A Replication-Defective Influenza Virus Vaccine Confers Complete Protection against H7N9 Viral Infection in Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Plasmids and the Generation of Reassortant Viruses
2.3. Western Blot Analysis
2.4. Plaque Assay and Viral Replication Curve
2.5. Ethics Statement
2.6. Mouse Trials and Sampling
2.7. Detection of IFN-γ and IL-5 Secreting Cells by Enzyme-Linked Immunospot (ELISPOT) Assay
2.8. Enzyme-Linked Immunosorbent Assay (ELISA) to Detect Antigen-Specific IgG, IgG1, and IgG2
2.9. Hemagglutination Inhibition (HAI) and the Serum Viral Neutralization (SVN) Assay
2.10. Virus Isolation and Titration
2.11. RNA Extraction and Quantitative RT-PCR (qRT-PCR)
2.12. Statistical Analysis
3. Results
3.1. The Generation of Two Reassortant H7N9 Viruses
3.2. The Reassortant Mutant QTV Virus Is Dependent on Elastase and Possesses Equivalent Replication Abilities as the WT Counterpart
3.3. The Reassortant Mutant QTV Virus Is Non-Virulent and Replication-Defective in Mice
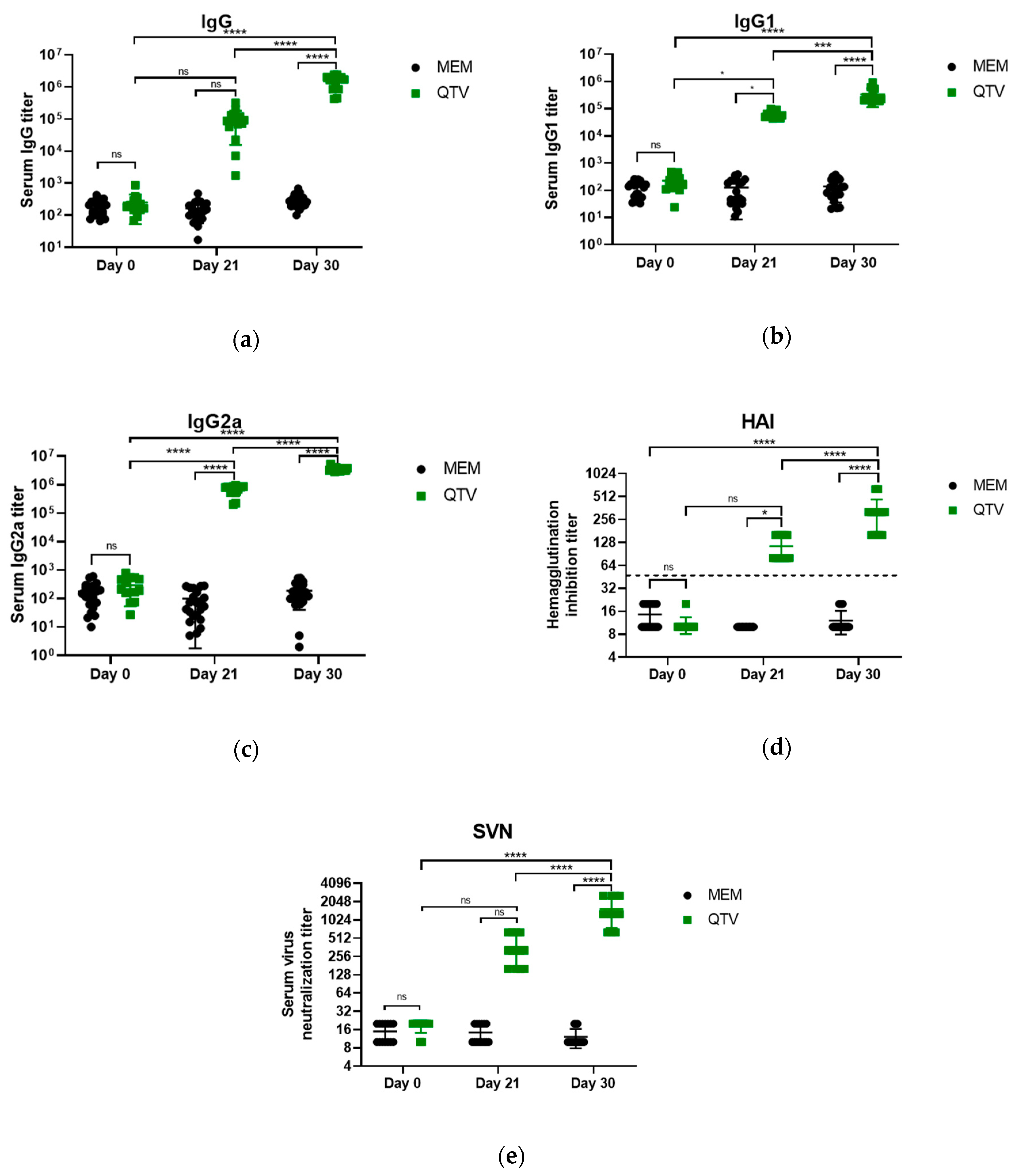
3.4. The Reassortant Mutant QTV Virus Induces Robust Antibody Responses
3.5. The Reassortant Mutant QTV Virus Produces High Numbers of Both IFN-γ and IL-5 Antigen-Specific Secreting Cells
3.6. Vaccination with the Reassortant Mutant QTV Virus Completely Protects Mice from a Lethal Challenge of BC15 (H7N9)
3.7. Vaccination with the Reassortant Mutant QTV Virus Reduces the Production of Proinflammatory Cytokines in the Mouse Lung Associated with the Influenza H7N9 Viral Infection
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. H7N9 Situation Update. Available online: http://www.fao.org/ag/againfo/programmes/en/empres/h7n9/situation_update.html (accessed on 15 March 2020).
- Gao, R.; Cao, B.; Hu, Y.; Feng, Z.; Wang, D.; Hu, W.; Chen, J.; Jie, Z.; Qiu, H.; Xu, K.; et al. Human Infection with a Novel Avian-Origin Influenza A (H7N9) Virus. N. Engl. J. Med. 2013, 368, 1888–1897. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; Gu, M.; Liu, D.; Cui, J.; Gao, G.F.; Zhou, J.; Liu, X. Epidemiology, Evolution, and Pathogenesis of H7N9 Influenza Viruses in Five Epidemic Waves since 2013 in China. Trends Microbiol. 2017, 25, 713–728. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wong, G.; Yang, L.; Tan, S.; Li, J.; Bai, B.; Xu, Z.; Li, H.; Xu, W.; Zhao, X.; et al. Comparison between human infections caused by highly and low pathogenic H7N9 avian influenza viruses in Wave Five: Clinical and virological findings. J. Infect. 2019, 78, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Skowronski, D.M.; Chambers, C.; Gustafson, R.; Purych, D.B.; Tang, P.; Bastien, N.; Krajden, M.; Li, Y. Avian Influenza A(H7N9) Virus Infection in 2 Travelers Returning from China to Canada. Emerg. Infect. Dis. 2016, 22, 71–74. [Google Scholar] [CrossRef]
