Humanized Mice in Dengue Research: A Comparison with Other Mouse Models

 , , , and
, , , and
Abstract
1. Introduction
1.1. Virus
1.2. Epidemiology
1.3. Immune Response and Pathogenesis of Dengue Virus
2. Mouse Models for Dengue Research
2.1. Wild Type Mouse Models
2.2. Knockout Mouse Models for the Study of DENV
2.2.1. AG129
2.2.2. STAT
2.2.3. C57BL/6 DKO, TKO, Irf1−/−, Ifnar1−/−, TNF−/−, and CCR5−/−
2.3. Transgenic Non-Humanized Model of DENV
C57BL/6J hTNF+++, IFNα/βR−/− Tg, Tg HLA-A*02:01, and B10.Tg HLA-DR3
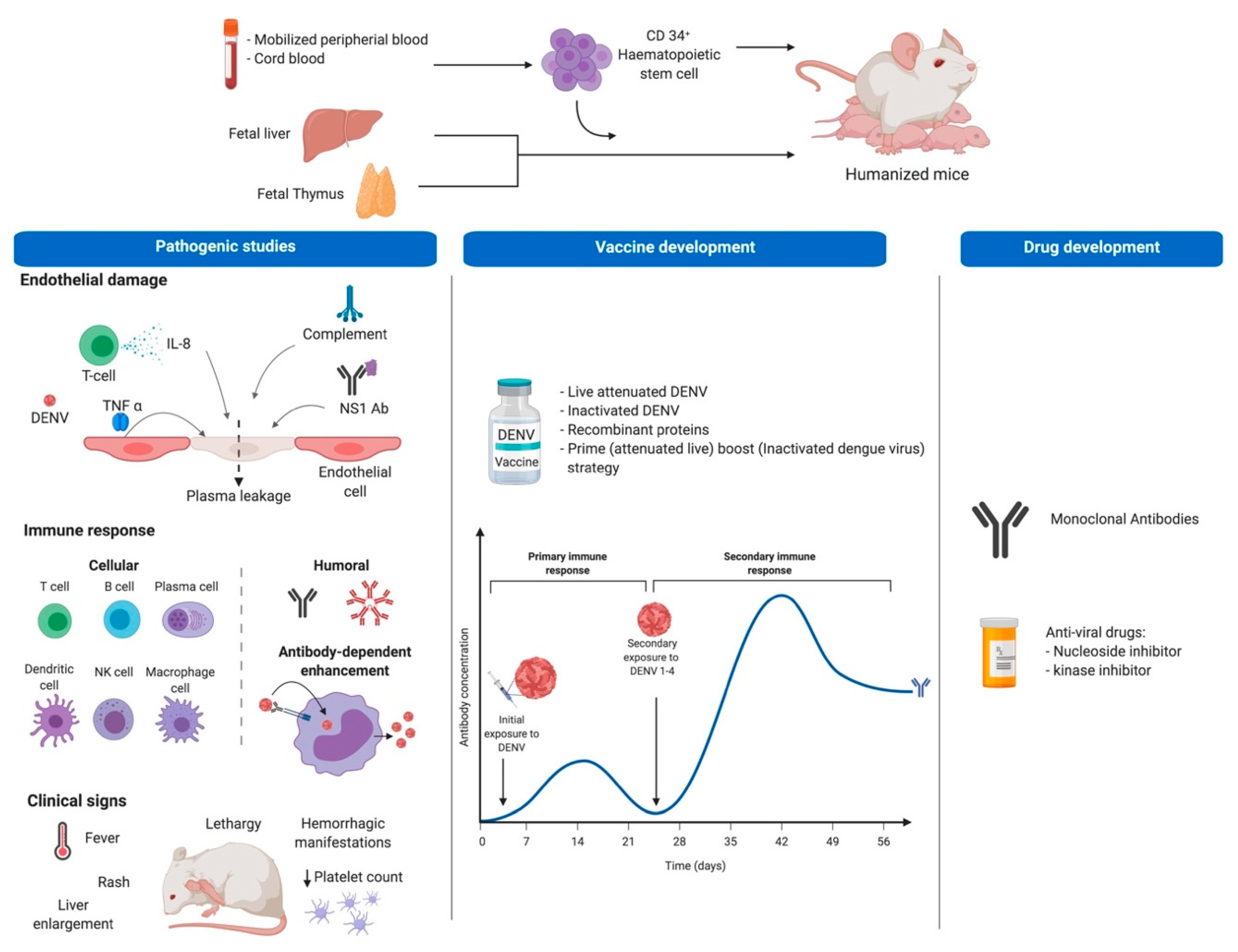
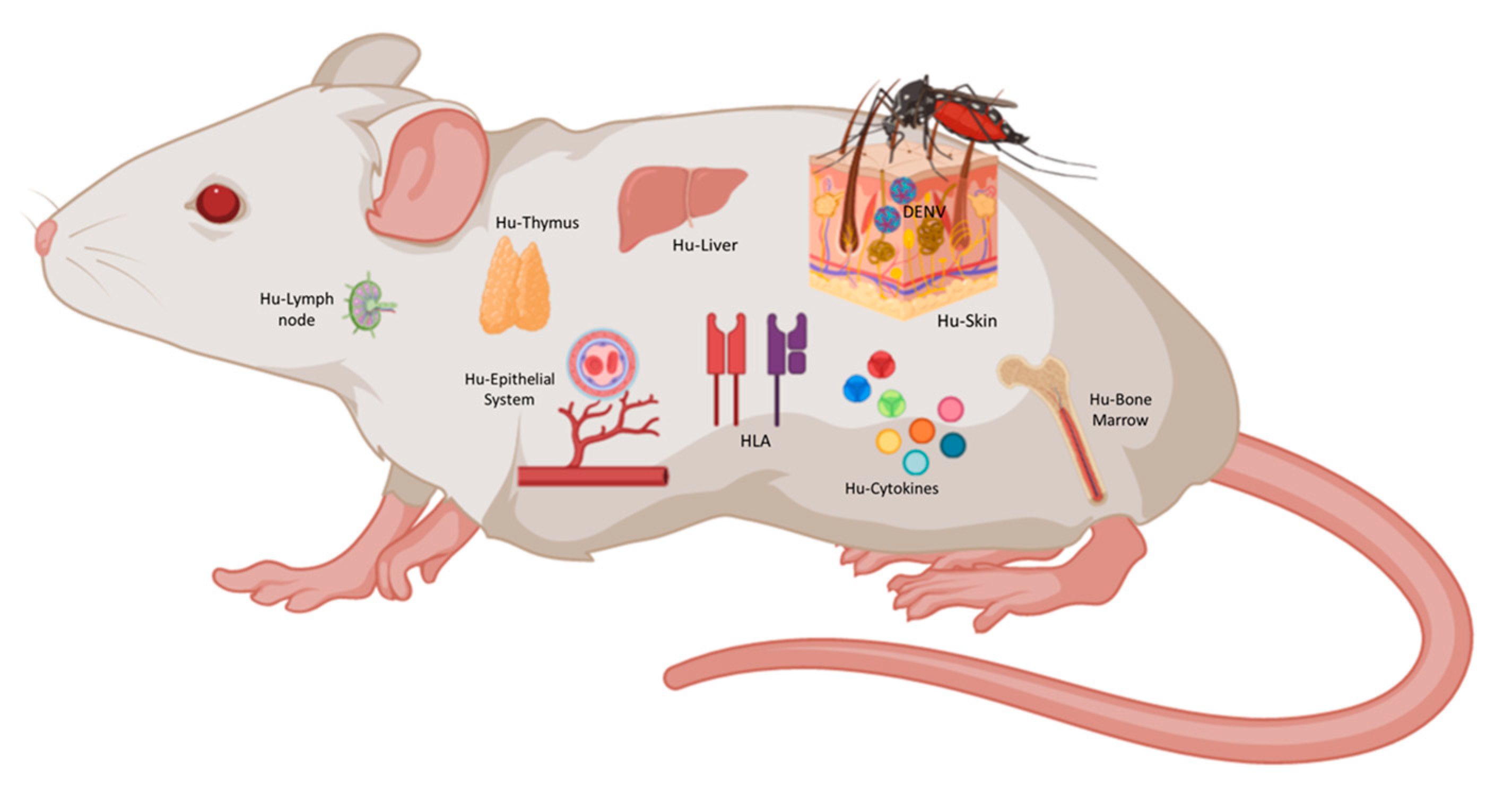
2.4. Humanized Mouse Models of DENV
2.4.1. SCID
2.4.2. NOD/SCID, and NSG
2.4.3. NSG-BLT and NOD/SCID-BLT
2.4.4. NSG HLA Class I Transgenic Strains, Expressing A2 Haplotypes
2.4.5. NSG-SGM3 and NOG-EXL Mouse Model
2.4.6. NRG
2.4.7. RAG2 and DRAG
3. Mouse Models for the Development of Dengue Vaccines
3.1. Mouse Models to Test Safety and Immunogenicity of DENV-Vaccine Candidates
3.2. Humanized Mice Models for the Development of Dengue Vaccines
3.2.1. Hu-NSG
3.2.2. BLT-NSG
3.2.3. Hu-DRAG
3.2.4. Ideal Humanized Mouse Model for DENV Pathogenesis Studies and Vaccine Development
4. Treatment
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lescar, J.; Soh, S.; Lee, L.T.; Vasudevan, S.G.; Kang, C.; Lim, S.P. The Dengue Virus Replication Complex: From RNA Replication to Protein-Protein Interactions to Evasion of Innate Immunity. In Advances in Experimental Medicine and Biology; Springer Nature: Singapore, 2018; pp. 115–129. ISBN 9789811087271. [Google Scholar]
- Rodenhuis-Zybert, I.A.; Wilschut, J.; Smit, J.M. Dengue virus life cycle: Viral and host factors modulating infectivity. Cell. Mol. Life Sci. 2010, 67, 2773–2786. [Google Scholar] [CrossRef]
- Guzman, M.G.; Gubler, D.J.; Izquierdo, A.; Martinez, E.; Halstead, S.B. Dengue infection. Nat. Rev. 2016, 2, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Sanchez, E.; Altmeyer, R.; Amara, A.; Schwartz, O.; Fieschi, F.; Virelizier, J.L.; Arenzana-Seisdedos, F.; Desprès, P. Dendritic-Cell-Specific ICAM3-Grabbing non-Integrin is essential for the productive infection of human dendritic cells by mosquito-Cell-Derived dengue viruses. EMBO Rep. 2003, 4, 723–728. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, S.Y.; King, C. Bacterial lipopolysaccharide inhibits dengue virus infection of primary human monocytes/macrophages by blockade of virus entry via a CD14-Dependent mechanism. J. Virol. 1999, 73, 2650–2657. [Google Scholar] [CrossRef]
- Reyes-del Valle, J.; Chavez-Salinas, S.; Medina, F.; del Angel, R.M. Heat Shock Protein 90 and Heat Shock Protein 70 Are Components of Dengue Virus Receptor Complex in Human Cells. J. Virol. 2005, 79, 4557–4567. [Google Scholar] [CrossRef] [PubMed]
- Jindadamrongwech, S.; Thepparit, C.; Smith, D.R. Identification of GRP 78 (BiP) as a liver cell expressed receptor element for dengue virus serotype 2. Arch. Virol. 2004, 149, 915–927. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Oliveira, C.; Freire, M.; Conceicao, T.M.; Higa, L.M.; Castancho, M.A.R.; Da Poian, A.T. Receptors and routes of dengue virus entry into the host cells. FEMS Microbiol. Rev. 2015, 39, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, M.N.; Sukumaran, B.; Pal, U.; Agaisse, H.; Murray, J.L.; Hodge, T.W.; Fikrig, E. Rab 5 Is Required for the Cellular Entry of Dengue and West Nile Viruses. J. Virol. 2007, 81, 4881–4885. [Google Scholar] [CrossRef]
