Novel Nested Peptide Epitopes Recognized by CD4+ T Cells Induced by HIV-1 Conserved-Region Vaccines

Abstract
1. Introduction
2. Materials and Methods
2.1. Ttial HIV-CORE 002l
2.2. HLA Typing
2.3. Peptides
2.4. Short-Term Cell Lines (STCL)
2.5. IFN-γ ELISPOT Assay
2.6. Intracellular Cytokine Staining and Flow Cytometry
2.7. Epitope Prediction
3. Results
3.1. The Study Subjects and Vaccination
3.2. CD4+ T-Cell Epitope Mapping
HC003 EVIPMFTALSEGATP (Gag 167–181)
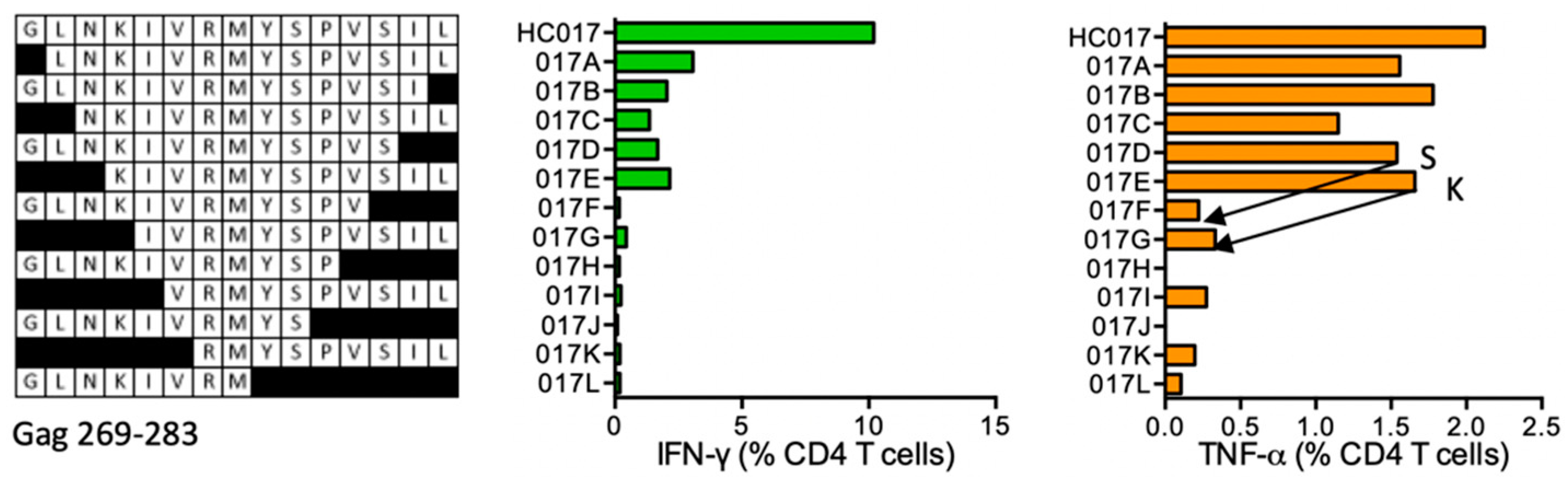
HC017 GLNKIVRMYSPVSIL (Gag 269–283)
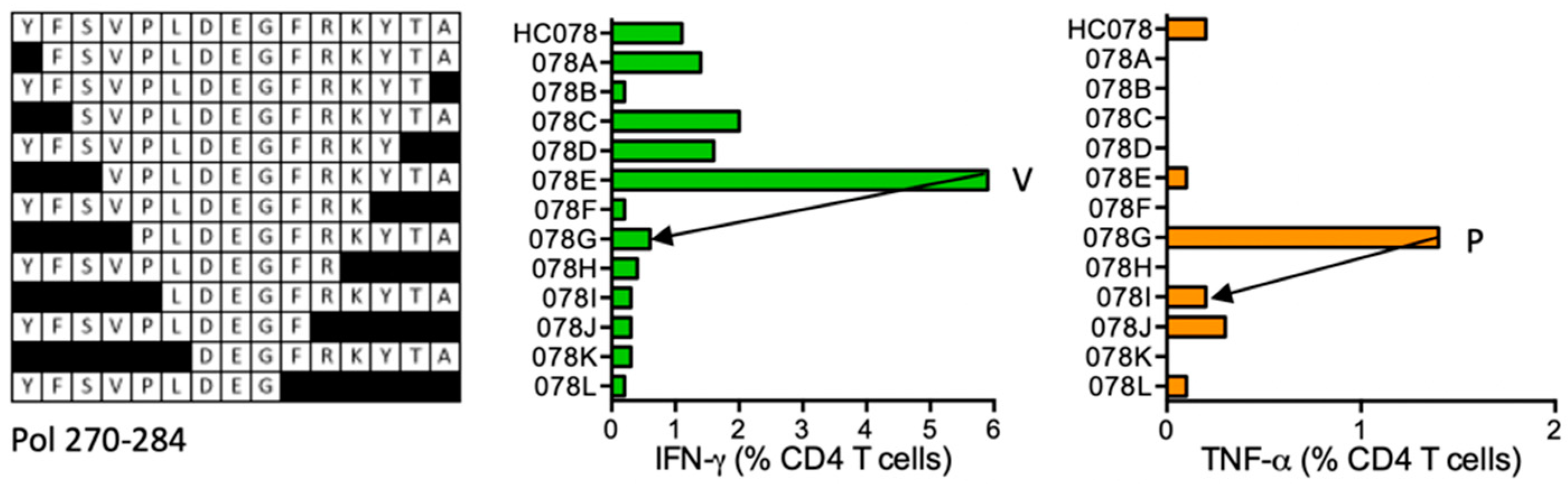
HC078 YFSVPLDEGFRKYTA (Pol 270–284)
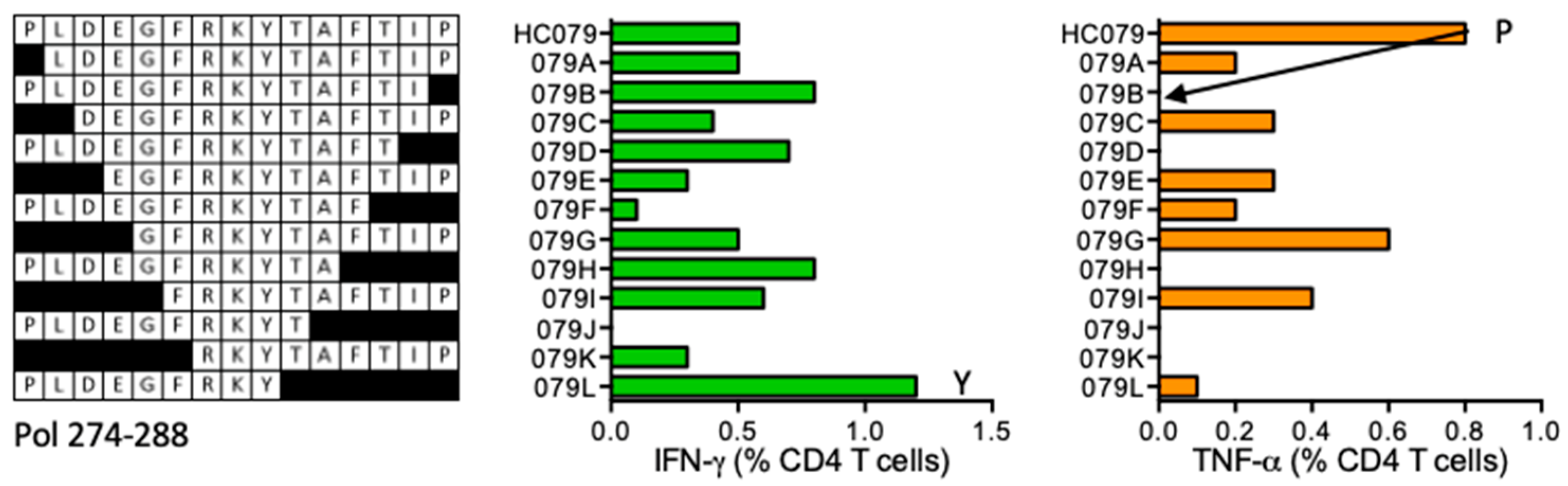
HC079 PLDEGFRKYTAFTIP (Pol 274–288)
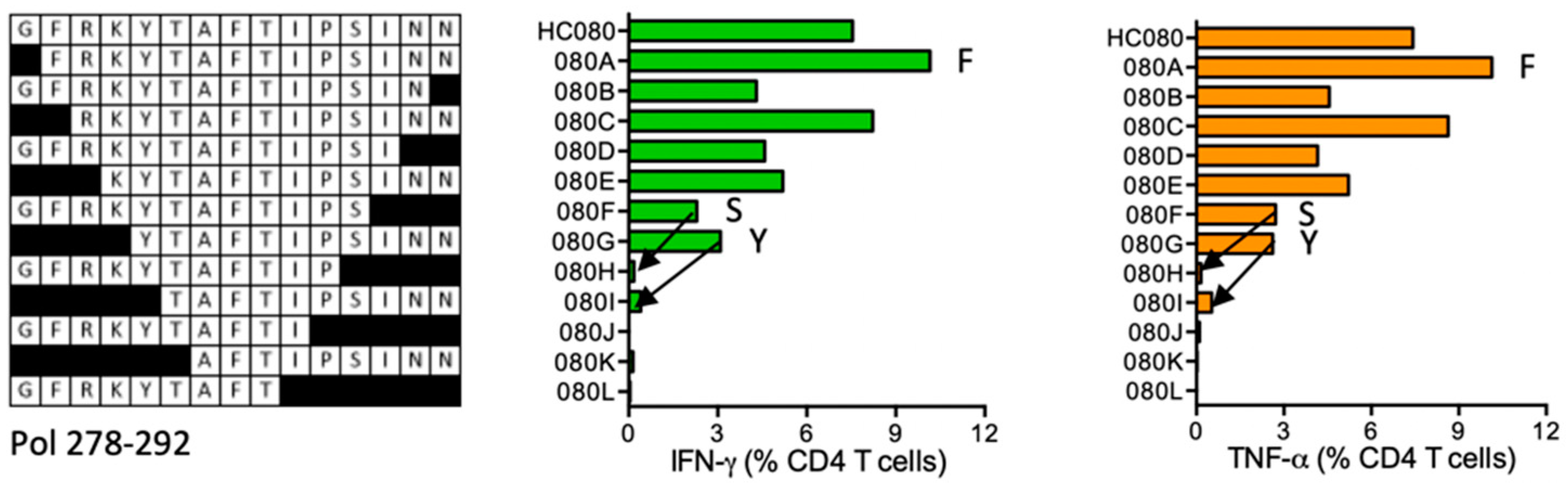
HC080 GFRKYTAFTIPSINN (Pol 278–292)
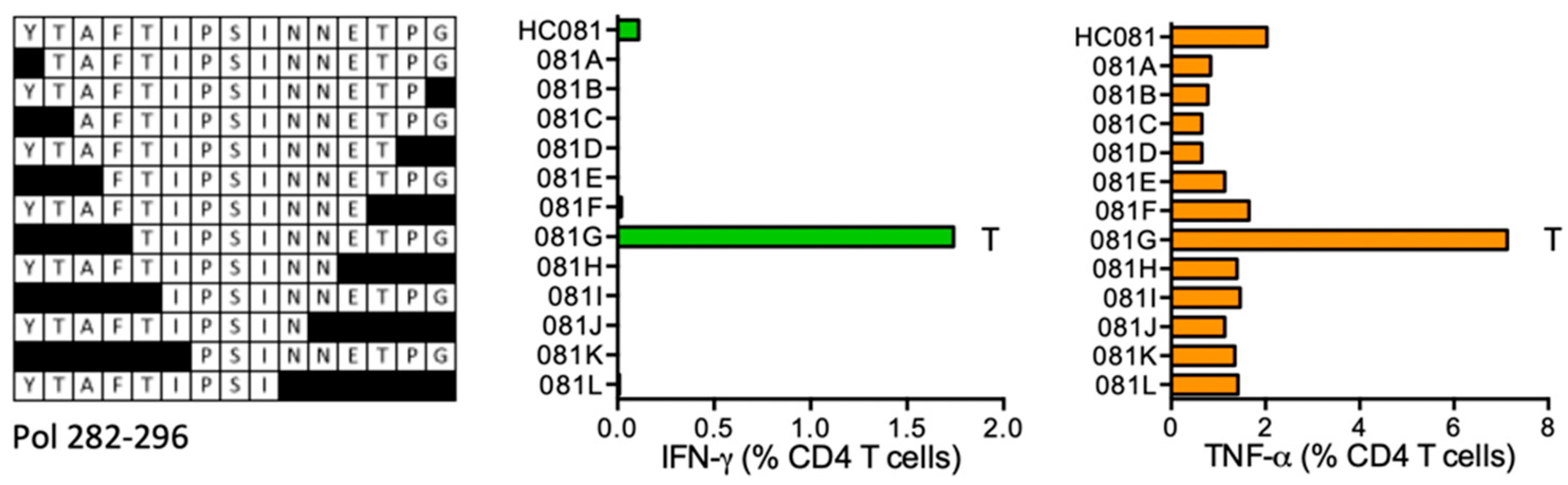
HC081 YTAFTIPSINNETPG (Pol 282–296)
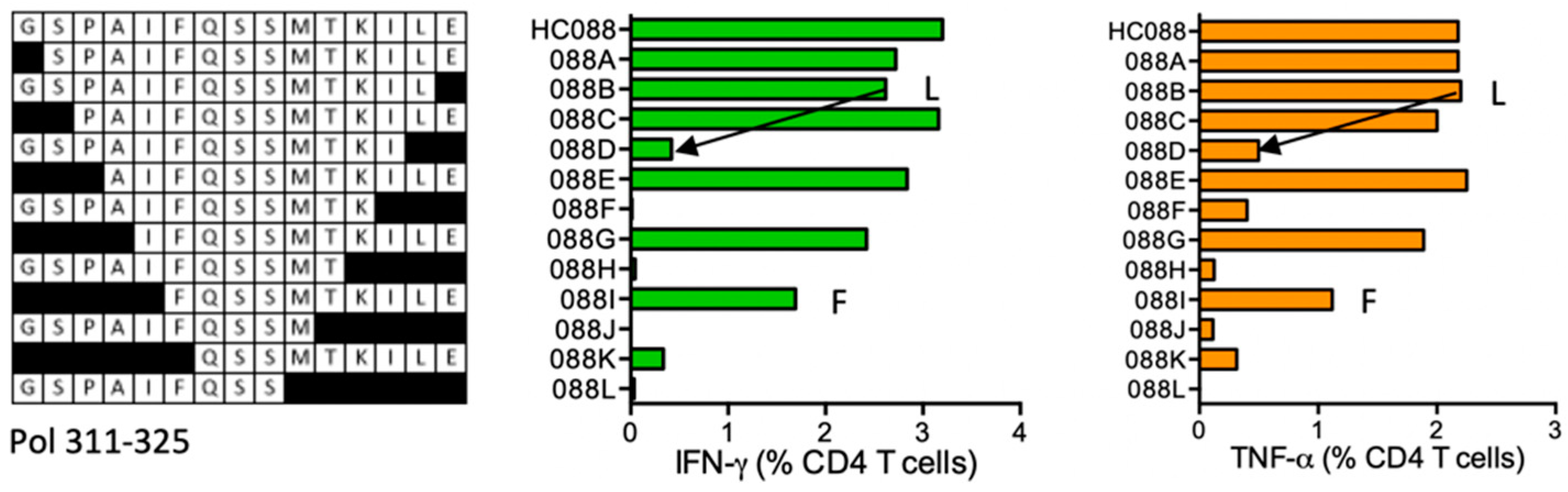
HC088 GSPAIFQSSMTKILE- Pol (311–325)
HC093 KNPEIVIYQYMDDLYV (Pol 330–344; K was added for solubility)
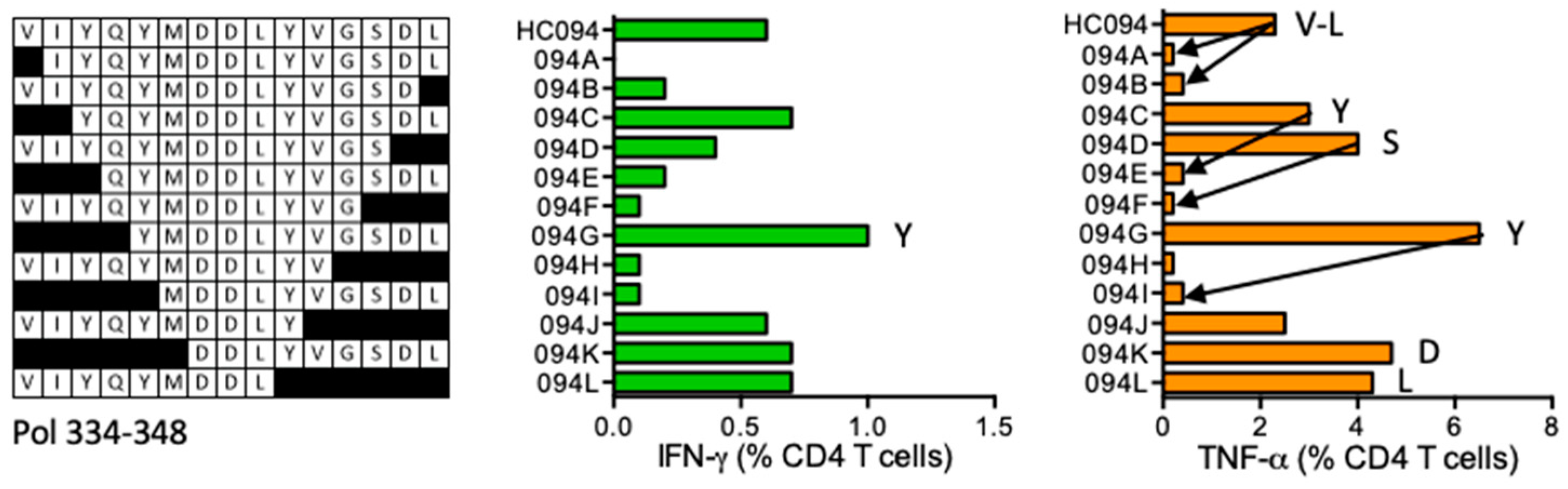
HC094 VIYQYMDDLYVGSDL (Pol 334–348)
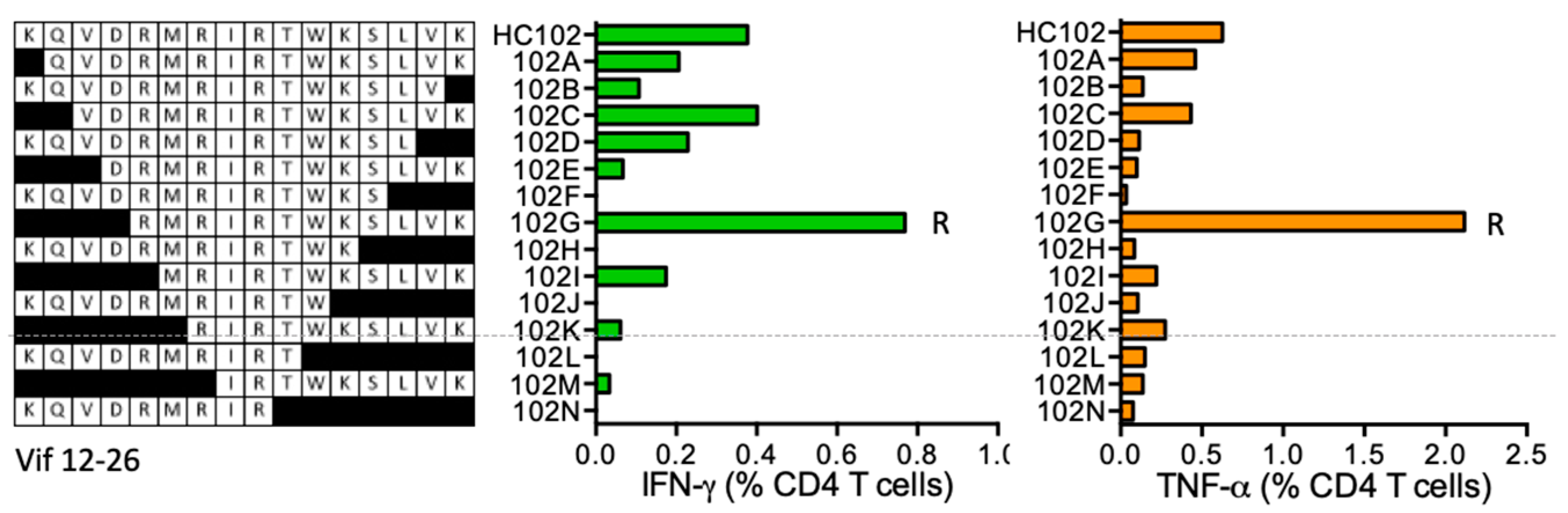
HC102 KQVDRMRIRTWKSLVK (Vif 12–26; K was added for solubility)
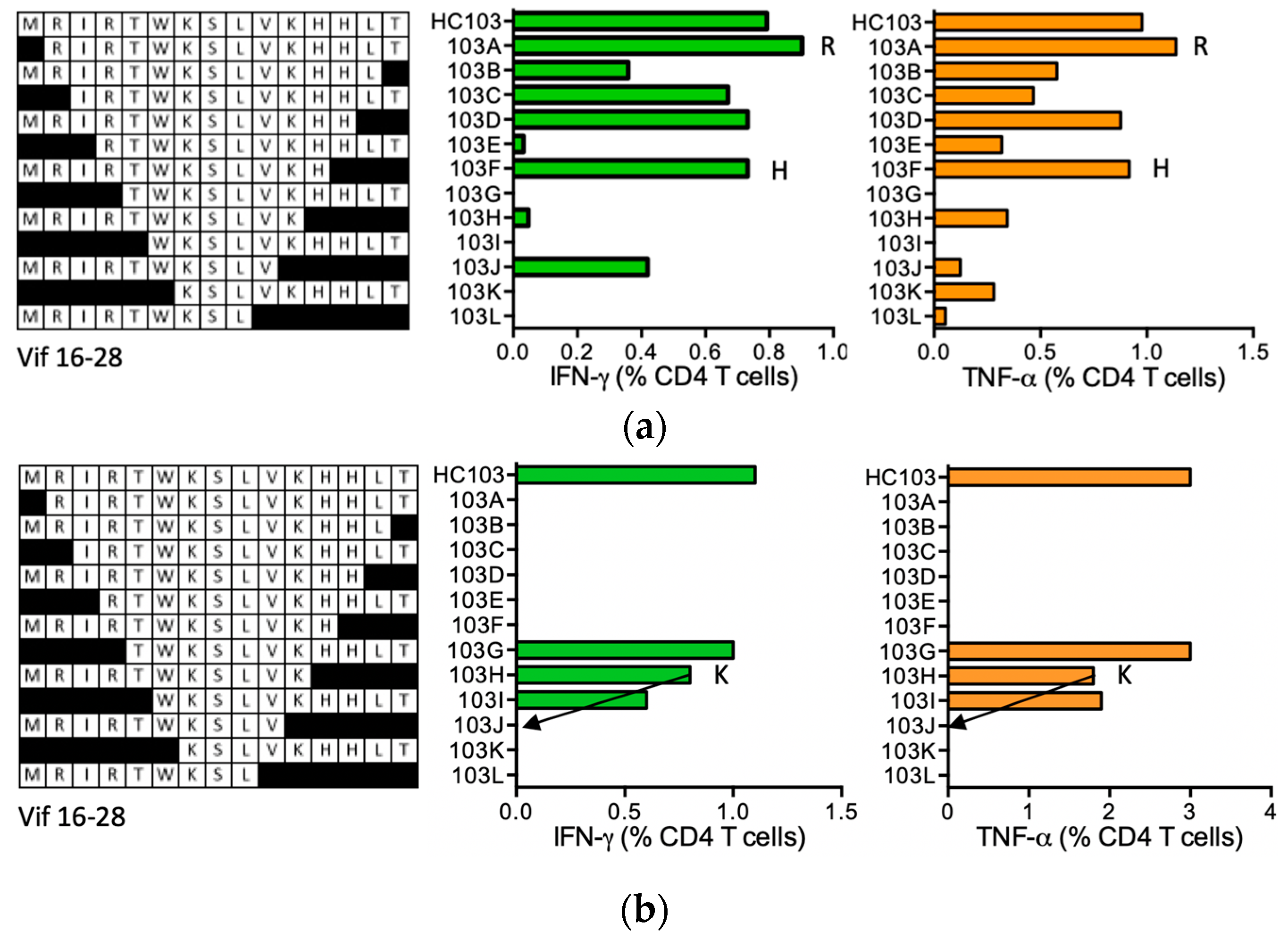
HC103 MRIRTWKSLVKHH-LT (Vif 16–28; ‘-’ indicates junction between two adjacent regions)
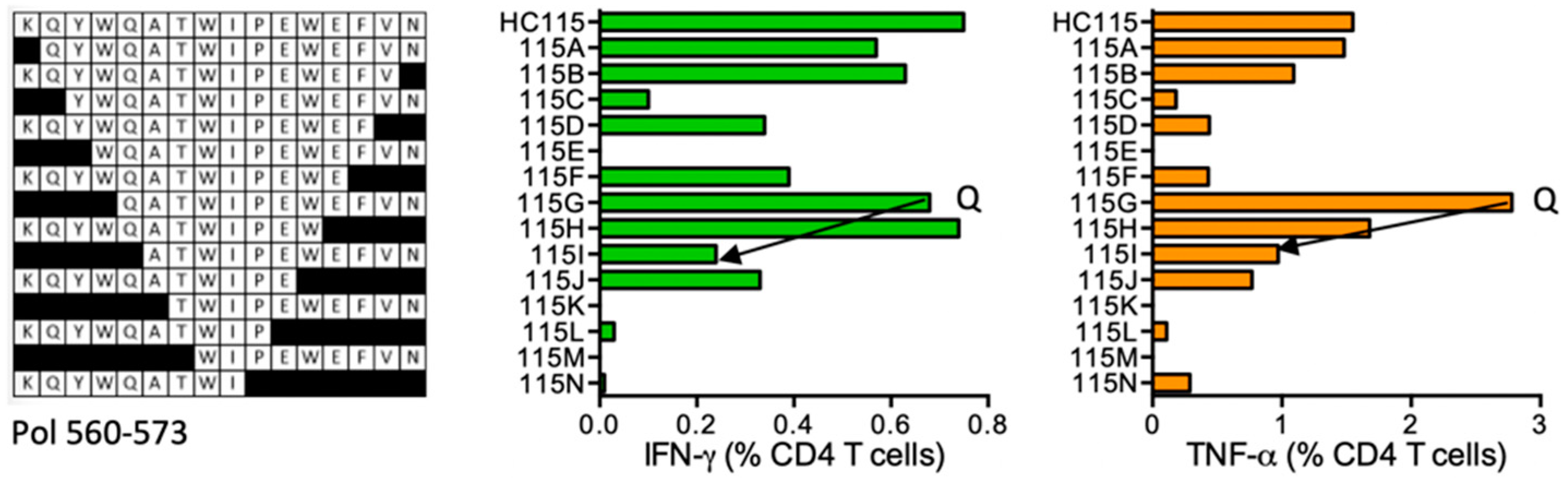
HC115 KQ-YWQATWIPEWEFVN (Pol 560–573; K was added for solubility; ‘-’ indicates junction between two adjacent regions)
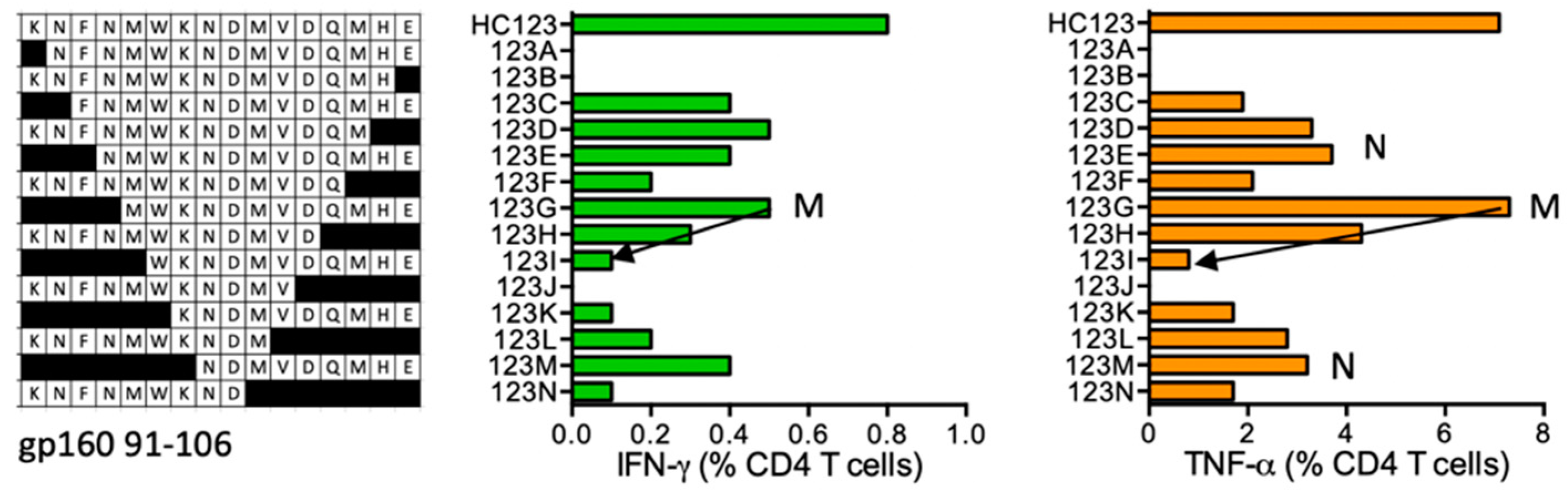
HC123 KNFNMWKNDMVDQMHE (gp160 91–106; K was added for solubility)
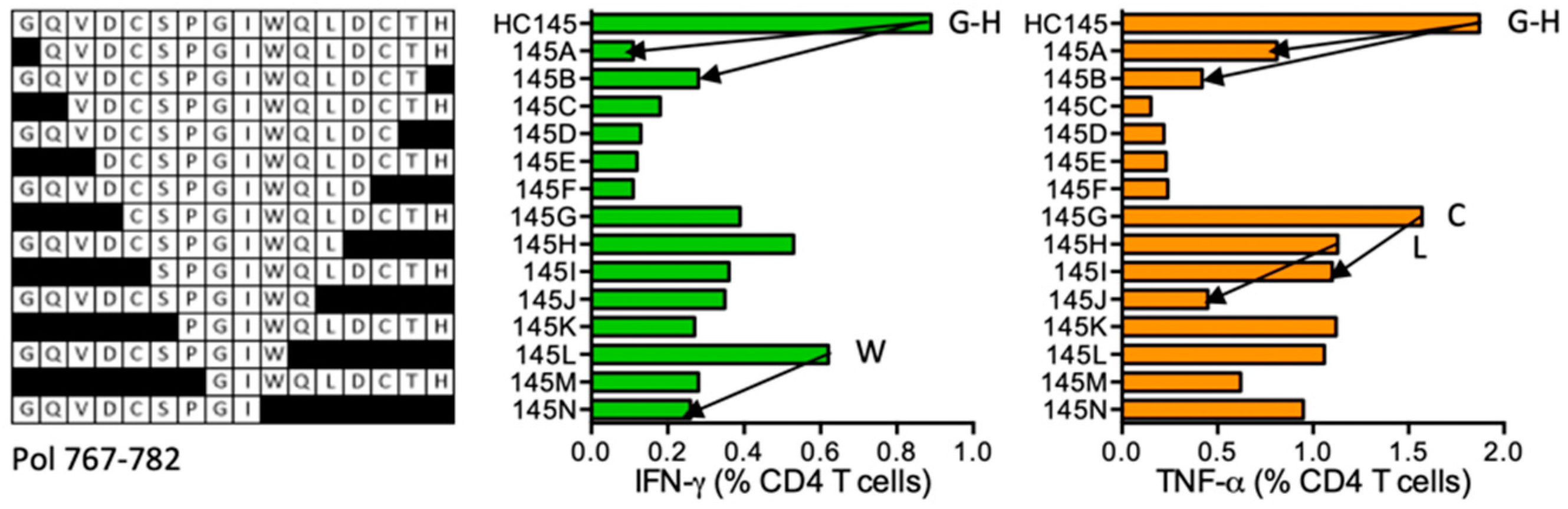
HC145 GQVDCSPGIWQLDCTH (Pol 767–782)
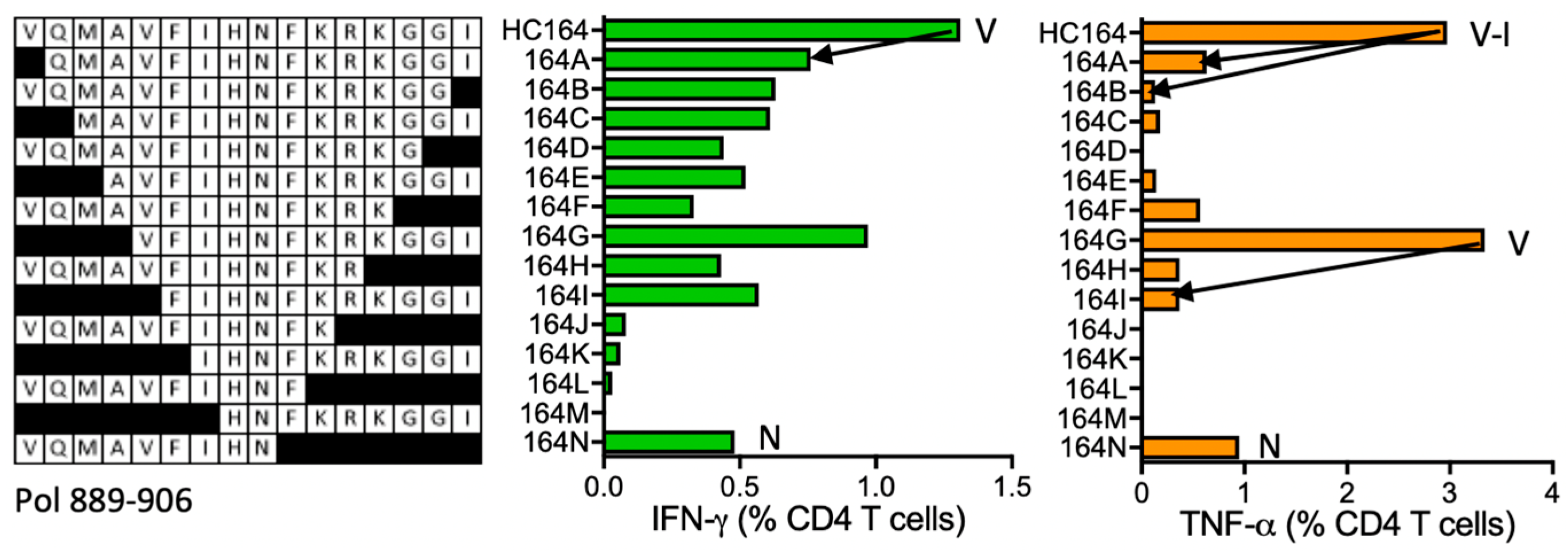
HC164 VQMAVFIHNFKRKGGI (Pol 891–906)
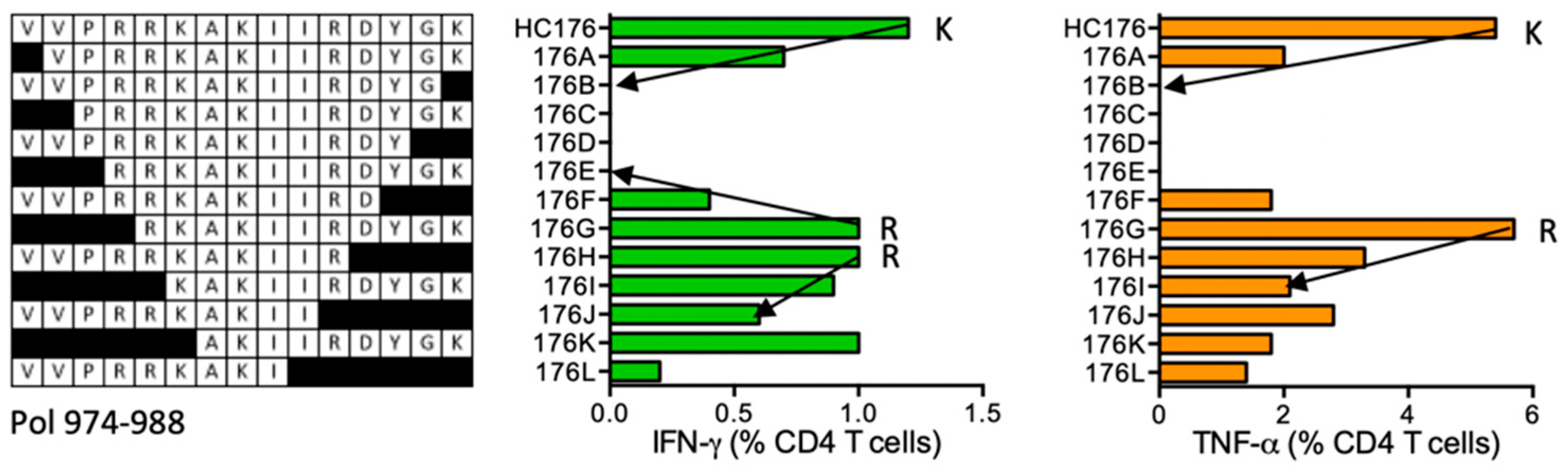
HC176 VVPRRKAKIIRDYGK (Pol 974–988)
HC177 RKAKIIRDYGKQMAG (Pol 978–992)
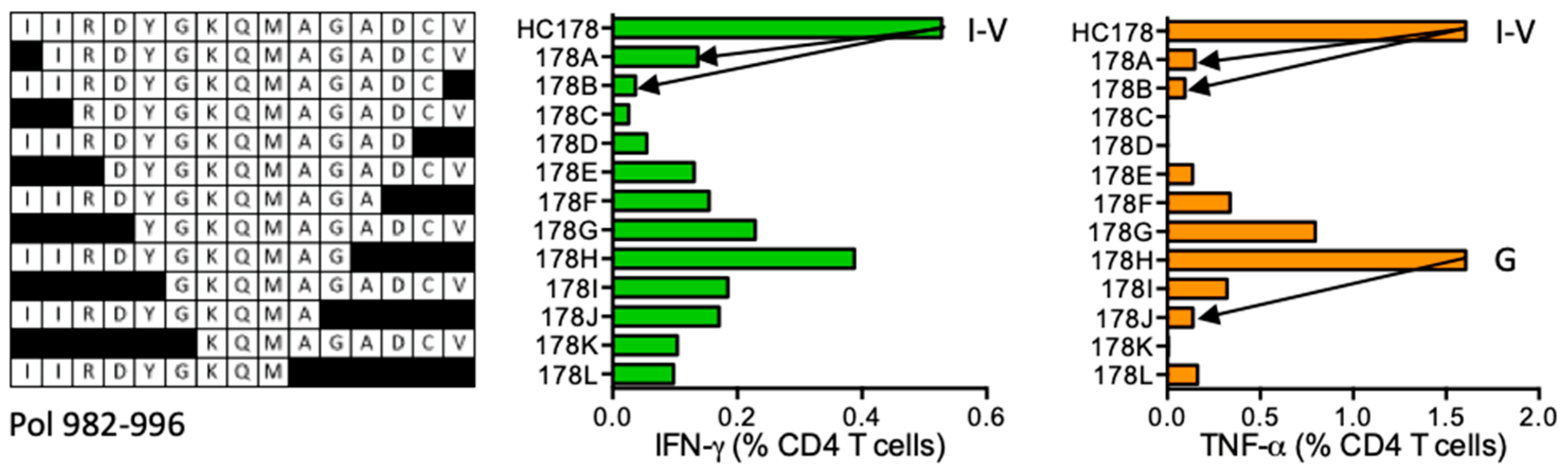
HC178 IIRDYGKQMAGADCV (Pol 982–996)
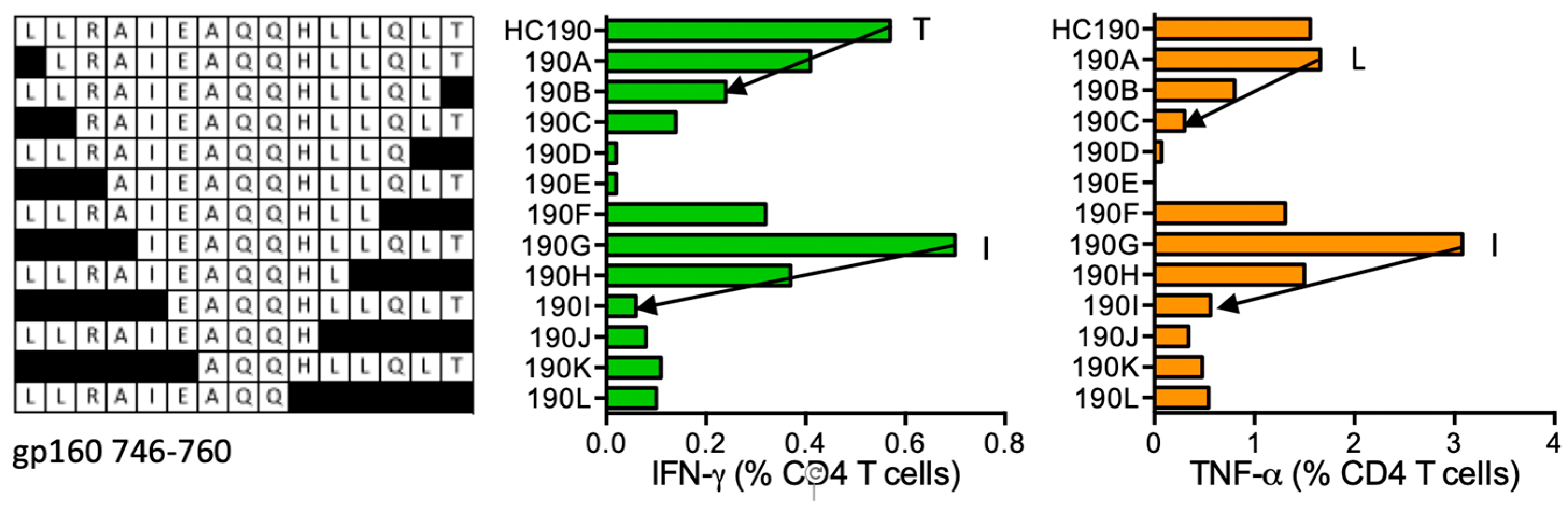
HC190 LLRAIEAQQHLLQLT (gp160 746–760)
