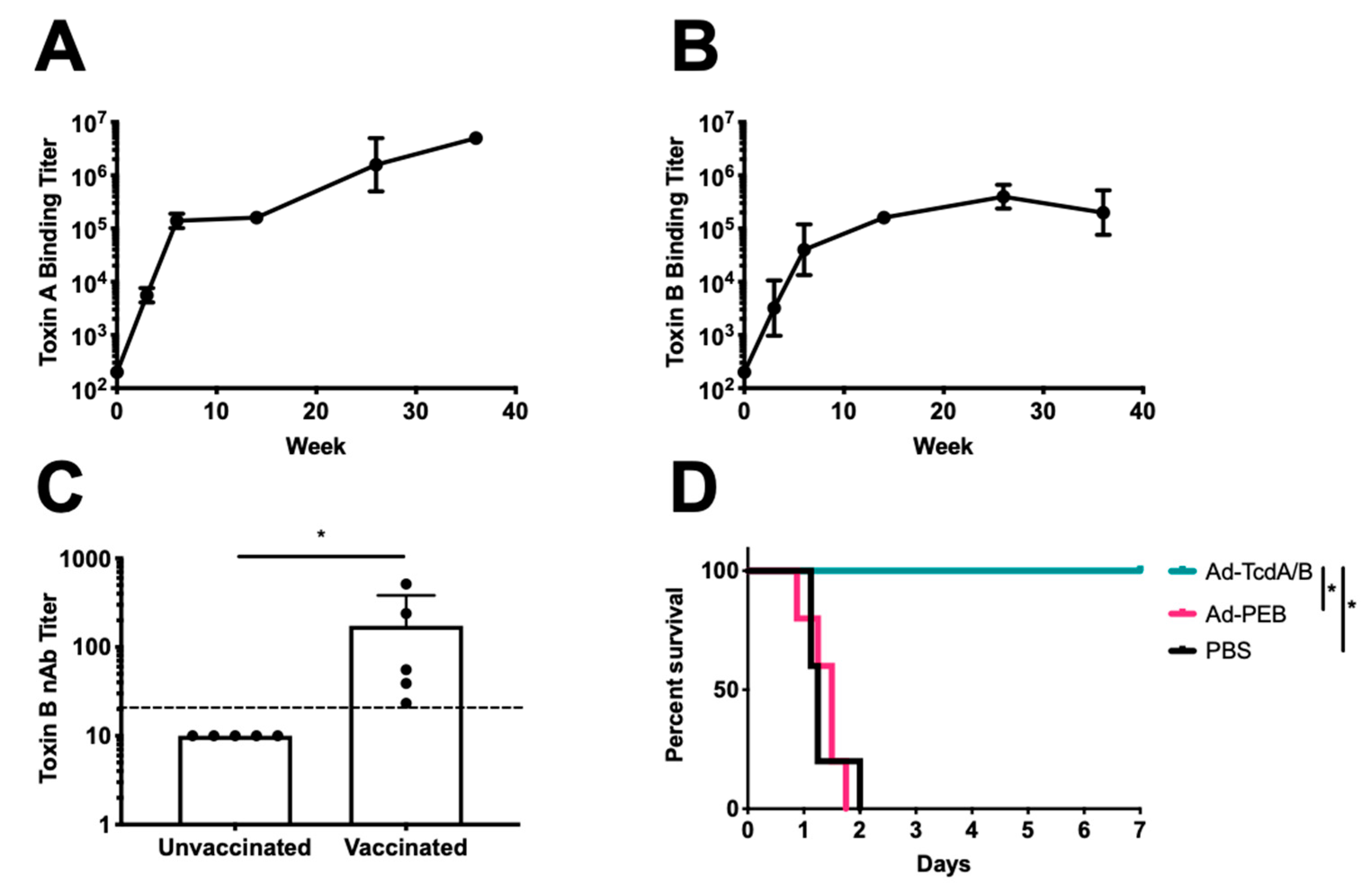
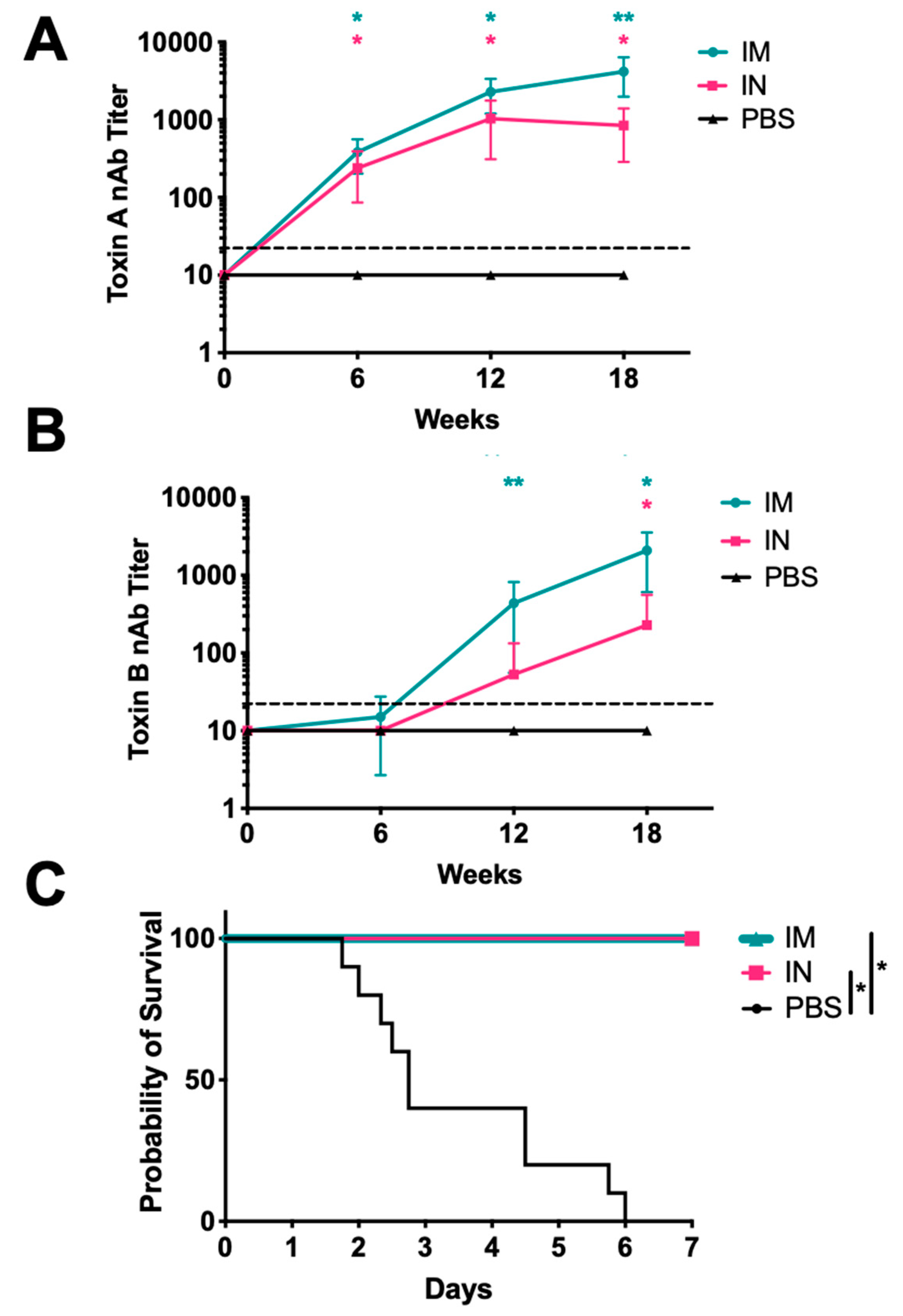
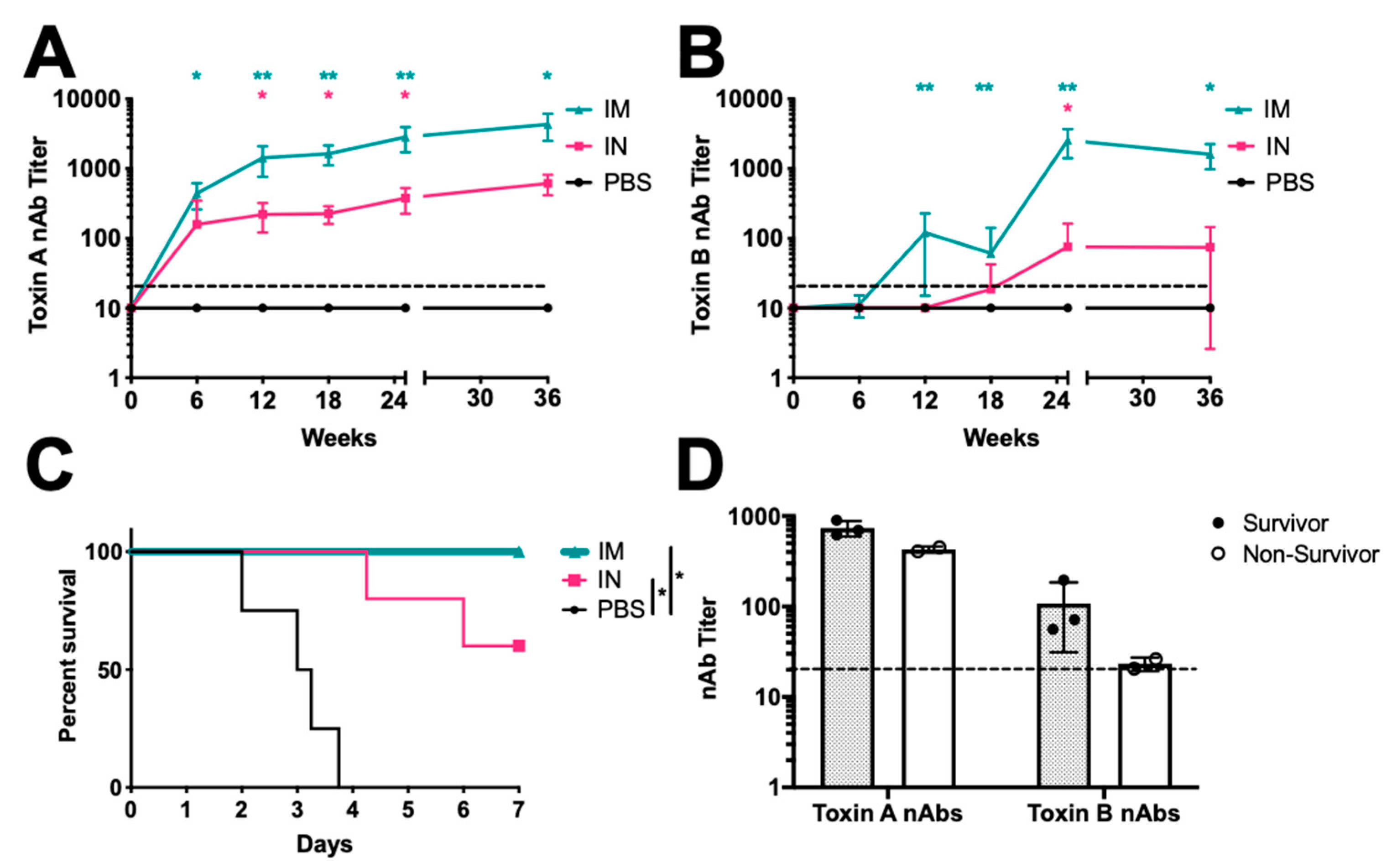
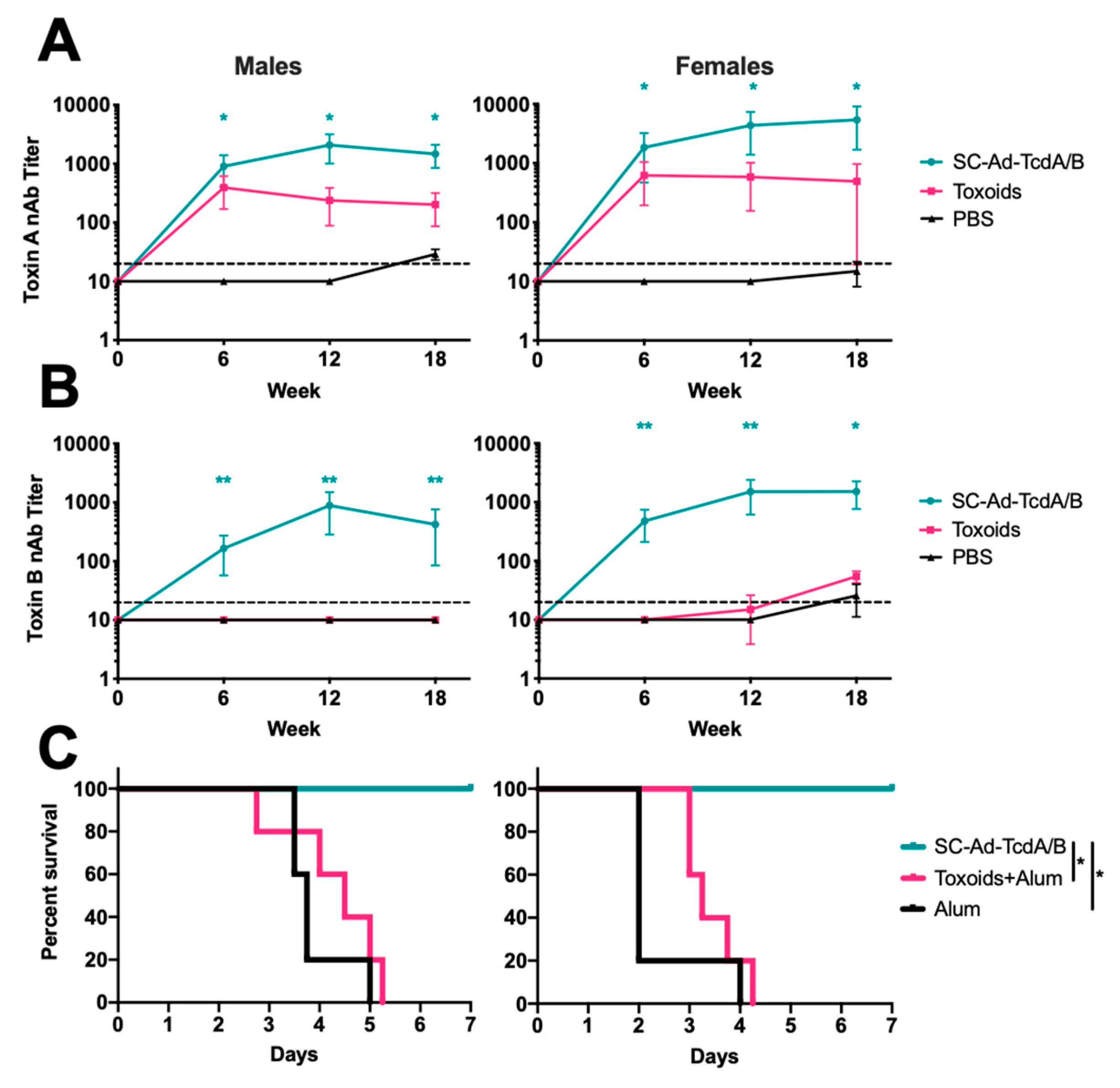
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Oren, A.; Rupnik, M. Clostridium difficile and Clostridioides difficile: Two validly published and correct names. Anaerobe 2018, 52, 125–126. [Google Scholar] [CrossRef]
- Leffler, D.A.; Lamont, J.T. Clostridium difficile infection. N. Engl. J. Med. 2015, 372, 1539–1548. [Google Scholar] [CrossRef] [PubMed]
- Lessa, F.C.; Mu, Y.; Bamberg, W.M.; Beldavs, Z.G.; Dumyati, G.K.; Dunn, J.R.; Farley, M.M.; Holzbauer, S.M.; Meek, J.I.; Phipps, E.C.; et al. Burden of Clostridium difficile Infection in the United States. N. Engl. J. Med. 2015, 372, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Surawicz, C.M.; Brandt, L.J.; Binion, D.G.; Ananthakrishnan, A.N.; Curry, S.R.; Gilligan, P.H.; McFarland, L.V.; Mellow, M.; Zuckerbraun, B.S. Guidelines for diagnosis, treatment, and prevention of Clostridium difficile infections. Am. J. Gastroenterol. 2013, 108, 478–498. [Google Scholar] [CrossRef] [PubMed]
- Kuipers, E.J.; Surawicz, C.M. Clostridium difficile infection. Lancet 2008, 371, 1486–1488. [Google Scholar] [CrossRef]
- McDonald, L.C.; Gerding, D.N.; Johnson, S.; Bakken, J.S.; Carroll, K.C.; Coffin, S.E.; Dubberke, E.R.; Garey, K.W.; Gould, C.V.; Kelly, C.; et al. Clinical Practice Guidelines for Clostridium difficile Infection in Adults and Children: 2017 Update by the Infectious Diseases Society of America (IDSA) and Society for Healthcare Epidemiology of America (SHEA). Clin. Infect. Dis. 2018, 66, e1–e48. [Google Scholar] [CrossRef] [PubMed]
- Catanzaro, A.T.; Koup, R.A.; Roederer, M.; Bailer, R.T.; Enama, M.E.; Moodie, Z.; Gu, L.; Martin, J.E.; Novik, L.; Chakrabarti, B.K.; et al. Phase 1 Safety and Immunogenicity Evaluation of a Multiclade HIV-1 Candidate Vaccine Delivered by a Replication-Defective Recombinant Adenovirus Vector. J. Infect. Dis. 2006, 194, 1638–1649. [Google Scholar] [CrossRef]