- Acosta, E.G.; Castilla, V.; Damonte, E.B. Functional entry of dengue virus into Aedes albopictus mosquito cells is dependent on clathrin-Mediated endocytosis. J. Gen. Virol. 2008, 89, 474–484. [Google Scholar] [CrossRef]
- Freire, J.M.; Veiga, A.S.; de la Torre, B.G.; Santos, N.C.; Andreu, D.; Da Poian, A.T.; Castanho, M.A. Peptides as models for the structure and function of viral capsid proteins: Insights on dengue virus capsid. Biopolymers 2013, 100, 325–336. [Google Scholar] [CrossRef]
- Arakawa, M.; Morita, E. Flavivirus replication organelle biogenesis in the endoplasmic reticulum: Comparison with other single-Stranded positive-sense RNA viruses. Int. J. Mol. Sci. 2019, 20, 2336. [Google Scholar] [CrossRef] [PubMed]
- Coloma, J.; Harris, E. Broad and strong: The ultimate antibody to dengue virus. Nat. Immunol. 2015, 16, 135–137. [Google Scholar] [CrossRef]
- Sangkawibha, N.; Rojanasuphot, S.; Ahandrik, S.; Viriyapongse, S.; Jatanasen, S.; Salitul, V.; Phanthumachinda, B.; Halstead, S.B. Risk factors in dengue shock syndrome: A prospective epidemiologic study in rayong, thailand. Am. J. Epidemiol. 1984, 120, 653–669. [Google Scholar] [CrossRef] [PubMed]
- Guzman, M.G.; Alvarez, M.; Halstead, S.B. Secondary infection as a risk factor for dengue hemorrhagic fever/dengue shock syndrome: An historical perspective and role of antibody-Dependent enhancement of infection. Arch. Virol. 2013, 158, 1445–1459. [Google Scholar] [CrossRef] [PubMed]
- Guzman, M.G.; Kouri, G.; Bravo, J.; Valdes, L.; Vasquez, S.; Halstead, S.B. Effect of age on outcome dengue 2 infections of secondary. Int. J. Infect. Dis. 2002, 6, 118–124. [Google Scholar] [CrossRef]
- Capeding, M.R.; Tran, N.H.; Hadinegoro, S.R.S.; Muhammad Ismail, H.I.; Chotpitayasunondh, T.; Chua, M.N.; Luong, C.Q.; Rusmil, K.; Wirawan, D.N.; Nallusamy, R. Clinical effi cacy and safety of a novel tetravalent dengue vaccine in healthy children in Asia: A phase 3, randomised, observer-Masked, placebo-Controlled trial. Lancet 2014, 384, 1358–1365. [Google Scholar] [CrossRef]
- Villar, L.; Dayan, G.H.; Arredondo-García, J.L.; Rivera, D.M.; Cunha, R.; Deseda, C.; Reynales, H.; Costa, M.S.; Morales-Ramírez, J.O.; Carrasquilla, G.; et al. Efficacy of a Tetravalent Dengue Vaccine in Children in Latin America. N. Engl. J. Med. 2014, 372, 141103114505002. [Google Scholar] [CrossRef]
- Sridhar, S.; Luedtke, A.; Langevin, E.; Zhu, M.; Bonaparte, M.; Savarino, S.; Zambrano, B.; Moureau, A.; Khromava, A.; Moodie, Z.; et al. Effect of Dengue Serostatus on Dengue Vaccine Safety and Efficacy. N. Engl. J. Med. 2018, 379, 327–330. [Google Scholar] [CrossRef]
- Roth, C.; Delgado, F.G.; Simon-Lorière, E.; Sakuntabhai, A. Immune responses to dengue and Zika viruses-Guidance for T cell vaccine development. Int. J. Environ. Res. Public Health 2018, 15, 385. [Google Scholar] [CrossRef]
- Halstead, S.B.; Dans, L.F. Dengue infection and advances in dengue vaccines for children. Lancet Child Adolesc. Health 2019, 3, 734–741. [Google Scholar] [CrossRef]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Dengue and Severe Dengue. 2019. Available online: https://www.who.int/en/news-room/fact-sheets/detail/dengue-and-severe-dengue (accessed on 27 October 2019).