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Hanke, T. Aiming for protective T-cell responses: A focus on the first generation conserved-region HIVconsv vaccines in preventive and therapeutic clinical trials. Expert Rev. Vaccines 2019, 18, 1029–1041. [Google Scholar] [CrossRef] [PubMed]
- Walker, B.; McMichael, A. The T-Cell Response to HIV. Cold Spring Harb. Perspect. Med. 2012, 2, a007054. [Google Scholar] [CrossRef] [PubMed]
- Buggert, M.; Nguyen, S.; McLane, L.M.; Steblyanko, M.; Anikeeva, N.; Paquin-Proulx, D.; Del Rio Estrada, P.M.; Ablanedo-Terrazas, Y.; Noyan, K.; Reuter, M.A.; et al. Limited immune surveillance in lymphoid tissue by cytolytic CD4+ T cells during health and HIV disease. PLoS Pathog. 2018, 14, e1006973. [Google Scholar] [CrossRef] [PubMed]
- Malyshkina, A.; Littwitz-Salomon, E.; Sutter, K.; Zelinskyy, G.; Windmann, S.; Schimmer, S.; Paschen, A.; Streeck, H.; Hasenkrug, K.J.; Dittmer, U. Fas Ligand-mediated cytotoxicity of CD4+ T cells during chronic retrovirus infection. Sci. Rep. 2017, 7, 7785. [Google Scholar] [CrossRef]
- Norris, P.J.; Moffett, H.F.; Yang, O.O.; Kaufmann, D.E.; Clark, M.J.; Addo, M.M.; Rosenberg, E.S. Beyond Help: Direct Effector Functions of Human Immunodeficiency Virus Type 1-Specific CD4+ T Cells. J. Virol. 2004, 78, 8844–8851. [Google Scholar] [CrossRef]
- Norris, P.J.; Sumaroka, M.; Brander, C.; Moffett, H.F.; Boswell, S.L.; Nguyen, T.; Sykulev, Y.; Walker, B.D.; Rosenberg, E.S. Multiple effector functions mediated by human immunodeficiency virus-specific CD4(+) T-cell clones. J. Virol. 2001, 75, 9771–9779. [Google Scholar] [CrossRef]
- Pissani, F.; Schulte, B.; Eller, M.A.; Schultz, B.T.; Ratto-Kim, S.; Marovich, M.; Thongcharoen, P.; Sriplienchan, S.; Rerks-Ngarm, S.; Pitisuttithum, P.; et al. Modulation of Vaccine-Induced CD4 T Cell Functional Profiles by Changes in Components of HIV Vaccine Regimens in Humans. J. Virol. 2018, 92, e01143-18. [Google Scholar] [CrossRef]
- Mothe, B.; Llano, A.; Ibarrondo, J.; Daniels, M.; Miranda, C.; Zamarreño, J.; Bach, V.; Zuniga, R.; Pérez-Álvarez, S.; Berger, C.T.; et al. Definition of the viral targets of protective HIV-1-specific T cell responses. J. Transl. Med. 2011, 9, 208. [Google Scholar] [CrossRef]