- Zhang, F.; Bi, Y.; Wang, J.; Wong, G.; Shi, W.; Hu, F.; Yang, Y.; Yang, L.; Deng, X.; Jiang, S.; et al. Human infections with recently-emerging highly pathogenic H7N9 avian influenza virus in China. J. Infect. 2017, 75, 71–75. [Google Scholar] [CrossRef]
- Shi, J.; Jagger, B.W.; Wise, H.M.; Digard, P.; Holmes, E.C.; Taubenberger, J.K. Rapid Evolution of H7N9 Highly Pathogenic Viruses that Emerged in China in 2017. Cell Host Microbe 2018, 24, 558–568. [Google Scholar] [CrossRef]
- Bouvier, N.M.; Palese, P. The Biology of Influenza Viruses. Vaccine 2011, 26, 49–53. [Google Scholar] [CrossRef]
- Wong, S.S.; Webby, R.J. A single dose of whole inactivated H7N9 influenza vaccine confers protection from severe disease but not infection in ferrets. Vaccine 2014, 32, 4571–4577. [Google Scholar] [CrossRef]
- Couch, R.B. A Randomized Clinical Trial of an Inactivated Avian Influenza A (H7N7) Vaccine. PLoS ONE 2012, 7, e49704. [Google Scholar] [CrossRef]
- Yang, W.; Yin, X.; Guan, L.; Li, M.; Ma, S.; Shi, J.; Deng, G.; Suzuki, Y.; Chen, H. A live attenuated vaccine prevents replication and transmission of H7N9 highly pathogenic influenza viruses in mammals. Emerg. Microbes Infect. 2019, 7, 1–10. [Google Scholar] [CrossRef]
- Lorenzo, M.M.G.; Fenton, M.J. Immunobiology of Influenza Vaccines. Chest 2013, 143, 502–510. [Google Scholar] [CrossRef]
- Sridhar, S.; Brokstad, K.A.; Cox, R.J. Influenza Vaccination Strategies: Comparing Inactivated and Live Attenuated Influenza Vaccines. Vaccine 2015, 3, 373–389. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Wantanabe, S.; Neumann, G.; Kida, H.; Kawaoka, Y. Immunogenicity and Protective Efficacy of ReplicationIncompetent Influenza Virus-Like Particles. J. Virol. 2001, 76, 767–773. [Google Scholar] [CrossRef]
- Dudek, T.; Knipe, D.M. Replication-defective viruses as vaccines and vaccine vectors. Virology 2006, 344, 230–239. [Google Scholar] [CrossRef]
- Masic, A.; Babiuk, L.A.; Zhou, Y. Reverse genetics-generated elastase-dependent swine influenza viruses are attenuated in pigs. J. Gen. Virol. 2008, 90, 375–385. [Google Scholar] [CrossRef]
- Böttcher, E.; Matrosovich, T.Y.; Beyerle, M.; Klenk, H.D.; Garten, W.; Matrosovich, M. Proteolytic Activation of Influenza Viruses by Serine Proteases TMPRSS2 and HAT from Human Airway Epithelium. J. Virol. 2006, 80, 9896–9898. [Google Scholar] [CrossRef]
