New Points of Departure for More Global Influenza Vaccine Use


{kind=link}
{kind=link}
Abstract
1. Introduction
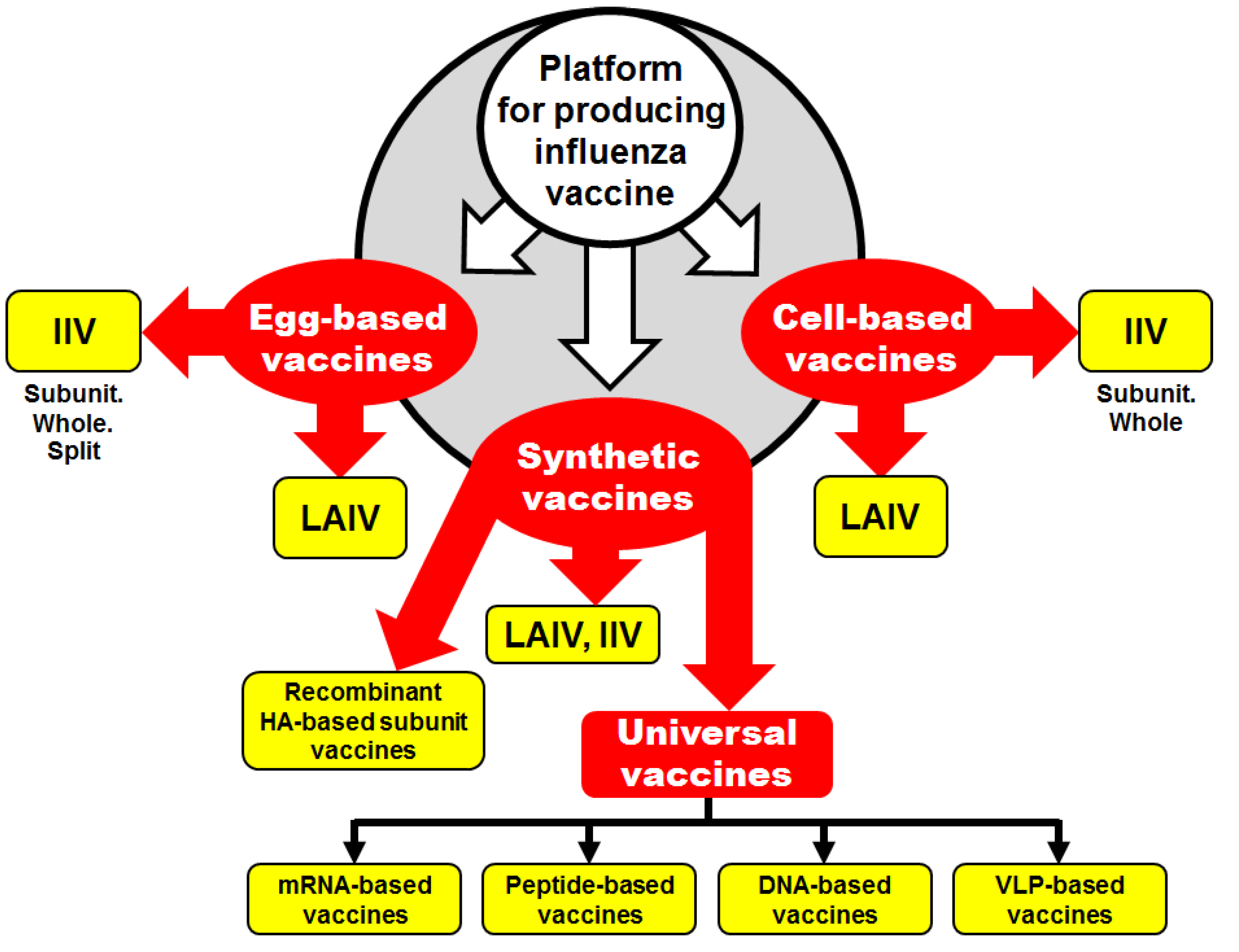
2. Influenza Vaccines: What is Next?
2.1. Egg-Derived Influenza Vaccines
2.2. Cell-Culture-Derived Influenza Vaccine
2.3. Synthetic Influenza Vaccines
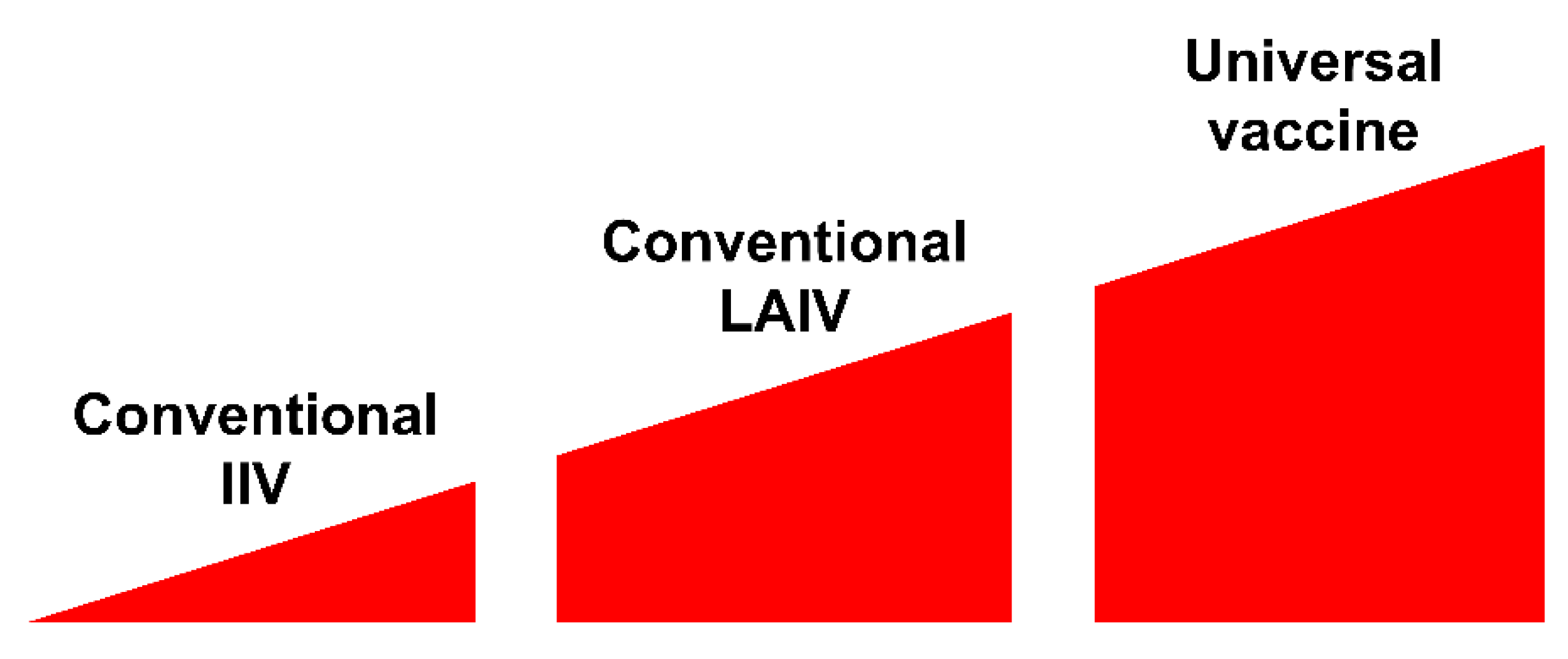
3. Cross-Protection Potency of Influenza Vaccines
4. The Possible Beneficial Effect of Bacillus Calmette-Guérin (BCG) and Influenza Vaccinations on the Severity of Illness after COVID-19 Infection
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Up to 650,000 People Die of Respiratory Diseases Linked to Seasonal Flu Each Year. Available online: https://www.who.int/news-room/detail/14-12-2017-up-to-650-000-people-die-of-respiratory-diseases-linked-to-seasonal-flu-each-year (accessed on 25 June 2020).