- Murakoshi, H.; Zou, C.; Kuse, N.; Akahoshi, T.; Chikata, T.; Gatanaga, H.; Oka, S.; Hanke, T.; Takiguchi, M. CD8+ T cells specific for conserved, cross-reactive Gag epitopes with strong ability to suppress HIV-1 replication. Retrovirology 2018, 15, 46. [Google Scholar] [CrossRef]
- Ondondo, B.; Murakoshi, H.; Clutton, G.; Abdul-Jawad, S.; Wee, E.G.-T.; Gatanaga, H.; Oka, S.; McMichael, A.J.; Takiguchi, M.; Korber, B.; et al. Novel Conserved-region T-cell Mosaic Vaccine with High Global HIV-1 Coverage Is Recognized by Protective Responses in Untreated Infection. Mol. Ther. 2016, 24, 832–842. [Google Scholar] [CrossRef]
- Zou, C.; Murakoshi, H.; Kuse, N.; Akahoshi, T.; Chikata, T.; Gatanaga, H.; Oka, S.; Hanke, T.; Takiguchi, M. Effective Suppression of HIV-1 Replication by Cytotoxic T Lymphocytes Specific for Pol Epitopes in Conserved Mosaic Vaccine Immunogens. J. Virol. 2019, 93, e02142-18. [Google Scholar] [CrossRef] [PubMed]
- Ferre, A.L.; Hunt, P.W.; McConnell, D.H.; Morris, M.M.; Garcia, J.C.; Pollard, R.B.; Yee, H.F., Jr.; Martin, J.N.; Deeks, S.G.; Shacklett, B.L. HIV controllers with HLA-DRB1*13 and HLA-DQB1*06 alleles have strong, polyfunctional mucosal CD4+ T-cell responses. J. Virol. 2010, 84, 11020–11029. [Google Scholar] [CrossRef] [PubMed]
- Oriol-Tordera, B.; Llano, A.; Ganoza, C.; Cate, S.; Hildebrand, W.; Sanchez, J.; Calle, M.L.; Brander, C.; Olvera, A. Impact of HLA-DRB1 allele polymorphisms on control of HIV infection in a Peruvian MSM cohort. HLA 2017, 90, 234–237. [Google Scholar] [CrossRef] [PubMed]
- Ranasinghe, S.; Cutler, S.; Davis, I.; Lu, R.; Soghoian, D.Z.; Qi, Y.; Sidney, J.; Kranias, G.; Flanders, M.D.; Lindqvist, M.; et al. Association of HLA-DRB1-restricted CD4(+) T cell responses with HIV immune control. Nat. Med. 2013, 19, 930–933. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, E.S.; Billingsley, J.M.; Caliendo, A.M.; Boswell, S.L.; Sax, P.E.; Kalams, S.A.; Walker, B.D. Vigorous HIV-1-specific CD4+ T cell responses associated with control of viremia. Science 1997, 278, 1447–1450. [Google Scholar] [CrossRef]