- Skehel, J.J.; Wiley, C. Receptor binding and membrane fusion in virus entry: The influenza hemagglutinin. Annu. Rev. Biochem. 2000, 69, 531–569. [Google Scholar] [CrossRef]
- Hoffmann, E.; Neumann, G.; Kawaoka, Y.; Hobom, G.; Webster, R.G. A DNA transfection system for generation of influenza A virus from eight plasmids. Proc. Natl. Acad. Sci. USA 2000, 97, 6108–6113. [Google Scholar] [CrossRef]
- Lu, Y.; Landreth, S.; Liu, G.; Brownlie, R.; Gaba, A.; Littel-van den Hurk, S.V.D.; Gerdts, V.; Zhou, Y. Innate immunemodulator containing adjuvant formulated HA based vaccine protects mice from lethal infection of highly pathogenic avian influenza H5N1 virus. Vaccines 2020, 38, 2387–2395. [Google Scholar] [CrossRef]
- Kärber, G. Beitrag zur kollektiven Behandlung pharmakologischer Reihenversuche. Naunyn-Schmiedebergs Arch. Exp. Pathol. Pharmakol. 1931, 162, 480–483. [Google Scholar] [CrossRef]
- Spearman, C. The Method of ‘Right and Wrong Cases’ (‘Constant Stimuli’) without Gauss’s Formulae. Br. J. Psychol. 1904–1920 1908, 2, 227–242. [Google Scholar] [CrossRef]
- Rao, X.; Huang, X.; Zhou, Z.; Lin, X. An improvement of the 2ˆ(–delta delta CT) method for quantitative real-time polymerase chain reaction data analysis. Biostat. Bioinform. Biomath. 2013, 3, 71–85. [Google Scholar]
- Lu, Y.; Landreth, S.; Gaba, A.; Hlasny, M.; Liu, G.; Huang, Y.; Zhou, Y. In Vivo Characterization of Avian Influenza A (H5N1) and (H7N9) Viruses Isolated from Canadian Travelers. Viruses 2019, 11, 193. [Google Scholar] [CrossRef]
- Trombetta, C.M.; Remarque, E.J.; Mortier, D.; Montomoli, E. Comparison of hemagglutination inhibition, single radial hemolysis, virus neutralization assays, and ELISA to detect antibody levels against seasonal influenza viruses. Influenza Other Respir. Viruses 2018, 12, 675–686. [Google Scholar] [CrossRef]
- Si, L.; Xu, H.; Zhou, X.; Zhang, Z.; Tian, Z.; Yang, Y.; Wu, Y.; Zhang, B.; Niu, Z.; Zhang, C.; et al. Generation of influenza A viruses as live but replication-incompetent virus vaccines. Science 2016, 354, 1170–1173. [Google Scholar] [CrossRef]
- Wacheck, V.; Egorov, A.; Groiss, F.; Pfeiffer, A.; Fuereder, T.; Hoeflmayer, D.; Kundi, M.; Popow-Kraupp, T.; Redlberger-Fritz, M.; Mueller, C.A.; et al. A novel type of influenza vaccine: Safety and immunogenicity of replication-deficient influenza virus created by deletion of the interferon antagonist NS1. J. Infect. Dis. 2010, 201, 354–362. [Google Scholar] [CrossRef]