- Brady, O.J.; Gething, P.W.; Bhatt, S.; Messina, J.P.; Brownstein, J.S.; Hoen, A.G.; Moyes, C.L.; Farlow, A.W.; Scott, T.W.; Hay, S.I. Refining the Global Spatial Limits of Dengue Virus Transmission by Evidence-Based Consensus. PLoS Med. 2012, 6, e1760. [Google Scholar] [CrossRef] [PubMed]
- Kelley, J.F.; Kaufusi, P.H.; Nerurkar, V.R. Dengue Hemorrhagic Fever-Associated Immunomediators Induced via Maturation of Dengue Virus Nonstructural 4B Protein in Monocytes Modulate Endothelial Cell Adhesion Molecules and Human Microvascular Endothelial Cells Permeability. Virology 2012, 422, 326–337. [Google Scholar] [CrossRef] [PubMed]
- John, A.L.S.; Rathore, A.P.S.; Raghavan, B.; Ng, M.; Abraham, S.N. Contributions of mast cells and vasoactive products, leukotrienes and chymase, to dengue virus-Induced vascular leakage. Elife 2013, 2, e00481. [Google Scholar] [CrossRef]
- Puerta-Guardo, H.; Raya-Sandino, A.; Gonzalez-Mariscal, L.; Rosales, V.H.; Ayala-Davila, J.; Chavez-Mungia, B.; Martinez-Fong, D.; Medina, F.; Ludert, J.E.; del Angel, R.M. The Cytokine Response of U937-Derived Macrophages Infected through Antibody-Dependent Enhancement of Dengue Virus Disrupts Cell Apical-Junction Complexes and Increases Vascular Permeability. J. Virol. 2013, 87, 7486–7501. [Google Scholar] [CrossRef][Green Version]
- Martina, B.E.E.; Koraka, P.; Osterhaus, A.D.M.E. Dengue Virus Pathogenesis: An Integrated View. Clin. Microbiol. Rev. 2009, 22, 564–581. [Google Scholar] [CrossRef]
- Vaughn, D.W.; Green, S.; Kalayanarooj, S.; Innis, B.L.; Nimmannitya, S.; Suntayakorn, S.; Endy, T.P.; Raengsakulrach, B.; Rothman, A.L.; Ennis, F.A. Dengue Viremia Titer, Antibody Response Pattern, and Virus Serotype Correlate with Disease Severity. J. Infect. Dis. 2000, 181, 2–9. [Google Scholar] [CrossRef]
- Libraty, D.H.; Young, P.R.; Pickering, D.; Endy, T.P.; Kalayanarooj, S.; Green, S.; Vaughn, D.W.; Nisalak, A.; Ennis, F.A.; Rothman, A.L. High Circulating Levels of the Dengue Virus Nonstructural Protein NS1 Early in Dengue Illness Correlate with the Development of Dengue Hemorrhagic Fever. J. Infect. Dis. 2002, 186, 2000. [Google Scholar] [CrossRef]
- Garcia, M.; Wehbe, M.; Lévêque, N.; Bodet, C. Skin innate immune response to flaviviral infection. Eur. Cytokine Netw. 2017, 28, 41–51. [Google Scholar] [CrossRef]
- Nasirudeen, A.M.A.; Wong, H.H.; Thien, P.; Xu, S.; Lam, K.P.; Liu, D.X. RIG-I, MDA5 and TLR3 synergistically play an important role in restriction of dengue virus infection. PLoS Negl. Trop. Dis. 2011, 5, e926. [Google Scholar] [CrossRef]
- Sariol, C.A.; Martínez, M.I.; Rivera, F.; Rodríguez, I.V.; Pantoja, P.; Abel, K.; Arana, T.; Giavedoni, L.; Hodara, V.; White, L.J.; et al. Decreased dengue replication and an increased anti-Viral humoral response with the use of combined toll-Like receptor 3 and 7/8 agonists in macaques. PLoS ONE 2011, 6, e19323. [Google Scholar] [CrossRef] [PubMed]
- Schoggins, J.W.; MacDuff, D.A.; Imanaka, N.; Gainey, M.D.; Shrestha, B.; Eitson, J.L.; Mar, K.B.; Richardson, R.B.; Ratushny, A.V.; Litvak, V.; et al. Pan-Viral specificity of IFN-Induced genes reveals new roles for cGAS in innate immunity. Nature 2014, 505, 691–695. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.; Wang, M.; Huang, C.; Wu, C.; Hung, L.; Yang, C.; Ke, P.; Luo, S.; Liu, S.; Ho, L. Infection with the dengue RNA virus activates TLR9 signaling in human dendritic cells. EMBO Rep. 2018, 19, e46182. [Google Scholar] [CrossRef] [PubMed]
- Cabezas, S.; Bracho, G.; Aloia, A.L.; Adamson, P.J.; Bonder, C.S.; Smith, J.R.; Gordon, D.L.; Carr, J.M. Dengue Virus Induces Increased Activity of the Complement Alternative Pathway in Infected Cells. J. Virol. 2018, 92, e00633-18. [Google Scholar] [CrossRef]
- Surasombatpattana, P.; Hamel, R.; Patramool, S.; Luplertlop, N.; Thomas, F.; Desprès, P.; Briant, L.; Yssel, H.; Missé, D. Dengue virus replication in infected human keratinocytes leads to activation of antiviral innate immune responses. Infect. Genet. Evol. 2011, 11, 1664–1673. [Google Scholar] [CrossRef]
- Bustos-Arriaga, J.; García-Machorro, J.; León-Juárez, M.; García-Cordero, J.; Santos-Argumedo, L.; Flores-Romo, L.; Méndez-Cruz, A.R.; Juárez-Delgado, F.J.; Cedillo-Barrón, L. Activation of the innate immune response against denv in normal non-Transformed human fibroblasts. PLoS Negl. Trop. Dis. 2011, 5, e1420. [Google Scholar] [CrossRef]
- Troupin, A.; Shirley, D.; Londono-Renteria, B.; Watson, A.M.; McHale, C.; Hall, A.; Hartstone-Rose, A.; Klimstra, W.B.; Gomez, G.; Colpitts, T.M. A Role for Human Skin Mast Cells in Dengue Virus Infection and Systemic Spread. J. Immunol. 2016, 197, 4382–4391. [Google Scholar] [CrossRef]
- Libraty, D.H.; Pichyangkul, S.; Ajariyakhajorn, C.; Endy, T.P.; Ennis, F.A. Human Dendritic Cells Are Activated by Dengue Virus Infection: Enhancement by Gamma Interferon and Implications for Disease Pathogenesis. J. Virol. 2001, 75, 3501–3508. [Google Scholar] [CrossRef] [PubMed]
- Lertjuthaporn, S.; Khowawisetsut, L.; Keawvichit, R.; Polsrila, K.; Chuansumrit, A.; Chokephaibulkit, K.; Thitilertdecha, P.; Onlamoon, N.; Ansari, A.A.; Pattanapanyasat, K. Identification of changes in dendritic cell subsets that correlate with disease severity in dengue infection. PLoS ONE 2018, 13, e0200564. [Google Scholar] [CrossRef]
- Martins, S.d.T.; Silveira, G.F.; Alves, L.R.; Duarte dos Santos, C.N.; Bordignon, J. Dendritic cell apoptosis and the pathogenesis of dengue. Viruses 2012, 4, 2736–2753. [Google Scholar] [CrossRef]