- Soghoian, D.Z.; Jessen, H.; Flanders, M.; Sierra-Davidson, K.; Cutler, S.; Pertel, T.; Ranasinghe, S.; Lindqvist, M.; Davis, I.; Lane, K.; et al. HIV-specific cytolytic CD4 T cell responses during acute HIV infection predict disease outcome. Sci. Transl. Med. 2012, 4, 123ra25. [Google Scholar] [CrossRef]
- Llano, A.; Cedeño, S.; Silva-Arrieta, S.; Brander, C. Optimal HIV CTL epitopes update: Growing diversity in epitope length and HLA restriction. In HIV Immunology and HIV/SIV Vaccine Databases; Los Alamos National Laboratory, Theoretical Biology and Biophysics: Los Alamos, NM, USA, 2019. [Google Scholar]
- Llano, A.; Williams, A.; Olvera, A.; Silva-Arrieta, S.; Brander, C. Best-Characterized HIV-1 CTL Epitopes: The 2013 Update. In HIV Immunology and HIV/SIV Vaccine Databases; Los Alamos National Laboratory, Theoretical Biology and Biophysics: Los Alamos, NM, USA, 2013. [Google Scholar]
- Cosimi, L.; Rosenberg, E.S. The characterization of HIV-1 specific CD4+ T helper epitopes. In HIV Molecular Immunology; Los Alamos National Laboratory, Theoretical Biology and Biophysics: Los Alamos, NM, USA, 2000. [Google Scholar]
- Korber, T.M.B.; Brander, C.; Haynes, F.B.; Koup, R.; Kuiken, C.; Moore, J.P.; Walker, B.D.; Watkins, D.I. HIV Molecular Immunology Compendium; Los Alamos National Laboratory: Los Alamos, NM, USA, 2019. [Google Scholar]
- Wilson, C.C.; Palmer, B.; Southwood, S.; Sidney, J.; Higashimoto, Y.; Appella, E.; Chesnut, R.; Sette, A.; Livingston, B.D. Identification and Antigenicity of Broadly Cross-Reactive and Conserved Human Immunodeficiency Virus Type 1-Derived Helper T-Lymphocyte Epitopes. J. Virol. 2001, 75, 4195–4207. [Google Scholar] [CrossRef]
- Khalil-Daher, I.; Boisgérault, F.; Feugeas, J.P.; Tieng, V.; Toubert, A.; Charron, D. Naturally processed peptides from HLA-DQ7 (alpha1*0501-beta1*0301): Influence of both alpha and beta chain polymorphism in the HLA-DQ peptide binding specificity. Eur. J. Immunol. 1998, 28, 3840–3849. [Google Scholar] [CrossRef]