- Victor, S.T.; Watanabe, S.; Katsura, H.; Ozawa, M.; Kawaoka, Y. A Replication-Incompetent PB2-Knockout Influenza A Virus Vaccine Vector. J. Virol. 2012, 86, 4123–4128. [Google Scholar] [CrossRef]
- Thomas, P.G.; Keating, R.; Hulse-Post, D.J.; Doherty, P.C. Cell-mediated Protection in Influenza Infection. Emerg. Infect. Dis. 2006, 12, 48–54. [Google Scholar] [CrossRef]
- Bahadoran, A.; Lee, S.H.; Wang, S.M.; Manikam, R.; Rajarajeswaran, J.; Raju, C.S.; Sekaran, S.D. Immune Responses to Influenza Virus and Its Correlation to Age and Inherited Factors. Front. Microbiol. 2016, 7, 1841. [Google Scholar] [CrossRef]
- Chen, X.; Liu, S.; Goraya, M.U.; Maarouf, M.; Huang, S.; Chen, J.L. Host Immune Response to Influenza A Virus Infection. Front. Immunol. 2018, 9, 320. [Google Scholar] [CrossRef]
- Tau, G.; Rothman, P. Biologic functions of the IFN-γ receptors. Allergy 1999, 54, 1233–1251. [Google Scholar] [CrossRef]
- Mountford, A.P.; Fisher, A.; Wilson, R.A. The profile of IgG1 and IgG2a antibody responses in mice exposed to Schistosoma mansoni. Parasite Immunol. 1994, 16, 521–527. [Google Scholar] [CrossRef]
- D’Elia, R.V.; Harrison, K.; Oyston, P.C.; Lukaszewski, R.A.; Clark, G.C. Targeting the “Cytokine Storm” for Therapeutic Benefit. Clin. Vaccine Immunol. 2013, 20, 319–327. [Google Scholar] [CrossRef]
- Tamada, T.; Kinoshita, T.; Kurihara, K.; Adachi, M.; Ohhara, T.; Imai, K.; Kuroki, R.; Tada, T. Combined High-Resolution Neutron and X-ray Analysis of Inhibited Elastase Confirms the Active-Site Oxyanion Hole but Rules against a Low-Barrier Hydrogen Bond. J. Am. Chem. Soc. 2009, 131, 11033–11040. [Google Scholar] [CrossRef]
- Appel, W. Elastase: General Information. Methods Enzym. Anal. 1974, 2, 1041–1045. [Google Scholar]
- Kuhn, C.; Senior, R.M. The role of elastases in the development of emphysema. Lung 1978, 155, 185–187. [Google Scholar] [CrossRef]
- Gramegna, A.; Amati, F.; Terranova, L.; Sotgiu, G.; Tarsia, P.; Miglietta, D.; Calderazzo, M.A.; Aliberti, S.; Blasi, F. Neutrophil elastase in bronchiectasis. Respir. Res. 2017, 18, 211. [Google Scholar] [CrossRef]
- Stech, J.; Garn, H.; Wegmann, M.; Wagner, R.; Klenk, H.D. A new approach to an influenza live vaccine: Modification of the cleavage site of hemagglutinin. Nat. Med. 2005, 11, 683–689. [Google Scholar] [CrossRef]
- Babiuk, S.; Masic, A.; Graham, J.; Neufeld, J.; Van Der Loop, M.; Copps, J.; Berhane, Y.; Pasick, J.; Potter, A.; Babiuk, L.A.; et al. An elastase-dependent attenuated heterologous swine influenza virus protects against pandemic H1N1 2009 influenza challenge in swine. Vaccine 2011, 29, 3118–3123. [Google Scholar] [CrossRef]