- Houser, K.; Subbarao, K. Influenza vaccines: Challenges and solutions. Cell Host Microbe 2015, 17, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Barberis, I.; Myles, P.; Ault, S.K.; Bragazzi, N.L.; Martini, M. History and evolution of influenza control through vaccination: From the first monovalent vaccine to universal vaccines. J. Prev. Med. Hyg. 2016, 57, E115–E120. [Google Scholar] [PubMed]
- Harding, A.T.; Heaton, N.S. Efforts to improve the seasonal influenza vaccine. Vaccines 2018, 6, 19. [Google Scholar] [CrossRef]
- Wareing, M.D.; Marsh, G.A.; Tannock, G.A. Preparation and characterisation of attenuated cold-adapted influenza A reassortants derived from the A/Leningrad/134/17/57 donor strain. Vaccine 2002, 20, 2082–2090. [Google Scholar] [CrossRef]
- Trucchi, C.; Paganino, C.; Amicizia, D.; Orsi, A.; Tisa, V.; Piazza, M.F.; Icardi, G.; Ansaldi, F. Universal influenza virus vaccines: What needs to happen next? Expert Opin. Biol. Ther. 2019, 19, 671–683. [Google Scholar] [CrossRef]
- WHO. Recommendations on the Composition of Influenza Virus Vaccines. Available online: https://www.who.int/influenza/vaccines/virus/recommendations/en/ (accessed on 25 June 2020).
- Dormitzer, P.R. Cell culture-derived influenza vaccines. In Influenza Vaccines for the Future. Birkhäuser Advances in Infectious Diseases; Rappuoli, R., Del Giudice, G., Eds.; Springer: Basel, Switzerland, 2011. [Google Scholar]
- Moro, P.L.; Winiecki, S.; Lewis, P.; Shimabukuro, T.T.; Cano, M. Surveillance of adverse events after the first trivalent inactivated influenza vaccine produced in mammalian cell culture (Flucelvax®) reported to the Vaccine Adverse Event Reporting System (VAERS), United States, 2013–2015. Vaccine 2015, 33, 6684–6688. [Google Scholar] [CrossRef]
- Wacheck, V.; Egorov, A.; Groiss, F.; Pfeiffer, A.; Fuereder, T.; Hoeflmayer, D.; Kundi, M.; Popow-Kraupp, T.; Redlberger-Fritz, M.; Mueller, C.A.; et al. A novel type of influenza vaccine: Safety and immunogenicity of replication-deficient influenza virus created by deletion of the interferon antagonist NS1. J. Infect. Dis. 2010, 201, 354–362. [Google Scholar] [CrossRef]
- Nachbagauer, R.; Palese, P. Is a universal influenza virus vaccine possible? Annu. Rev. Med. 2020, 71, 315–327. [Google Scholar] [CrossRef]
- Hoffmann, E.; Neumann, G.; Kawaoka, Y.; Hobom, G.; Webster, R.G. A DNA transfection system for generation of influenza A virus from eight plasmids. Proc. Natl. Acad. Sci. USA 2000, 97, 6108–6113. [Google Scholar] [CrossRef]
- Rajão, D.S.; Pérez, D.R. Universal vaccines and vaccine platforms to protect against influenza viruses in humans and agriculture. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Brazzoli, M.; Magini, D.; Bonci, A.; Buccato, S.; Giovani, C.; Kratzer, R.; Zurli, V.; Mangiavacchi, S.; Casini, D.; Brito, L.M.; et al. Induction of broad-based immunity and protective efficacy by self-amplifying mRNA vaccines encoding influenza virus hemagglutinin. J. Virol. 2016, 90, 332–344. [Google Scholar] [CrossRef]
- Bahl, K.; Senn, J.J.; Yuzhakov, O.; Bulychev, A.; Brito, L.A.; Hassett, K.J.; Laska, M.E.; Smith, M.; Almarsson, O.; Thompson, J.; et al. Preclinical and clinical demonstration of immunogenicity by mRNA vaccines against H10N8 and H7N9 influenza viruses. Mol. Ther. J. Am. Soc. Gene Ther. 2017, 25, 1316–1327. [Google Scholar] [CrossRef] [PubMed]