- Stephens, H.A.F.; Klaythong, R.; Sirikong, M.; Vaughn, D.W.; Green, S.; Kalayanarooj, S.; Endy, T.P.; Libraty, D.H.; Nisalak, A.; Innis, B.L.; et al. HLA-A and -B allele associations with secondary dengue virus infections correlate with disease severity and the infecting viral serotype in ethnic Thais. Tissue Antigens 2002, 60, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Xavier Eurico De Alencar, L.; De Mendonça Braga-Neto, U.; Moura Do Nascimento, E.J.; Tenório Cordeiro, M.; Silva, A.M.; Antunes De Brito, C.A.; Carvalho Da Silva, M.D.P.; Vega Gonzales Gil, L.H.; Lucena Montenegro, S.M.; Torres De Azevedo Marques, E. HLA-B * 44 is associated with dengue severity caused by DENV-3 in a brazilian population. J. Trop. Med. 2013, 2013, 648475. [Google Scholar] [CrossRef] [PubMed]
- Chong, H.Y.; Leow, C.Y.; Majeed, A.; Bakar, A.; Leow, C.H. Flavivirus Infection-A Review of Immunopathogenesis, Immunological Response, and Immunodiagnosis. Virus Res. 2019, 274, 197770. [Google Scholar] [CrossRef] [PubMed]
- Begum, F.; Das, S.; Mukherjee, D.; Ray, U. Hijacking the Host Immune Cells by Dengue Virus: Molecular Interplay of Receptors and Dengue Virus Envelope. Microorganisms 2019, 7, 323. [Google Scholar] [CrossRef] [PubMed]
- Ngono, A.E.; Shresta, S. Immune Response to Dengue and Zika. Annu. Rev. Immunol. 2018, 36, 279–308. [Google Scholar] [CrossRef]
- Uno, N.; Ross, T.M. Dengue virus and the host innate immune response. Emerg. Microbes Infect. 2018, 7, 167. [Google Scholar] [CrossRef]
- Kurane, I.; Innis, B.L.; Nimmannitya, S.; Meager, A.; Janus, J.; Ennis, F.A. Activation of T Lymphocytes in Dengue Virus Infections. J. Clin. Investig. 1991, 2, 1473–1480. [Google Scholar] [CrossRef]
- St. John, A.; Rathore, A.P. Adaptive immune responses to primary and secondary dengue virus infections. Nat. Rev. Immunol. 2019, 19, 218–230. [Google Scholar] [CrossRef]
- Weiskopf, D.; Sette, A. T-Cell immunity to infection with dengue virus in humans. Front. Immunol. 2014, 5, 1–6. [Google Scholar] [CrossRef]
- Yauch, L.E.; Zellweger, R.M.; Kotturi, M.F.; Qutubuddin, A.; Sidney, J.; Peters, B.; Prestwood, T.R.; Sette, A.; Shresta, S. A Protective Role for Dengue Virus-Specific CD8 + T Cells. J. Immunol. 2009, 182, 4865–4873. [Google Scholar]
- De Matos, A.M.; Carvalho, K.I.; Rosa, D.S.; Villas-Boas, L.S.; da Silva, W.C.; Rodrigues, C.L.; Oliveira, O.M.; Levi, J.E.; Araujo, E.S.; Pannuti, C.S.; et al. CD8+ T Lymphocyte Expansion, Proliferation and Activation in Dengue Fever. PLoS Negl. Trop. Dis. 2015, 9, e0003520. [Google Scholar] [CrossRef] [PubMed]
- Halstead, S.B. Controversies in dengue pathogenesis. Paediatr. Int. Child Health 2012, 32, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Rothman, A. Immunity to dengue virus: A tale of original antigenic sin and tropical cytokine storms. Nat. Rev. Immunol. 2011, 11, 532–543. [Google Scholar] [CrossRef] [PubMed]
- Manlove, L.S.; Berquam-vrieze, K.E.; Kristen, E.; Williams, R.T.; Jenkins, M.K.; Farrar, A. Adaptive Immunity to Leukemia Is Inhibited by Cross-Reactive Induced Regulatory T Cells. J. Immunol. 2015, 195, 4028–4037. [Google Scholar] [CrossRef]
- Weiskopf, D.; Angelo, M.A.; Sidney, J.; Peters, B.; Shresta, S.; Sette, A. Immunodominance Changes as a Function of the Infecting Dengue Virus Serotype and Primary versus Secondary Infection. J. Virol. 2014. [Google Scholar] [CrossRef]
- Mongkolsapaya, J.; Dejnirattisai, W.; Xu, X.; Vasanawathana, S.; Tangthawornchaikul, N.; Chairunsri, A.; Sawasdivorn, S.; Duangchinda, T.; Dong, T.; Rowland-Jones, S.; et al. Original antigenic sin and apoptosis in the pathogenesis of dengue hemorrhagic fever. Nat. Med. 2003, 9, 921–927. [Google Scholar] [CrossRef]
- Srikiatkhachorn, A.; Mathew, A.; Rothman, A.L. Immune Mediated Cytokine Storm and Its Role in Severe Dengue. Semin. Immunopathol. 2017, 39, 563–574. [Google Scholar] [CrossRef]
- Reyes-Sandoval, A.; Ludert, J.E. The Dual Role of the Antibody Response Against the Flavivirus Non-Structural Protein 1 (NS1) in Protection and Immuno-Pathogenesis. Front. Immunol. 2019, 10, 1–6. [Google Scholar] [CrossRef]
- Malavige, G.N. Pathogenesis of vascular leak in dengue virus infection. Immunology 2017, 151, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Chareonsirisuthigul, T.; Kalayanarooj, S.; Ubol, S. Dengue virus (DENV) antibody-Dependent enhancement of infection upregulates the production of anti-inflammatory cytokines, but suppresses anti-DENV free radical and pro-Inflammatory cytokine production, in THP-1 cells. J. Gen. Virol. 2007, 88, 365–375. [Google Scholar] [CrossRef]
- Azeredo, E.L.; Zagne, S.M.O.; Alvarenga, A.R.; Nogueira, R.M.R.; Kubelka, C.F.; De Oliveira-Pinto, L.M. Activated Peripheral Lymphocytes with Increased Expression of Cell Adhesion Molecules and Cytotoxic Markers are Associated with Dengue Fever Disease. Memórias do Instituto Oswaldo Cruz 2006, 101, 437–449. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, T.; Setthapramote, C.; Kurosu, T.; Nishimura, M.; Asai, A.; Omokoko, M.D.; Pipattanaboon, C.; Pitaksajjakul, P.; Limkittikul, K.; Subchareon, A.; et al. Dengue virus neutralization and antibody-dependent enhancement activities of human monoclonal antibodies derived from dengue patients at acute phase of secondary infection. Antivir. Res. 2013, 98, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Mota, J.; Rico-Hesse, R. Dengue Virus Tropism in Humanized Mice Recapitulates Human Dengue Fever. PLoS ONE 2011, 6, e20762. [Google Scholar] [CrossRef]
- Onlamoon, N.; Noisakran, S.; Hsiao, H.M.; Duncan, A.; Villinger, F.; Ansari, A.A.; Perng, G.C. Dengue virus -Induced hemorrhage in a nonhuman primate model. Blood 2010, 115, 1823–1834. [Google Scholar] [CrossRef] [PubMed]
- Alves dos Santos, E.; Fink, K. Animal Models for Dengue and Zika Vaccine Development. Adv. Exp. Med. Biol. 2018, 1062, 215–239. [Google Scholar] [PubMed]
- Clark, K.B.; Onlamoon, N.; Hsiao, H.-M.; Perng, G.C.; Villinger, F. Can non-Human primates serve as models for investigating dengue disease pathogenesis? Front. Microbiol. 2013, 4, 305. [Google Scholar] [CrossRef]
- Zellweger, R.M.; Shresta, S. Mouse models to study dengue virus immunology and pathogenesis. Front. Immunol. 2014, 5, 151. [Google Scholar] [CrossRef]