- Brown, J.H.; Jardetzky, T.S.; Gorga, J.C.; Stern, L.J.; Urban, R.G.; Strominger, J.L.; Wiley, N.C. Three-dimensional structure of the human class II histocompatibility antigen HLA-DR1. Nature 1993, 364, 33–39. [Google Scholar] [CrossRef]
- Dessen, A.; Lawrence, C.; Cupo, S.; Zaller, D.M.; Wiley, N.C. X-ray crystal structure of HLA-DR4 (DRA*0101, DRB1*0401) complexed with a peptide from human collagen II. Immunity 1997, 7, 473–481. [Google Scholar] [CrossRef]
- Fremont, D.H.; Hendrickson, W.A.; Marrack, P.; Kappler, J. Structures of an MHC Class II Molecule with Covalently Bound Single Peptides. Science 1996, 272, 1001–1004. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.J.; Pyrdol, J.; Gauthier, L.; Wiley, D.C.; Wucherpfennig, K.W. Crystal Structure of HLA-DR2 (DRA*0101, DRB1*1501) Complexed with a Peptide from Human Myelin Basic Protein. J. Exp. Med. 1998, 188, 1511–1520. [Google Scholar] [CrossRef] [PubMed]
- Stern, L.J.; Brown, J.H.; Jardetzky, T.S.; Gorga, J.C.; Urban, R.G.; Strominger, J.L.; Wiley, N.C. Crystal structure of the human class II MHC protein HLA-DR1 complexed with an influenza virus peptide. Nature 1994, 368, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Chicz, R.M.; Urban, R.G.; Lane, W.S.; Gorga, J.C.; Stern, L.J.; Vignali, D.A.A.; Strominger, J.L. Predominant naturally processed peptides bound to HLA-DR1 are derived from MHC-related molecules and are heterogeneous in size. Nature 1992, 358, 764–768. [Google Scholar] [CrossRef]
- Rudensky, A.Y.; Preston-Hurlburt, P.; Hong, S.-C.; Barlow, A.; Janeway, C.A. Sequence analysis of peptides bound to MHC class II molecules. Nature 1991, 353, 622–627. [Google Scholar] [CrossRef]
- Vignali, D.A.A.; Urban, R.G.; Chicz, R.M.; Strominger, J.L. Minute quantities of a single immunodominant foreign epitope are presented as large nested sets by major histocompatibility complex class II molecules. Eur. J. Immunol. 1993, 23, 1602–1607. [Google Scholar] [CrossRef]