- Atsmon, J.; Caraco, Y.; Ziv-Sefer, S.; Shaikevich, D.; Abramov, E.; Volokhov, I.; Bruzil, S.; Haima, K.Y.; Gottlieb, T.; Ben-Yedidia, T. Priming by a novel universal influenza vaccine (Multimeric-001)—A gateway for improving immune response in the elderly population. Vaccine 2014, 32, 5816–5823. [Google Scholar] [CrossRef]
- Krammer, F.; Palese, P. Advances in the development of influenza virus vaccines. Nat. Rev. Drug. Discov. 2015, 14, 167–182. [Google Scholar] [CrossRef] [PubMed]
- Valero-Pacheco, N.; Perez-Toledo, M.; Villasis-Keever, M.A.; Nunez-Valencia, A.; Bosco-Garate, I.; Lozano-Dubernard, B.; Lara-Puente, H.; Espitia, C.; Alpuche-Aranda, C.; Bonifaz, L.C.; et al. Antibody persistence in adults two years after vaccination with an H1N1 2009 pandemic influenza virus-like particle vaccine. PLOS ONE 2016, 11, e0150146. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S.F.; McDermott, A.B. Shaping a universally broad antibody response to influenza amidst a variable immunoglobulin landscape. Curr. Opin. Immunol. 2018, 53, 96–101. [Google Scholar] [CrossRef]
- Elbahesh, H.; Saletti, G.; Gerlach, T.; Rimmelzwaan, G.F. Broadly protective influenza vaccines: Design and production platforms. Curr. Opin. Virol. 2019, 34, 1–9. [Google Scholar] [CrossRef]
- Fox, A.; Quinn, K.M.; Subbarao, K. Extending the breadth of influenza vaccines: Status and prospects for a universal vaccine. Drugs 2018, 78, 1297–1308. [Google Scholar] [CrossRef]
- Isakova-Sivak, I.; Matyushenko, V.; Kotomina, T.; Kiseleva, I.; Krutikova, E.; Donina, S.; Rekstin, A.; Larionova, N.; Mezhenskaya, D.; Sivak, K.; et al. Sequential immunization with universal live attenuated influenza vaccine candidates protects ferrets against a high-dose heterologous virus challenge. Vaccines 2019, 7, 61. [Google Scholar] [CrossRef]
- Jang, Y.H.; Seong, B.L. The quest for a truly universal influenza vaccine. Front. Cell. Infect. Microbiol. 2019, 9, 344. [Google Scholar] [CrossRef]
- Jiang, W.; Wang, S.; Chen, H.; Ren, H.; Huang, X.; Wang, G.; Chen, Z.; Chen, L.; Chen, Z.; Zhou, P. A bivalent heterologous DNA virus-like-particle prime-boost vaccine elicits broad protection against both group 1 and 2 influenza A viruses. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Meldgaard, T.S.; Bertholet, S. Novel platforms for the development of a universal influenza vaccine. Front. Immunol. 2018, 9, 600. [Google Scholar] [CrossRef] [PubMed]
- Madsen, A.; Cox, R.J. Prospects and challenges in the development of universal influenza vaccines. Vaccines 2020, 8, 361. [Google Scholar] [CrossRef] [PubMed]
- Nachbagauer, R.; Palese, P. Development of next generation hemagglutinin-based broadly protective influenza virus vaccines. Curr. Opin. Immunol. 2018, 53, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Atsmon, J.; Kate-Ilovitz, E.; Shaikevich, D.; Singer, Y.; Volokhov, I.; Haim, K.Y.; Ben-Yedidia, T. Safety and immunogenicity of multimeric-001—A novel universal influenza vaccine. J. Clin. Immunol. 2012, 32, 595–603. [Google Scholar] [CrossRef]
- WHO. Draft Landscape of COVID-19 Candidate Vaccines. Update of 9 July 2020. Available online: https://www.who.int/publications/m/item/draft-landscape-of-covid-19-candidate-vaccines (accessed on 10 July 2020).