- Smith, D.R.; Holbrook, M.R.; Gowen, B.B. Animal models of viral hemorrhagic fever. Antivir. Res. 2014, 112, 59–79. [Google Scholar] [CrossRef]
- Huang, K.J.; Li, S.Y.J.; Chen, S.C.; Liu, H.S.; Lin, Y.S.; Yeh, T.M.; Liu, C.C.; Lei, H.Y. Manifestation of thrombocytopenia in dengue-2-Virus-Infected mice. J. Gen. Virol. 2000, 81, 2177–2182. [Google Scholar] [CrossRef]
- Paes, M.V.; Pinhão, A.T.; Barreto, D.F.; Costa, S.M.; Oliveira, M.P.; Nogueira, A.C.; Takiya, C.M.; Farias-Filho, J.C.; Schatzmayr, H.G.; Alves, A.M.B.; et al. Liver injury and viremia in mice infected with dengue-2 virus. Virology 2005, 338, 236–246. [Google Scholar] [CrossRef]
- Chen, H.-C.; Hofman, F.M.; Kung, J.T.; Lin, Y.-D.; Wu-Hsieh, B.A. Both Virus and Tumor Necrosis Factor Alpha Are Critical for Endothelium Damage in a Mouse Model of Dengue Virus-Induced Hemorrhage. J. Virol. 2007, 81, 5518–5526. [Google Scholar] [CrossRef] [PubMed]
- Velandia-Romero, M.L.; Acosta-Losada, O.; Castellanos, J.E. In vivo infection by a neuroinvasive neurovirulent dengue virus. J. Neurovirol. 2012, 18, 374–387. [Google Scholar] [CrossRef] [PubMed]
- Yauch, L.E.; Shresta, S. Mouse models of dengue virus infection and disease. Antivir. Res. 2008, 80, 87–93. [Google Scholar] [CrossRef]
- Van den Broek, M.F.; Müller, U.; Huang, S.; Aguet, M.; Zinkernagel, R.M. Antiviral defense in mice lacking both alpha/beta and gamma interferon receptors. J. Virol. 1995, 69, 4792–4796. [Google Scholar] [CrossRef]
- Johnson, A.J.; Roehrig, J.T. New mouse model for dengue virus vaccine testing. J. Virol. 1999, 73, 783–786. [Google Scholar] [CrossRef] [PubMed]
- Tan, G.K.X.; Ng, J.K.W.; Lim, A.H.Y.; Yeo, K.P.; Angeli, V.; Alonso, S. Subcutaneous infection with non-Mouse adapted dengue virus D2Y98P strain induces systemic vascular leakage in AG129 mice. Ann. Acad. Med. Singap. 2011, 40, 523–532. [Google Scholar]
- Sarathy, V.V.; White, M.; Li, L.; Gorder, S.R.; Pyles, R.B.; Campbell, G.A.; Milligan, G.N.; Bourne, N.; Barrett, A.D.T. A Lethal Murine Infection Model for Dengue Virus 3 in AG129 Mice Deficient in Type I and II Interferon Receptors Leads to Systemic Disease. J. Virol. 2015, 89, 1254–1266. [Google Scholar] [CrossRef]
- Milligan, G.N.; Sarathy, V.V.; White, M.M.; Greenberg, M.B.; Campbell, G.A.; Pyles, R.B.; Barrett, A.D.T.; Bourne, N. A lethal model of disseminated dengue virus type 1 infection in AG129 mice. J. Gen. Virol. 2017, 98, 2507–2519. [Google Scholar] [CrossRef]
- Perry, S.T.; Buck, M.D.; Lada, S.M.; Schindler, C.; Shresta, S. STAT2 mediates innate immunity to dengue virus in the absence of STAT1 via the type I interferon receptor. PLoS Pathog. 2011, 7, e1001297. [Google Scholar] [CrossRef]
- Whitehead, S.S.; Blaney, J.E.; Durbin, A.P.; Murphy, B.R. Prospects for a dengue virus vaccine. Nat. Rev. Microbiol. 2007, 5, 518–528. [Google Scholar] [CrossRef]
- Chen, S.T.; Lin, Y.L.; Huang, M.T.; Wu, M.F.; Cheng, S.C.; Lei, H.Y.; Lee, C.K.; Chiou, T.W.; Wong, C.H.; Hsieh, S.L. CLEC5A is critical for dengue-Virus-Induced lethal disease. Nature 2008, 453, 672–676. [Google Scholar] [CrossRef] [PubMed]
- Carlin, A.F.; Plummer, E.M.; Vizcarra, E.A.; Sheets, N.; Joo, Y.; Tang, W.; Day, J.; Greenbaum, J.; Glass, C.K.; Diamond, M.S.; et al. An IRF-3-, IRF-5-, and IRF-7-Independent Pathway of Dengue Viral Resistance Utilizes IRF-1 to Stimulate Type I and II Interferon Responses. Cell Rep. 2017, 21, 1600–1612. [Google Scholar] [CrossRef] [PubMed]
- Marques, R.E.; Guabiraba, R.; Del Sarto, J.L.; Rocha, R.F.; Queiroz, A.L.; Cisalpino, D.; Marques, P.E.; Pacca, C.C.; Fagundes, C.T.; Menezes, G.B.; et al. Dengue virus requires the CC-Chemokine receptor CCR5 for replication and infection development. Immunology 2015, 145, 583–596. [Google Scholar] [CrossRef] [PubMed]
- Jhan, M.K.; HuangFu, W.C.; Chen, Y.F.; Kao, J.C.; Tsai, T.T.; Ho, M.R.; Shen, T.J.; Tseng, P.C.; Wang, Y.T.; Lin, C.F. Anti-TNF-α restricts dengue virus-Induced neuropathy. J. Leukoc. Biol. 2018, 104, 961–968. [Google Scholar] [CrossRef] [PubMed]
- Krishnqkumar, V.; Durairajan, S.S.K.; Alagarasu, K.; Li, M.; Dash, A.P. Recent Updates on Mouse Models for Human. Viruses 2019, 11, 252. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Chen, C.; Yang, Z.; Chi, X.; Zhang, J.; Chen, J.L. Targeted disruption of influenza A virus hemagglutinin in genetically modified mice reduces viral replication and improves disease outcome. Sci. Rep. 2016, 6, 23746. [Google Scholar] [CrossRef]
- Di Mario, G.; Garulli, B.; Sciaraffia, E.; Facchini, M.; Donatelli, I.; Castrucci, M.R. A heat-Inactivated H7N3 vaccine induces cross-Reactive cellular immunity in HLA-A2.1 transgenic mice. Virol. J. 2016, 13, 4–13. [Google Scholar] [CrossRef]
- Shresta, S.; Kyle, J.L.; Beatty, P.R.; Harris, E. Early activation of natural killer and B cells in response to primary dengue virus infection in A/J mice. Virology 2004, 319, 262–273. [Google Scholar] [CrossRef]
- Shresta, S.; Kyle, J.L.; Snider, H.M.; Basavapatna, M.; Beatty, P.R.; Harris, E. Interferon-Dependent Immunity Is Essential for Resistance to Primary Dengue Virus Infection in Mice, Whereas T- and B-Cell-Dependent Immunity Are Less Critical. J. Virol. 2004, 78, 2701–2710. [Google Scholar] [CrossRef]
- Shresta, S.; Sharar, K.L.; Prigozhin, D.M.; Beatty, P.R.; Harris, E. Murine Model for Dengue Virus-Induced Lethal Disease with Increased Vascular Permeability. J. Virol. 2006, 80, 10208–10217. [Google Scholar] [CrossRef]
- Prestwood, T.R.; Morar, M.M.; Zellweger, R.M.; Miller, R.; May, M.M.; Yauch, L.E.; Lada, S.M.; Shresta, S. Gamma Interferon (IFN-) Receptor Restricts Systemic Dengue Virus Replication and Prevents Paralysis in IFN-/ Receptor-Deficient Mice. J. Virol. 2012, 86, 12561–12570. [Google Scholar] [CrossRef]