- Kaufmann, D.E.; Bailey, P.M.; Sidney, J.; Wagner, B.; Norris, P.J.; Johnston, M.N.; Cosimi, L.A.; Addo, M.M.; Lichterfeld, M.; Altfeld, M.; et al. Comprehensive Analysis of Human Immunodeficiency Virus Type 1-Specific CD4 Responses Reveals Marked Immunodominance of gag and nef and the Presence of Broadly Recognized Peptides. J. Virol. 2004, 78, 4463–4477. [Google Scholar] [CrossRef]
- Sette, A.; Sidney, J. Nine major HLA class I supertypes account for the vast preponderance of HLA-A and -B polymorphism. Immunogenetics 1999, 50, 201–212. [Google Scholar] [CrossRef]
- Daniels, M.A.; Schober, S.L.; Hogquist, K.A.; Jameson, S.C. Cutting edge: A test of the dominant negative signal model for TCR antagonism. J. Immunol. 1999, 162, 3761–3764. [Google Scholar]
- Jacobs, E.S.; Persad, D.; Ran, L.; Danesh, A.; Heitman, J.W.; Deng, X.; Cameron, M.J.; Kelvin, D.J.; Norris, P.J. A CD4+ T cell antagonist epitope down-regulates activating signaling proteins, up-regulates inhibitory signaling proteins and abrogates HIV-specific T cell function. Retrovirology 2014, 11, 57. [Google Scholar] [CrossRef]
- Norris, P.J.; Stone, J.D.; Anikeeva, N.; Heitman, J.W.; Wilson, I.C.; Hirschkorn, D.F.; Clark, M.J.; Moffett, H.F.; Cameron, T.O.; Sykulev, Y.; et al. Antagonism of HIV-specific CD4+ T cells by C-terminal truncation of a minimum epitope. Mol. Immunol. 2006, 43, 1349–1357. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Robertson, J.M.; Evavold, B.D. Cutting edge: Dueling TCRs: Peptide antagonism of CD4+ T cells with dual antigen specificities. J. Immunol. 1999, 163, 1750–1754. [Google Scholar] [PubMed]
- Stotz, S.H.; Bolliger, L.; Carbone, F.R.; Palmer, E. T Cell Receptor (TCR) Antagonism without a Negative Signal: Evidence from T Cell Hybridomas Expressing Two Independent TCRs. J. Exp. Med. 1999, 189, 253–264. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Grey, H.M. Study of the mechanism of TCR antagonism using dual-TCR-expressing T cells. J. Immunol. 2003, 170, 4532–4538. [Google Scholar] [CrossRef] [PubMed]
- Los Alamos National Laboratory HIV Sequence Database. 2019. Available online: www.hiv.lanl.gov (accessed on 2 December 2019).