- Kink, J.A.; Williams, J.A. Antibodies to recombinant Clostridium difficile toxins A and B are an effective treatment and prevent relapse of C. difficile-associated disease in a hamster model of infection. Infect. Immun. 1998, 66, 2018–2025. [Google Scholar] [CrossRef]
- Babcock, G.J.; Broering, T.J.; Hernandez, H.J.; Mandell, R.B.; Donahue, K.; Boatright, N.; Stack, A.M.; Lowy, I.; Graziano, R.; Molrine, D.; et al. Human monoclonal antibodies directed against toxins A and B prevent Clostridium difficile-induced mortality in hamsters. Infect. Immun. 2006, 74, 6339–6347. [Google Scholar] [CrossRef]
- Kim, P.H.; Iaconis, J.P.; Rolfe, R.D. Immunization of adult hamsters against Clostridium difficile-associated ileocecitis and transfer of protection to infant hamsters. Infect. Immun. 1987, 55, 2984–2992. [Google Scholar] [CrossRef]
- Aboudola, S.; Kotloff, K.L.; Kyne, L.; Warny, M.; Kelly, E.C.; Sougioultzis, S.; Giannasca, P.J.; Monath, T.P.; Kelly, C.P. Clostridium difficile vaccine and serum immunoglobulin G antibody response to toxin A. Infect. Immun. 2003, 71, 1608–1610. [Google Scholar] [CrossRef] [PubMed]
- Carter, G.P.; Rood, J.I.; Lyras, D. The role of toxin A and toxin B in Clostridium difficile-associated disease: Past and present perspectives. Gut Microbes 2010, 1, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Aktories, K.; Schwan, C.; Jank, T. Clostridium difficile Toxin Biology. Annu. Rev. Microbiol. 2017, 71, 281–307. [Google Scholar] [CrossRef] [PubMed]
- Greco, A.; Ho, J.G.; Lin, S.J.; Palcic, M.M.; Rupnik, M.; Ng, K.K. Carbohydrate recognition by Clostridium difficile toxin A. Nat. Struct. Mol. Biol. 2006, 13, 460–461. [Google Scholar] [CrossRef] [PubMed]