- Schul, W.; Liu, W.; Xu, H.; Flamand, M.; Vasudevan, S.G. A Dengue Fever Viremia Model in Mice Shows Reduction in Viral Replication and Suppression of the Inflammatory Response after Treatment with Antiviral Drugs. J. Infect. Dis. 2007, 195, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Tan, G.K.; Ng, J.K.W.; Trasti, S.L.; Schul, W.; Yip, G.; Alonso, S. A non mouse-Adapted dengue virus strain as a new model of severe dengue infection in AG129 mice. PLoS Negl. Trop. Dis. 2010, 4, e672. [Google Scholar] [CrossRef]
- Sarathy, V.V.; Infante, E.; Li, L.; Campbell, G.A.; Wang, T.; Paessler, S.; Beatty, P.R.; Harris, E.; Milligan, G.N.; Bourne, N.; et al. Characterization of lethal dengue virus type 4 (DENV-4) TVP-376 infection in mice lacking both IFN-a/b and IFN-c receptors (AG129) and comparison with the DENV-2 AG129 mouse model. J. Gen. Virol. 2015, 96, 3035–3048. [Google Scholar] [CrossRef] [PubMed]
- Sarathy, V.V.; White, M.; Li, L.; Kaiser, J.A.; Campbell, G.A. Characterization of a murine model of non-Lethal, symptomatic dengue virus infection. Sci. Rep. 2018, 8, 4900. [Google Scholar] [CrossRef]
- Milligan, G.N.; Sarathy, V.V.; Infante, E.; Li, L.; Campbell, G.A.; Beatty, P.R.; Harris, E.; Barrett, A.D.T.; Bourne, N. A dengue virus type 4 model of disseminated lethal infection in AG129 mice. PLoS ONE 2015, 10, e0125476. [Google Scholar] [CrossRef]
- Orozco, S.; Schmid, M.A.; Parameswaran, P.; Lachica, R.; Henn, M.R.; Beatty, R.; Harris, E. Characterization of a model of lethal dengue virus 2 infection in C57BL/6 mice deficient in the alpha/beta interferon receptor. J. Gen. Virol. 2012, 93, 2152–2157. [Google Scholar] [CrossRef]
- María, J.; Gómez, M.; Ong, L.C.; Lam, J.H.; Binte, S.A. Maternal Antibody-Mediated Disease Enhancement in Type I Interferon-Deficient Mice Leads to Lethal Disease Associated with Liver Damage. PLoS Negl. Trop. Dis. 2016, 10, e0004536. [Google Scholar]
- Zust, R.; Toh, Y.-X.; Valdes, I.; Cerny, D.; Heinrich, J.; Hermida, L.; Marcos, E.; Guillen, G.; Kalinke, U.; Shi, P.-Y.; et al. Type I Interferon Signals in Macrophages and Dendritic Cells Control Dengue Virus Infection: Implications for a New Mouse Model To Test Dengue Vaccines. J. Virol. 2014, 88, 7276–7285. [Google Scholar] [CrossRef]
- Perry, S.T.; Prestwood, T.R.; Lada, S.M.; Benedict, C.A.; Shresta, S. Cardif-Mediated Signaling Controls the Initial Innate Response to Dengue Virus In Vivo. J. Virol. 2009, 83, 8276–8281. [Google Scholar] [CrossRef]
- Shresta, S.; Sharar, K.L.; Prigozhin, D.M.; Snider, H.M.; Beatty, P.R.; Harris, E. Critical Roles for Both STAT1-Dependent and STAT1-Independent Pathways in the Control of Primary Dengue Virus Infection in Mice. J. Immunol. 2005, 175, 3946–3954. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.L.; Liao, C.L.; Liu, C.I.; Ma, S.H.; Huang, Y.Y.; Huang, Y.L.; Chen, L.K.; Yeh, C.T.; Kao, C.L.; King, C.C. Study of dengue virus infection in SCID mice engrafted with human K562 cells. J. Virol. 1998, 72, 9729–9737. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Kimura-Kuroda, J.; Hirabayashi, Y.; Yasui, K. Development of a novel mouse model for dengue virus infection. Virology 1999, 263, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Blaney, J.E.; Matro, J.M.; Murphy, B.R.; Whitehead, S.S. Recombinant, Live-Attenuated Tetravalent Dengue Virus Vaccine Formulations Induce a Balanced, Broad, and Protective Neutralizing Antibody Response against Each of the Four Serotypes in Rhesus Monkeys. J. Virol. 2005, 79, 5516–5528. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.J.; Hayes, C.G.; Dubois, D.R.; Windheuser, M.G.; Kang, Y.H.; Watts, D.M.; Sieckmann, D.G. Evaluation of the severe combined immunodeficient (SCID) mouse as an animal model for dengue viral infection. Am. J. Trop. Med. Hyg. 1995, 52, 468–476. [Google Scholar] [CrossRef] [PubMed]
- Bente, D.A.; Melkus, M.W.; Garcia, J.V.; Rico-Hesse, R. Dengue Fever in Humanized NOD/SCID Mice. J. Virol. 2005, 79, 13797–13799. [Google Scholar] [CrossRef] [PubMed]
- Mota, J.; Rico-Hesse, R. Humanized Mice Show Clinical Signs of Dengue Fever according to Infecting Virus Genotype. J. Virol. 2009, 83, 8638–8645. [Google Scholar] [CrossRef]
- Sridharan, A.; Chen, Q.; Tang, K.F.; Ooi, E.E.; Hibberd, M.L.; Chen, J. Inhibition of Megakaryocyte Development in the Bone Marrow Underlies Dengue Virus-Induced Thrombocytopenia in Humanized Mice. J. Virol. 2013, 87, 11648–11658. [Google Scholar] [CrossRef]
- Costa, V.V.; Ye, W.; Qingfeng, C.; Teixeira, M.M.; Preiser, P.; Ooi, E.E.; Chena, J. Dengue Virus-Infected Dendritic Cells but Not Monocytes, Activate Natural Killer Cells through a Contact-Dependent. MBio 2017, 8, e00741-17. [Google Scholar] [CrossRef]
- Jaiswal, S.; Pazoles, P.; Woda, M.; Shultz, L.D.; Greiner, D.L.; Brehm, M.A.; Mathew, A. Enhanced humoral and HLA-A2-Restricted dengue virus-Specific T-Cell responses in humanized BLT NSG mice. Immunology 2012, 136, 334–343. [Google Scholar] [CrossRef]
- Frias-Staheli, N.; Dorner, M.; Marukian, S.; Billerbeck, E.; Labitt, R.N.; Rice, C.M.; Ploss, A. Utility of Humanized BLT Mice for Analysis of Dengue Virus Infection and Antiviral Drug Testing. J. Virol. 2014, 88, 2205–2218. [Google Scholar] [CrossRef] [PubMed]
- Mathew, A.; Townsley, E.; Ennis, F.A. Elucidating the role of T cells in protection against and pathogenesis of dengue virus infections. Future Microbiol. 2014, 9, 411–425. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, S.; Pearson, T.; Friberg, H.; Shultz, L.D.; Greiner, D.L.; Rothman, A.L.; Mathew, A. Dengue virus infection and virus-specific HLA-A2 restricted immune responses in humanized NOD-scid IL2rγnull mice. PLoS ONE 2009, 4, e7251. [Google Scholar] [CrossRef] [PubMed]
- Taconic Biosciences. Comparison Guide: huNOG-EXL, NSG-SGM3 AND MISTRG. 2016. Available online: https://www.taconic.com/taconic-insights/immunology/comparison-guide-hunog-exl-nsg-sgm3-and-mistrg.html (accessed on 20 September 2019).