- Létourneau, S.; Im, E.-J.; Mashishi, T.; Brereton, C.; Bridgeman, A.; Yang, H.; Dorrell, L.; Dong, T.; Korber, B.; McMichael, A.J.; et al. Design and Pre-Clinical Evaluation of a Universal HIV-1 Vaccine. PLoS ONE 2007, 2, e984. [Google Scholar] [CrossRef]
- Ahmed, T.; Borthwick, N.J.; Gilmour, J.; Hayes, P.; Dorrell, L.; Hanke, T. Control of HIV-1 replication in vitro by vaccine-induced human CD8(+) T cells through conserved subdominant Pol epitopes. Vaccine 2016, 34, 1215–1224. [Google Scholar] [CrossRef][Green Version]
- Borthwick, N.; Lin, Z.; Akahoshi, T.; Llano, A.; Silva-Arrieta, S.; Ahmed, T.; Dorrell, L.; Brander, C.; Murakoshi, H.; Takiguchi, M.; et al. Novel, in-natural-infection subdominant HIV-1 CD8+ T-cell epitopes revealed in human recipients of conserved-region T-cell vaccines. PLoS ONE 2017, 12, e0176418. [Google Scholar] [CrossRef]
- Frahm, N.; Kiepiela, P.; Adams, S.; Linde, C.H.; Hewitt, H.S.; Sango, K.; Feeney, M.E.; Addo, M.M.; Lichterfeld, M.; Lahaie, M.P.; et al. Control of human immunodeficiency virus replication by cytotoxic T lymphocytes targeting subdominant epitopes. Nat. Immunol. 2006, 7, 173–178. [Google Scholar] [CrossRef]
- Borthwick, N.; Ahmed, T.; Ondondo, B.; Hayes, P.; Rose, A.; Ebrahimsa, U.; Hayton, E.-J.; Black, A.; Bridgeman, A.; Rosario, M.; et al. Vaccine-elicited Human T Cells Recognizing Conserved Protein Regions Inhibit HIV-1. Mol. Ther. 2014, 22, 464–475. [Google Scholar] [CrossRef]
- Hayton, E.-J.; Rose, A.; Ibrahimsa, U.; Del Sorbo, M.; Capone, S.; Crook, A.; Black, A.P.; Dorrell, L.; Hanke, T. Safety and Tolerability of Conserved Region Vaccines Vectored by Plasmid DNA, Simian Adenovirus and Modified Vaccinia Virus Ankara Administered to Human Immunodeficiency Virus Type 1-Uninfected Adults in a Randomized, Single-Blind Phase I Trial. PLoS ONE 2014, 9, e101591. [Google Scholar] [CrossRef]
- ELF Epitope Location Finder. 2018. Available online: www.hiv.lanl.gov/content/sequence/ELF/epitope_analyzer.html (accessed on 2 December 2019).
- Helper Epitopes. 2019. Available online: www.hiv.lanl.gov/content/immunology/tables/helper_summary.html (accessed on 2 December 2019).
- Allan, E.R.O.; Tailor, P.; Balce, D.R.; Pirzadeh, P.; McKenna, N.T.; Renaux, B.; Warren, A.L.; Jirik, F.R.; Yates, R.M. NADPH Oxidase Modifies Patterns of MHC Class II–Restricted Epitopic Repertoires through Redox Control of Antigen Processing. J. Immunol. 2014, 192, 4989–5001. [Google Scholar] [CrossRef] [PubMed]
- Cardinaud, S.; Moris, A.; Février, M.; Rohrlich, P.-S.; Weiss, L.; Langlade-Demoyen, P.; Lemonnier, F.A.; Schwartz, O.; Habel, A. Identification of Cryptic MHC I–restricted Epitopes Encoded by HIV-1 Alternative Reading Frames. J. Exp. Med. 2004, 199, 1053–1063. [Google Scholar] [CrossRef] [PubMed]
- Frey, B.F.; Jiang, J.; Sui, Y.; Boyd, L.F.; Yu, B.; Tatsuno, G.; Billeskov, R.; Solaymani-Mohammadi, S.; Berman, P.W.; Margulies, D.H.; et al. Effects of Cross-Presentation, Antigen Processing, and Peptide Binding in HIV Evasion of T Cell Immunity. J. Immunol. 2018, 200, 1853–1864. [Google Scholar] [CrossRef] [PubMed]
- Norris, P.J.; Moffett, H.F.; Brander, C.; Allen, T.M.; O’Sullivan, K.M.; Cosimi, L.A.; Kaufmann, D.E.; Walker, B.D.; Rosenberg, E.S. Fine specificity and cross-clade reactivity of HIV type 1 Gag-specific CD4+ T cells. AIDS Res. Hum. Retrovir. 2004, 20, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Sturniolo, T.; Bono, E.; Ding, J.; Raddrizzani, L.; Tuereci, O.; Sahin, U.; Braxenthaler, M.; Gallazzi, F.; Protti, M.P.; Sinigaglia, F.; et al. Generation of tissue-specific and promiscuous HLA ligand databases using DNA microarrays and virtual HLA class II matrices. Nat. Biotechnol. 1999, 17, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Appay, V.; Dunbar, P.R.; Callan, M.; Klenerman, P.; Gillespie, G.M.; Papagno, L.; Ogg, G.S.; King, A.; Lechner, F.; Spina, C.A.; et al. Memory CD8+ T cells vary in differentiation phenotype in different persistent virus infections. Nat. Med. 2002, 8, 379–385. [Google Scholar] [CrossRef]
- Hanke, T. Conserved immunogens in prime-boost strategies for the next-generation HIV-1 vaccines. Expert Opin. Biol. Ther. 2014, 14, 601–616. [Google Scholar] [CrossRef]
- Betts, M.R.; Ambrozak, D.R.; Douek, D.C.; Bonhoeffer, S.; Brenchley, J.M.; Casazza, J.P.; Koup, R.A.; Picker, L.J. Analysis of total human immunodeficiency virus (HIV)-specific CD4(+) and CD8(+) T-cell responses: Relationship to viral load in untreated HIV infection. J. Virol. 2001, 75, 11983–11991. [Google Scholar] [CrossRef]
- Havenar-Daughton, C.; Reiss, S.M.; Carnathan, D.G.; Wu, J.E.; Kendric, K.; De La Peña, A.T.; Kasturi, S.P.; Dan, J.M.; Bothwell, M.; Sanders, R.W.; et al. Cytokine-independent detection of antigen-specific germinal center T follicular helper (Tfh) cells in immunized non-human primates using a live cell Activation Induced Marker (AIM) technique1. J. Immunol. 2016, 197, 994–1002. [Google Scholar] [CrossRef]
- Irvine, D.J.; Purbhoo, M.A.; Krogsgaard, M.; Davis, M.M. Direct observation of ligand recognition by T cells. Nature 2002, 419, 845–849. [Google Scholar] [CrossRef]
- Janeway, C.A. The T Cell Receptor as a Multicomponent Signalling Machine: CD4/CD8 Coreceptors and CD45 in T Cell Activation. Annu. Rev. Immunol. 1992, 10, 645–674. [Google Scholar] [CrossRef] [PubMed]
- Mothe, B.; Manzardo, C.; Snachez-Bernabeau, A.; Coll, P.; Moron-Lopez, S.; Puertas, M.C.; Rosas, M.; Cobarsi, P.; Escrig, N.; Perez-Alvarez, N.; et al. Therapeutic vaccination refocused T-cell responses to conserved regions of HIV-1 in early reated individuals (BCN 01 study). EClinicalMedicine 2019, 1, 65–80. [Google Scholar] [CrossRef] [PubMed]
- Mothe, B.; Rosas-Umbert, M.; Coll, P.; Manzardo, C.; Puertas, M.C.; Morón-López, S.; Llano, A.; Miranda, C.; Cedeño, S.; López, M.; et al. HIVconsv vaccines and romidepsin in early-treated HIV-1-infected individuals: Safety, immunogenicity and effect on the viral reservoir (study BCN 02). Sci. Transl. Med. submitted.