- Spencer, J.; Leuzzi, R.; Buckley, A.; Irvine, J.; Candlish, D.; Scarselli, M.; Douce, G.R. Vaccination against Clostridium difficile using toxin fragments: Observations and analysis in animal models. Gut Microbes 2014, 5, 225–232. [Google Scholar] [CrossRef]
- Sheldon, E.; Kitchin, N.; Peng, Y.; Eiden, J.; Gruber, W.; Johnson, E.; Jansen, K.U.; Pride, M.W.; Pedneault, L. A phase 1, placebo-controlled, randomized study of the safety, tolerability, and immunogenicity of a Clostridium difficile vaccine administered with or without aluminum hydroxide in healthy adults. Vaccine 2016, 34, 2082–2091. [Google Scholar] [CrossRef] [PubMed]
- Henderson, M.; Bragg, A.; Fahim, G.; Shah, M.; Hermes-DeSantis, E.R. A Review of the Safety and Efficacy of Vaccines as Prophylaxis for Clostridium difficile Infections. Vaccines 2017, 5, 25. [Google Scholar] [CrossRef] [PubMed]
- Rees, W.D.; Steiner, T.S. Adaptive immune response to Clostridium difficile infection: A perspective for prevention and therapy. Eur. J. Immunol. 2018, 48, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Donald, R.G.; Flint, M.; Kalyan, N.; Johnson, E.; Witko, S.E.; Kotash, C.; Zhao, P.; Megati, S.; Yurgelonis, I.; Lee, P.K.; et al. A novel approach to generate a recombinant toxoid vaccine against Clostridium difficile. Microbiology 2013, 159, 1254–1266. [Google Scholar] [CrossRef]
- Lyerly, D.M.; Phelps, C.J.; Toth, J.; Wilkins, T.D. Characterization of toxins A and B of Clostridium difficile with monoclonal antibodies. Infect. Immun. 1986, 54, 70–76. [Google Scholar] [CrossRef]
- Chai, J.; Lee, C.H. Management of Primary and Recurrent Clostridium difficile Infection: An Update. Antibiotics 2018, 7, 54. [Google Scholar] [CrossRef] [PubMed]
- Crosby, C.M.; Barry, M.A. IIIa deleted adenovirus as a single-cycle genome replicating vector. Virology 2014, 462–463, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Crosby, C.M.; Barry, M.A. Transgene Expression and Host Cell Responses to Replication-Defective, Single-Cycle, and Replication-Competent Adenovirus Vectors. Genes 2017, 8, 79. [Google Scholar] [CrossRef]