- Jangalwe, S.; Shultz, L.D.; Mathew, A.; Brehm, M.A. Improved B cell development in humanized NOD-scid IL2Rγ null mice transgenically expressing human stem cell factor, granulocyte-macrophage colony-Stimulating factor and interleukin-3. Immun. Inflamm. Dis. 2016, 4, 427–440. [Google Scholar] [CrossRef]
- Douam, F.; Ziegler, C.G.K.; Hrebikova, G.; Fant, B.; Leach, R.; Parsons, L.; Wang, W.; Gaska, J.M.; Winer, B.Y.; Heller, B.; et al. Selective expansion of myeloid and NK cells in humanized mice yields human-Like vaccine responses. Nat. Commun. 2018, 9, 5031. [Google Scholar] [CrossRef]
- Kuruvilla, J.G.; Troyer, R.M.; Devi, S.; Akkina, R. Dengue virus infection and immune response in humanized RAG2−/−γc−/− (RAG-hu) mice. Virology 2007, 369, 143–152. [Google Scholar] [CrossRef]
- Danner, R.; Chaudhari, S.N.; Rosenberger, J.; Surls, J.; Richie, T.L.; Brumeanu, T.D.; Casares, S. Expression of HLA class II molecules in humanized NOD.Rag1KO.IL2RgcKO mice is critical for development and function of human T and B cells. PLoS ONE 2011, 6, e19826. [Google Scholar] [CrossRef]
- Cox, J.; Mota, J.; Sukupolvi-Petty, S.; Diamond, M.S.; Rico-Hesse, R. Mosquito Bite Delivery of Dengue Virus Enhances Immunogenicity and Pathogenesis in Humanized Mice. J. Virol. 2012, 86, 7637–7649. [Google Scholar] [CrossRef]
- Osorio, J.E.; Partidos, C.D.; Wallace, D.; Stinchcomb, D.T. Development of a recombinant, chimeric tetravalent dengue vaccine candidate. Vaccine 2015, 33, 7112–7120. [Google Scholar] [CrossRef]
- Clements, D.; Coller, B.; Lieberman, M.; Ogata, S.; Wang, G.; Harada, K.; Putnak, J.; Ivy, J.; McDonell, M.; Bignami, G.; et al. Development of a recombinant tetravalent dengue virus vaccine: Immunogenicity and efficacy studies in mice and monkeys. Vaccine 2010, 28, 2705–2715. [Google Scholar] [CrossRef]
- Sabin, A.B. Research on Dengue during World War II. Am. J. Trop. Med. Hyg. 1952, 1, 30–50. [Google Scholar] [CrossRef] [PubMed]
- Cole, G.A.; Wisseman, C.L.; Nathanson, N. Pathogenesis of type 1 dengue virus infection in suckling, weaned and adult mice. Am. J. Epidemiol. 1973, 83, 243–252. [Google Scholar] [CrossRef]
- White, L.J.; Parsons, M.M.; Whitmore, A.C.; Williams, B.M.; de Silva, A.; Johnston, R.E. An Immunogenic and Protective Alphavirus Replicon Particle-Based Dengue Vaccine Overcomes Maternal Antibody Interference in Weanling Mice. J. Virol. 2007, 81, 10329–10339. [Google Scholar] [CrossRef] [PubMed]
- Mellado-Sánchez, G.; García-Cordero, J.; Luria-Pérez, R.; Lázaro-Olan, L.; Santos-Argumedo, L.; Gutiérrez-Castañeda, B.; Estrada-García, I.; Cedillo-Barrón, L. DNA priming E and NS1 constructs-Homologous proteins boosting immunization strategy to improve immune response against dengue in mice. Viral Immunol. 2005, 18, 709–721. [Google Scholar] [CrossRef] [PubMed]
- Mellado-Sánchez, G.; García-Machorro, J.; Sandoval-Montes, C.; Gutiérrez-Castañeda, B.; Rojo-Domínguez, A.; García-Cordero, J.; Santos-Argumedo, L.; Cedillo-Barrón, L. A plasmid encoding parts of the dengue virus E and NS1 proteins induces an immune response in a mouse model. Arch. Virol. 2010, 155, 847–856. [Google Scholar] [CrossRef]
- Blaney, J.E.; Johnson, D.H.; Firestone, C.-Y.; Hanson, C.T.; Murphy, B.R.; Whitehead, S.S. Chemical Mutagenesis of Dengue Virus Type 4 Yields Mutant Viruses Which Are Temperature Sensitive in Vero Cells or Human Liver Cells and Attenuated in Mice. J. Virol. 2001, 75, 9731–9740. [Google Scholar] [CrossRef][Green Version]
- Huang, C.Y.-H.; Butrapet, S.; Tsuchiya, K.R.; Bhamarapravati, N.; Gubler, D.J.; Kinney, R.M. Dengue 2 PDK-53 Virus as a Chimeric Carrier for Tetravalent Dengue Vaccine Development. J. Virol. 2003, 77, 11436–11447. [Google Scholar] [CrossRef]
- Brewooa, J.N.; Kinney, R.M.; Powell, T.D.; Arguello, J.J.; Silengo, S.J.; Partidos, C.D.; Huang, C.Y.-H.; Stinchcomb, D.T.; Osorio, J.E. Immunogenicity and efficacy of chimeric dengue vaccine (DENVax) formulations in interferon-Deficient AG129 mice. Physiol. Behav. 2017, 176, 139–148. [Google Scholar] [CrossRef]
- Zellweger, R.M.; Miller, R.; Eddy, W.E.; White, L.J.; Johnston, R.E.; Shresta, S. Role of Humoral versus Cellular Responses Induced by a Protective Dengue Vaccine Candidate. PLoS Pathog. 2013, 9, e1003723. [Google Scholar] [CrossRef]
- Züst, R.; Dong, H.; Li, X.-F.; Chang, D.C.; Zhang, B.; Balakrishnan, T.; Toh, Y.X.; Jiang, T.; Li, S.H.; Deng, Y.Q.; et al. Rational Design of a Live Attenuated Dengue Vaccine: 2′-O-Methyltransferase Mutants Are Highly Attenuated and Immunogenic in Mice and Macaques. PLoS Pathog. 2013, 9, e1003521. [Google Scholar] [CrossRef]
- Wijayalath, W.; Majji, S.; Villasante, E.F.; Brumeanu, T.D.; Richie, T.L.; Casares, S. Humanized HLA-DR4.RagKO.IL2RγcKO.NOD (DRAG) mice sustain the complex vertebrate life cycle of Plasmodium falciparum malaria. Malar. J. 2014, 13, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Allam, A.; Majji, S.; Peachman, K.; Jagodzinski, L.; Kim, J.; Ratto-Kim, S.; Wijayalath, W.; Merbah, M.; Kim, J.H.; Michael, N.L.; et al. TFH cells accumulate in mucosal tissues of humanized-DRAG mice and are highly permissive to HIV-1. Sci. Rep. 2015, 5, 10443. [Google Scholar] [CrossRef] [PubMed]