- Burton, D.R.; Walker, L.M. Rational vaccine design in the time of COVID-19. Cell Host Microbe 2020, 27, 695–698. [Google Scholar] [CrossRef]
- Graham, B.S. Rapid COVID-19 vaccine development. Science 2020, 368, 945–946. [Google Scholar] [CrossRef]
- Eyal, N.; Lipsitch, M.; Smith, P.G. Human challenge studies to accelerate coronavirus vaccine licensure. J. Infect. Dis. 2020, 221, 1752–1756. [Google Scholar] [CrossRef]
- Coperchini, F.; Chiovato, L.; Croce, L.; Magri, F.; Rotondi, M. The cytokine storm in COVID-19: An overview of the involvement of the chemokine/chemokine-receptor system. Cytokine Growth Factor Rev. 2020, 53, 25–32. [Google Scholar] [CrossRef]
- Ozdemir, C.; Kucuksezer, U.C.; Tamay, Z.U. Is BCG vaccination affecting the spread and severity of COVID-19? Allergy 2020. [Google Scholar] [CrossRef]
- WHO. Bacille Calmette-Guérin (BCG) Vaccination and COVID-19. Scientific Brief of 12 April 2020. Available online: https://www.who.int/news-room/commentaries/detail/bacille-calmette-gu%C3%A9rin-(bcg)-vaccination-and-covid-19 (accessed on 25 June 2020).
- Dunning, J.; Thwaites, R.S.; Openshaw, P.J.M. Seasonal and pandemic influenza: 100 years of progress, still much to learn. Mucosal Immunol. 2020. [Google Scholar] [CrossRef]
- Salem, M.L.; El-Hennawy, D. The possible beneficial adjuvant effect of influenza vaccine to minimize the severity of COVID-19. Med Hypotheses 2020, 140, 109752. [Google Scholar] [CrossRef] [PubMed]
- Antony, S.J.; Almaghlouth, N.K.; Heydemann, E.L. Are co-infections with COVID-19 and Influenza low or underreported? An observational study examining current published literature including three new unpublished cases. J. Med. Virol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Ozaras, R.; Cirpin, R.; Duran, A.; Duman, H.; Arslan, O.; Bakcan, Y.; Kaya, M.; Mutlu, H.; Isayeva, L.; Kebanlı, F.; et al. Influenza and COVID-19 co-infection: Report of 6 cases and review of the Literature. J. Med. Virol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Osterhaus, A.D.M.E. Influenza and RSV in a COVID-19 World. The First ESWI Webinar 11 June 2020. Available online: https://eswi.org/eswi-tv/influenza-and-rsv-in-a-covid-19-world-full-webinar/ (accessed on 25 June 2020).
- Kiseleva, I. Will we ever be able to defeat human influenza? Open Microbiol. J. 2019, 13, 313–314. [Google Scholar] [CrossRef]
- Ozawa, M.; Kawaoka, Y. Cross talk between animal and human influenza viruses. Annu. Rev. Anim. Biosci. 2013, 1, 21–42. [Google Scholar] [CrossRef]


© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiseleva, I. New Points of Departure for More Global Influenza Vaccine Use. Vaccines 2020, 8, 410. https://doi.org/10.3390/vaccines8030410
Kiseleva I. New Points of Departure for More Global Influenza Vaccine Use. Vaccines. 2020; 8(3):410. https://doi.org/10.3390/vaccines8030410
Chicago/Turabian StyleKiseleva, Irina. 2020. "New Points of Departure for More Global Influenza Vaccine Use" Vaccines 8, no. 3: 410. https://doi.org/10.3390/vaccines8030410
APA StyleKiseleva, I. (2020). New Points of Departure for More Global Influenza Vaccine Use. Vaccines, 8(3), 410. https://doi.org/10.3390/vaccines8030410