- Crosby, C.M.; Matchett, W.E.; Anguiano-Zarate, S.S.; Parks, C.A.; Weaver, E.A.; Pease, L.R.; Webby, R.J.; Barry, M.A. Replicating Single-Cycle Adenovirus Vectors Generate Amplified Influenza Vaccine Responses. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed]
- Matchett, W.E.; Anguiano-Zarate, S.S.; Nehete, P.N.; Shelton, K.; Nehete, B.P.; Yang, G.; Dorta-Estremera, S.; Barnette, P.; Xiao, P.; Byrareddy, S.N.; et al. Divergent HIV-1-Directed Immune Responses Generated by Systemic and Mucosal Immunization with Replicating Single-Cycle Adenoviruses in Rhesus Macaques. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [PubMed]
- Anguiano-Zarate, S.S.; Matchett, W.E.; Nehete, P.N.; Sastry, J.K.; Marzi, A.; Barry, M.A. A Replicating Single-Cycle Adenovirus Vaccine Against Ebola Virus. J. Infect. Dis. 2018, 218, 1883–1889. [Google Scholar] [CrossRef]
- Baliban, S.M.; Michael, A.; Shammassian, B.; Mudakha, S.; Khan, A.S.; Cocklin, S.; Zentner, I.; Latimer, B.P.; Bouillaut, L.; Hunter, M.; et al. An optimized, synthetic DNA vaccine encoding the toxin A and toxin B receptor binding domains of Clostridium difficile induces protective antibody responses in vivo. Infect. Immun. 2014, 82, 4080–4091. [Google Scholar] [CrossRef]
- Crosby, C.M.; Nehete, P.; Sastry, K.J.; Barry, M.A. Amplified and persistent immune responses generated by single-cycle replicating adenovirus vaccines. J. Virol. 2015, 89, 669–675. [Google Scholar] [CrossRef]
- Nyberg-Hoffman, C.; Shabram, P.; Li, W.; Giroux, D.; Aguilar-Cordova, E. Sensitivity and reproducibility in adenoviral infectious titer determination. Nat. Med. 1997, 3, 808–811. [Google Scholar] [CrossRef]
- Ji, H. Lysis of cultured cells for immunoprecipitation. Cold Spring Harb. Protoc. 2010, 2010, pdb.prot5466. [Google Scholar] [CrossRef]