- Racki, W.J.; Covassin, L.; Brehm, M.; Pino, S.; Ignotz, R.; Dunn, R.; Laning, J.; Graves, S.K.; Rossini, A.A.; Shultz, L.D.; et al. NOD-Scid IL2rγnull mouse model of human skin transplantation and allograft rejection. Transplantation 2010, 89, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Klicznik, M.M.; Benedetti, A.; Gail, L.M.; Holly, R.; Laimer, M.; Stoecklinger, A.; Sir, A.; Reitsamer, R.; Rosenblum, M.D.; Murauer, E.M.; et al. A novel humanized mouse model to study human antigen-Specific cutaneous T cell responses in vivo. BioRxlv 2018, 1–36. [Google Scholar]
- Yong, K.S.M.; Her, Z.; Chen, Q. Humanized Mouse Models for the Study of Hepatitis C and Host Interactions. Cells 2019, 8, 604. [Google Scholar] [CrossRef] [PubMed]
- Zellweger, R.M.; Prestwood, T.R.; Shresta, S. Antibodies enhance infection of LSECs in a model of ADE-induced severe dengue disease. Cell Host Microbe 2010, 7, 128–139. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Mouse Model | Route | Asymptomatic Infection | Neurologic Signs | Hematological Changes | Disease Signs | Pro-Inflammatory Cytokines | Ref. |
|---|---|---|---|---|---|---|---|
| BALB/c | IP, I.C. | NR | Yes | Yes | NR | [74] | |
| A/J | IV | NR | Yes | Transient thrombocytopenia | NR | NR | [71] |
| IV | Yes | Yes | Transient ↑ hematocrit and ↓ WBC count | NR | NR | [90] | |
| IV | NR | Yes | Hematocrit: Slight ↑ | NR | NR | [91] | |
| IV | NR | NR | NR | Vascular permeability: High | NR | [92] | |
| IV | NR | Yes | NR | Yes | IFN-γ: ↑ | [93] | |
| IP | NR | NR | NR | Yes | IFN-γ, IL-6, IL-12, TNF-α: ↑ | [94] | |
| IP | Yes | Yes | ↑ levels of transaminases | Vascular leakage: High | IFN-γ, IL-6 and TNF-α: ↑ | [77,95] | |
| IP | Yes | Yes | Early leukopenia | Yes | IL-1α, IL-6, IL-12p40, IL-17A, IFN-γ and TNF-α: ↑ | [96] | |
| IP | NR | NR | Thrombocytopenia | Plasma leakage | IL-2, IL-4, and TNFα: ↑ | [97] | |
| IP | NR | NR | Coagulopathy | Plasma leakage | IL-2, IL-4, and TNFα: ↑ | ||
| IP | NR | NR | Thrombocytopenia | Plasma leakage | IL-1α, IL-6, IL-10, IL-12p40, IFN-γ and G-CSF: ↑ | [80] | |
| IP | NR | Yes | Leukopenia Thrombocytopenia | Yes | IL-1α, IL-6, IL-10, IL-12p40, IFN-γ, and G-CSF: ↑ | [98] | |
| A129 | IV | NR | Yes | NR | Yes | IFN-γ: ↑ | [93] |
| IV | NR | Yes | NR | Yes | TNF-α and IL-10: ↑ | [99] | |
| IV | NR | NR | Thrombocytopenia and Lymphopenia: Moderate AST: Elevated | Severe liver damage Vascular leakage Middle | DENV-2: IL-6: ↑ | [100] | |
| IFNAR−/− | IV | NR | Yes | NR | NR | TNF-α and IL-10: ↑ | [99] |
| IV | NR | NR | NR | Yes | NR | [52] | |
| IP | NR | NR | NR | Yes | IL-6, INF-α, IP10, IFN-a, IFN-γ: ↑ | [101] | |
| Cardif−/− C57BL/6 | IV | Yes | NR | NR | NR | IFN-α: ↑ | [102] |
| STAT1−/− | IV | NR | NR | NR | NR | IFNα: ↓ IFNβ: ↑ | [81] |
| IC | NR | Yes | NR | Yes | NR | [103] | |
| STAT2−/− | IV | NR | NR | NR | IFNα: ↓ IFNβ: ↑ | [81] | |
| STAT1−/− STAT2−/−and STAT1−/− IFNAR−/− | IV | NR | NR | NR | NR | IFNα: ↓ IFNβ: ↑ |
| Mouse Model | Route | Hematological Changes | Disease | Immune Response | Ref | ||
|---|---|---|---|---|---|---|---|
| Pro-Inflammatory Cytokines | Cellular Response | Humoral Response | |||||
NSG mice also known as:
| ID | Thrombocytopenia: High | Yes | IFN-γ, TNF-α, IL-2 and VEGF: ↑ | CD4 Th1, TH2 and B cells | NR | [65] |
| ID, Mosquito | Thrombocytopenia | NR | IFN-γ, IL-4, Il-10: ↑ | NR | IgM, IgG, IgA | [121] | |
| SC | Thrombocytopenia | Yes | NR | NR | IgM, IgG, IgA | [109] | |
| IV | Thrombocytopenia: High hTPO; Middle Megakaryocytes: ↓ | NR | NR | NR | NR | [110] | |
| SC, IP | NR | NR | NR | CD4+ | IgM | [115] | |
| NSG-BLT mice (HLA-A2) mice | SC | NR | Yes | IFN-γ: ↑ | CD8+ | IgM | [112] |
| NSG-BLT | IV | Thrombocytopenia: (7 or 14 dpi) | Yes | IL-R1A, VEGF, IP10: ↑ (3dpi) | CD4+, CD8+ | IgM | [113] |
| SCID-HuH-7 | IP | NR | NR | NR | NR | Neutralizing Antibodies | [106] |
| RAG2−/− gamma c−/− | SC | NR | Yes | NR | NR | IgM, IgG | [119] |
| NSG-Tg (HLA-A2/H2-D/beta2M)1DVs/Sz mice also known as: -NOD-scid IL2rgamma null TG (HLA-A2/HuBeta2M) | SC | NR | Yes | IFNγ, TNFα, IL-2: ↑ | NR | IgM, IgG, IgA | [115] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coronel-Ruiz, C.; Gutiérrez-Barbosa, H.; Medina-Moreno, S.; Velandia-Romero, M.L.; Chua, J.V.; Castellanos, J.E.; Zapata, J.C. Humanized Mice in Dengue Research: A Comparison with Other Mouse Models. Vaccines 2020, 8, 39. https://doi.org/10.3390/vaccines8010039
Coronel-Ruiz C, Gutiérrez-Barbosa H, Medina-Moreno S, Velandia-Romero ML, Chua JV, Castellanos JE, Zapata JC. Humanized Mice in Dengue Research: A Comparison with Other Mouse Models. Vaccines. 2020; 8(1):39. https://doi.org/10.3390/vaccines8010039
Chicago/Turabian StyleCoronel-Ruiz, Carolina, Hernando Gutiérrez-Barbosa, Sandra Medina-Moreno, Myriam L. Velandia-Romero, Joel V. Chua, Jaime E. Castellanos, and Juan C. Zapata. 2020. "Humanized Mice in Dengue Research: A Comparison with Other Mouse Models" Vaccines 8, no. 1: 39. https://doi.org/10.3390/vaccines8010039
APA StyleCoronel-Ruiz, C., Gutiérrez-Barbosa, H., Medina-Moreno, S., Velandia-Romero, M. L., Chua, J. V., Castellanos, J. E., & Zapata, J. C. (2020). Humanized Mice in Dengue Research: A Comparison with Other Mouse Models. Vaccines, 8(1), 39. https://doi.org/10.3390/vaccines8010039