- Gabriel, G.; Garn, H.; Wegmann, M.; Renz, H.; Herwig, A.; Klenk, H.D.; Stech, J. The potential of a protease activation mutant of a highly pathogenic avian influenza virus for a pandemic live vaccine. Vaccine 2008, 26, 956–965. [Google Scholar] [CrossRef]
- Mamerow, S.; Scheffter, R.; Röhrs, S.; Stech, O.; Blohm, U.; Schwaiger, T.; Schröder, C.; Ulrich, R.; Schinköthe, J.; Beer, M.; et al. Double-attenuated influenza virus elicits broad protection against challenge viruses with different serotypes in swine. Vet. Microbiol. 2019, 231, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Masic, A.; Booth, J.S.; Mutwiri, G.K.; Babiuk, L.A.; Zhou, Y. Elastase-dependent Live Attenuated Swine Influenza A Viruses Are Immunogenic and Confer Protection against Swine Influenza A Virus Infection in Pigs. J. Virol. 2009, 83, 10198–10210. [Google Scholar] [CrossRef] [PubMed]
- Masic, A.; Lu, X.; Li, J.; Mutwiri, G.K.; Babiuk, L.A. Immunogenicity and protective efficacy of an elastase-dependent live attenuated swine influenza virus vaccine administered intranasally in pigs. Vaccine 2010, 28, 7098–7108. [Google Scholar] [CrossRef] [PubMed]
- Stech, J.; Garn, H.; Herwig, A.; Stech, O.; Dauber, B.; Wolff, T.; Mettenleiter, T.C.; Klenk, H.D. Influenza B Virus with Modified Hemagglutinin Cleavage Site as a Novel Attenuated Live Vaccine. J. Infect. Dis. 2011, 204, 1483–1490. [Google Scholar] [CrossRef] [PubMed]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Landreth, S.; Lu, Y.; Pandey, K.; Zhou, Y. A Replication-Defective Influenza Virus Vaccine Confers Complete Protection against H7N9 Viral Infection in Mice. Vaccines 2020, 8, 207. https://doi.org/10.3390/vaccines8020207
Landreth S, Lu Y, Pandey K, Zhou Y. A Replication-Defective Influenza Virus Vaccine Confers Complete Protection against H7N9 Viral Infection in Mice. Vaccines. 2020; 8(2):207. https://doi.org/10.3390/vaccines8020207
Chicago/Turabian StyleLandreth, Shelby, Yao Lu, Kannupriya Pandey, and Yan Zhou. 2020. "A Replication-Defective Influenza Virus Vaccine Confers Complete Protection against H7N9 Viral Infection in Mice" Vaccines 8, no. 2: 207. https://doi.org/10.3390/vaccines8020207
APA StyleLandreth, S., Lu, Y., Pandey, K., & Zhou, Y. (2020). A Replication-Defective Influenza Virus Vaccine Confers Complete Protection against H7N9 Viral Infection in Mice. Vaccines, 8(2), 207. https://doi.org/10.3390/vaccines8020207