- Frey, A.; Di Canzio, J.; Zurakowski, D. A statistically defined endpoint titer determination method for immunoassays. J. Immunol. Methods 1998, 221, 35–41. [Google Scholar] [CrossRef]
- Xie, J.; Zorman, J.; Indrawati, L.; Horton, M.; Soring, K.; Antonello, J.M.; Zhang, Y.; Secore, S.; Miezeiewski, M.; Wang, S.; et al. Development and optimization of a novel assay to measure neutralizing antibodies against Clostridium difficile toxins. Clin. Vaccine Immunol. 2013, 20, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, D.F.; Rosenberg, T.; Zaharatos, J.; Franco, D.; Ho, D.D. A DNA vaccine targeting the receptor-binding domain of Clostridium difficile toxin A. Vaccine 2009, 27, 3598–3604. [Google Scholar] [CrossRef]
- Seregin, S.S.; Aldhamen, Y.A.; Rastall, D.P.W.; Godbehere, S.; Amalfitano, A. Adenovirus-based vaccination against Clostridium difficile toxin A allows for rapid humoral immunity and complete protection from toxin A lethal challenge in mice. Vaccine 2012, 30, 1492–1501. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Qiu, H.; Cassan, R.; Johnstone, D.; Han, X.; Joyee, A.G.; McQuoid, M.; Masi, A.; Merluza, J.; Hrehorak, B.; Reid, R.; et al. Novel Clostridium difficile Anti-Toxin (TcdA and TcdB) Humanized Monoclonal Antibodies Demonstrate In Vitro Neutralization across a Broad Spectrum of Clinical Strains and In Vivo Potency in a Hamster Spore Challenge Model. PLoS ONE 2016, 11, e0157970. [Google Scholar] [CrossRef] [PubMed]
- Sorg, J.A.; Sonenshein, A.L. Inhibiting the initiation of Clostridium difficile spore germination using analogs of chenodeoxycholic acid, a bile acid. J. Bacteriol. 2010, 192, 4983–4990. [Google Scholar] [CrossRef]
- Anosova, N.G.; Brown, A.M.; Li, L.; Liu, N.; Cole, L.E.; Zhang, J.; Mehta, H.; Kleanthous, H. Systemic antibody responses induced by a two-component Clostridium difficile toxoid vaccine protect against C. difficile-associated disease in hamsters. J. Med. Microbiol. 2013, 62, 1394–1404. [Google Scholar] [CrossRef]
- Kuehne, S.A.; Cartman, S.T.; Heap, J.T.; Kelly, M.L.; Cockayne, A.; Minton, N.P. The role of toxin A and toxin B in Clostridium difficile infection. Nature 2010, 467, 711–713. [Google Scholar] [CrossRef]
- Suckow, M.; Stevens, K.; Wilson, R. The Laboratory Rabbit, Guinea Pig, Hamster, and Other Rodents; Academic Press: London, UK, 2012. [Google Scholar] [CrossRef]
- Hutton, M.L.; Mackin, K.E.; Chakravorty, A.; Lyras, D. Small animal models for the study of Clostridium difficile disease pathogenesis. FEMS Microbiol. Lett. 2014, 352, 140–149. [Google Scholar] [CrossRef]
- Rupnik, M.; Janezic, S. An Update on Clostridium difficile Toxinotyping. J. Clin. Microbiol. 2016, 54, 13–18. [Google Scholar] [CrossRef]
- Depestel, D.D.; Aronoff, D.M. Epidemiology of Clostridium difficile infection. J. Pharm. Pract. 2013, 26, 464–475. [Google Scholar] [CrossRef] [PubMed]
- Fischinger, S.; Boudreau, C.M.; Butler, A.L.; Streeck, H.; Alter, G. Sex differences in vaccine-induced humoral immunity. Semin. Immunopathol. 2019, 41, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Maeda, K.; West, K.; Hayasaka, D.; Ennis, F.A.; Terajima, M. Recombinant Adenovirus Vector Vaccine Induces Stronger Cytotoxic T-Cell Responses Than Recombinant Vaccinia Virus Vector, Plasmid DNA, or a Combination of These. Viral Immunol. 2005, 18, 657–667. [Google Scholar] [CrossRef] [PubMed]
- Quinn, K.M.; Da Costa, A.; Yamamoto, A.; Berry, D.; Lindsay, R.W.B.; Darrah, P.A.; Wang, L.; Cheng, C.; Kong, W.-P.; Gall, J.G.D.; et al. Comparative Analysis of the Magnitude, Quality, Phenotype, and Protective Capacity of Simian Immunodeficiency Virus Gag-Specific CD8+ T Cells following Human-, Simian-, and Chimpanzee-Derived Recombinant Adenoviral Vector Immunization. J. Immunol. 2013, 190, 2720–2735. [Google Scholar] [CrossRef]
- Best, E.L.; Freeman, J.; Wilcox, M.H. Models for the study of Clostridium difficile infection. Gut Microbes 2012, 3, 145–167. [Google Scholar] [CrossRef]
- Kuehne, S.A.; Collery, M.M.; Kelly, M.L.; Cartman, S.T.; Cockayne, A.; Minton, N.P. Importance of toxin A, toxin B, and CDT in virulence of an epidemic Clostridium difficile strain. J. Infect. Dis. 2014, 209, 83–86. [Google Scholar] [CrossRef]
- Wilcox, M.H.; Gerding, D.N.; Poxton, I.R.; Kelly, C.; Nathan, R.; Birch, T.; Cornely, O.A.; Rahav, G.; Bouza, E.; Lee, C.; et al. Bezlotoxumab for Prevention of Recurrent Clostridium difficile Infection. N. Engl. J. Med. 2017, 376, 305–317. [Google Scholar] [CrossRef]
- Mullard, A. FDA approves antitoxin antibody. Nat. Rev. Drug Discov. 2016, 15, 811. [Google Scholar] [CrossRef]
- Kyne, L.; Warny, M.; Qamar, A.; Kelly, C.P. Asymptomatic carriage of Clostridium difficile and serum levels of IgG antibody against toxin A. N. Engl. J. Med. 2000, 342, 390–397. [Google Scholar] [CrossRef]
- Océane, P. A review of 65 years of human adenovirus seroprevalence. Expert Rev. Vaccines 2019, 18. [Google Scholar] [CrossRef]
- Fausther-Bovendo, H.; Kobinger, G.P. Pre-existing immunity against Ad vectors: Humoral, cellular, and innate response, what’s important? Hum. Vaccines Immunother. 2014, 10, 2875–2884. [Google Scholar] [CrossRef] [PubMed]
- Vogels, R.; Zuijdgeest, D.; van Rijnsoever, R.; Hartkoorn, E.; Damen, I.; de Bethune, M.P.; Kostense, S.; Penders, G.; Helmus, N.; Koudstaal, W.; et al. Replication-deficient human adenovirus type 35 vectors for gene transfer and vaccination: Efficient human cell infection and bypass of preexisting adenovirus immunity. J. Virol. 2003, 77, 8263–8271. [Google Scholar] [CrossRef] [PubMed]
- D’Ambrosio, E.; Del Grosso, N.; Chicca, A.; Midulla, M. Neutralizing antibodies against 33 human adenoviruses in normal children in Rome. J. Hyg. 1982, 89, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Croyle, M.A.; Patel, A.; Tran, K.N.; Gray, M.; Zhang, Y.; Strong, J.E.; Feldmann, H.; Kobinger, G.P. Nasal delivery of an adenovirus-based vaccine bypasses pre-existing immunity to the vaccine carrier and improves the immune response in mice. PLoS ONE 2008, 3, e3548. [Google Scholar] [CrossRef]
- Richardson, J.S.; Pillet, S.; Bello, A.J.; Kobinger, G.P. Airway delivery of an adenovirus-based Ebola virus vaccine bypasses existing immunity to homologous adenovirus in nonhuman primates. J. Virol. 2013, 87, 3668–3677. [Google Scholar] [CrossRef]
- Munoz, R.E.; Piedra, P.A.; Demmler, G.J. Disseminated Adenovirus Disease in Immunocompromised and Immunocompetent Children. Clin. Infect. Dis. 1998, 27, 1194–1200. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matchett, W.E.; Anguiano-Zarate, S.; Malewana, G.B.R.; Mudrick, H.; Weldy, M.; Evert, C.; Khoruts, A.; Sadowsky, M.; Barry, M.A. A Replicating Single-Cycle Adenovirus Vaccine Effective against Clostridium difficile. Vaccines 2020, 8, 470. https://doi.org/10.3390/vaccines8030470
Matchett WE, Anguiano-Zarate S, Malewana GBR, Mudrick H, Weldy M, Evert C, Khoruts A, Sadowsky M, Barry MA. A Replicating Single-Cycle Adenovirus Vaccine Effective against Clostridium difficile. Vaccines. 2020; 8(3):470. https://doi.org/10.3390/vaccines8030470
Chicago/Turabian StyleMatchett, William E., Stephanie Anguiano-Zarate, Goda Baddage Rakitha Malewana, Haley Mudrick, Melissa Weldy, Clayton Evert, Alexander Khoruts, Michael Sadowsky, and Michael A. Barry. 2020. "A Replicating Single-Cycle Adenovirus Vaccine Effective against Clostridium difficile" Vaccines 8, no. 3: 470. https://doi.org/10.3390/vaccines8030470
APA StyleMatchett, W. E., Anguiano-Zarate, S., Malewana, G. B. R., Mudrick, H., Weldy, M., Evert, C., Khoruts, A., Sadowsky, M., & Barry, M. A. (2020). A Replicating Single-Cycle Adenovirus Vaccine Effective against Clostridium difficile. Vaccines, 8(3), 470. https://doi.org/10.3390/vaccines8030470