- Bliss, C.M.; Bowyer, G.; Anagnostou, N.A.; Havelock, T.; Snudden, C.M.; Davies, H.; De Cassan, S.C.; Grobbelaar, A.; Lawrie, A.M.; Venkatraman, N.; et al. Assessment of novel vaccination regimens using viral vectored liver stage malaria vaccines encoding ME-TRAP. Sci. Rep. 2018, 8, 3390. [Google Scholar] [CrossRef]
- Borthwick, N.J.; Lane, T.; Moyo, N.; Crook, A.; Shim, J.M.; Baines, I.; Wee, E.G.; Hawkins, P.H.; Gillmore, J.D.; Hanke, T.K. Randomized phase I trial HIV-CORE 003: Depletion of serum amyloid P component and immunogenicity of DNA vaccination against HIV-1. PLoS ONE 2018, 13, e0197299. [Google Scholar] [CrossRef]
- Moore, J.P.; Burton, D.R. HIV-1 neutralizing antibodies: How full is the bottle? Nat. Med. 1999, 5, 204–216. [Google Scholar] [CrossRef]





















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Volunteer ID | HLA Class II | Regimen 1 | Peak IFN-γ ELISPOT | ||
|---|---|---|---|---|---|
| DRB1 | DRB2/3/4 | DQB1 | (SFU/106 PBMC) 2 | ||
| 403 | *07:01 *16:01 | DRB4 DRB5 | 02 05 | CM | 1475 |
| 404 | *13:01 *13:02 | DRB3 DRB3 | *06:02 *06:05 | CM | 3780 |
| 406 | *01:01 *07:01 | DRB4 DRB4 | 02 05 | CM | 5170 |
| 409 | *03:01 *15:01 | DRB3 DRB5 | 02 *06:02 | CM | 5150 |
| 411 | *03:01 *09:01 | BRB3 DRB5 | 02 *03:03 | CM | 12,020 |
| 413 | *03:01 *11:01 | DRB3 DRB3 | 02 *03:01 | DDDCM | 6240 |
| 414 | *01:02 *04:01 | DRB4 DRB4 | 05 *03:01 | DDDCM | 7960 |
| 415 | *11:04 *15:01 | DRB3 DRB5 | *06:02 *03:01 | DDDCM | 5340 |
| 417 | *01:01 *07:01 | BRB4 DRB4 | 02 05 | DDDCM | 4260 |
| 418 | *04:01 *07:01 | DRB4 DRB4 | *03:01 *03:03 | DDDCM | 7060 |
| 420 | *11:01 *15:01 | DRB3 DRB5 | *03:01 *06:02 | DDDCM | 1530 |
| Peptide No. | Protein (HXB2 Position) | Stimulatory Peptides | IFN-γ/TNF-α | Responder’s HLA-DRB1 | Predicted HLA-DRB1 | Reported Epitopes | Reported Epitopes’ Restriction | Epitope Core | Epitope Core HLA-DRB1 |
|---|---|---|---|---|---|---|---|---|---|
| HC003 | Gag (167–181) | EVIPMFTALSEGATP EVIPMFTALSEGAT EVIPMFTALSEGA EVIPMFTALSEG EVIPMFTALSE EVIPMFTALS | IFN-γ ≈ TNF-α | *04:01 *07:01 | *04:01 *07:01 | PMFTALSEGAT PEVIPMFSALSEGA EVIPMFSALS | Human DR1 *01:01 DR4 | EVIPMFTALS FTALSEGAT VIPMFT LSEGAT | *04:01 *04:01 *07:01 *07:01 |
| HC017 | Gag (269–283) | GLNKIVRMYSPVSIL GLNKIVRMYSPVSI GLNKIVRMYSPVS | IFN-γ > TNF-α | *04:01 *07:01 | *04:01 *07:01 | GLNKIVRMYSPVSIL GLNKIVRMYSPTSIL LNKIVRMYSPVSILD LNKIVRMYSPTSILD KIVRMYSPTS KIVRMYSPT | human *01:01 human human DR4 *01:01 | LNKIVRMYS IVRMYS | *04:01 *07:01 |
| LNKIVRMYSPVSIL NKIVRMYSPVSIL KIVRMYSPVSIL | |||||||||
| HC078 | Pol (270–284) | YFSVPLDEGFRKYTA FSVPLDEGFRKYTA SVPLDEGFRKYTA VPLDEGFRKYTA PLDEGFRKYTA | IFN-γ > TNF-α IFN-γ TNF-α | *01:01 *07:01 | |||||
| HC079 | Pol (274–288) | PLDEGFRKYTAFTIP FRKYTAFTIP | TNF-α IFN-γ | *01:01 *07:01 | |||||
| HC080 | Pol (278–292) | GFRKYTAFTIPSINN GFRKYTAFTIPSIN GFRKYTAFTIPSI GFRKYTAFTIPS | IFN-γ ≈ TNF-α | *03:01 *09:01 | *07:01 | FRKYTAFTIPSINNE | DR supermotif | ||
| FRKYTAFTIPSINN RKYTAFTIPSINN KYTAFTIPSINN YTAFTIPSINN | |||||||||
| HC081 | Pol (282–296) | YTAFTIPSINNETPG TIPSINNETPG | TNF-α IFN-γ < TNF-α | *11:04 *15:01 | |||||
| HC088 | Pol (311–325) | GSPAIFQSSMTKILE. GSPAIFQSSMTKIL | IFN-γ ≈ TNF-α | *03:01 *09:01 | *04:01 | GSPAIFQSSMTKILE SPAIFQSSMTKILEP | human DR supermotif | ||
| SPAIFQSSMTKILE PAIFQSSMTKILE AIFQSSMTKILE IFQSSMTKILE FQSSMTKILE | |||||||||
| HC093 | Pol (330–344) | KNPEIVIYQYMDDLYV KNPEIVIYQYMD NPEIVIYQYMDDLYV IVIYQYMDDLYV | TNF-α TNF-α | *11:04 *15:01 *01:01 *07:01 | IVIYQYM | *15:01 | |||
| HC094 | Pol (334–348) | VIYQYMDDLYVGSDL VIYQYMDDLYVGS VIYQYMDDL | IFN-γ < TNF-α | *11:01 *15:01 | *11:01 | ||||
| YQYMDDLYVGSDL YMDDLYVGSDL DDLYVGSDL | |||||||||
| HC102 | Vif (12–26) | ----RMRIRTWKSLVK | IFN-γ < TNF-α | *07:01 *16:01 | *07:01 | IRTWKS | *07:01 | ||
| HC103 | Vif (16–28) | MRIRTWKSLVKHH-LT MRIRTWKSLVKHH MRIRTWKSLVKH RIRTWKSLVKHH-LT | IFN-γ < TNF-α | *07:01 *16:01 | *07:01 | ||||
| HC103 | Vif (16–28) | MRIRTWKSLVKHH-LT MRIRTWKSLVK | IFN-γ < TNF-α | *03:01 *15:01 | *03:01 *15:01 | IRTWKSLV | *03:01 *15:01 | ||
| HC115 | Pol (560–573) | KQ-YWQATWIPEWEFVN QATWIPEWEFVN | IFN-γ < TNF-α | *01:02 *04:01 | |||||
| HC123 | gp160 (91–106) | KNFNMWKNDMVDQMHE FNMWKNDMVDQMHE NMWKNDMVDQMHE MWKNDMVDQMHE NDMVDQMHE | IFN-γ < TNF-α TNF-α | *11:01 *15:01 | NVTENFNMWKNNMVEQMH | *11:01 *15:01 *15:02 | |||
| HC145 | Pol (767–782) | GQVDCSPGIWQLDCTH GQVDCSPGIWQL GQVDCSPGIW CSPGIWQLDCTH SPGIWQLDCTH | IFN-γ ≈ TNF-α | *13:01 *13:02 | |||||
| HC164 | Pol (891–906) | VQMAVFIHNFKRKGGI VQMAVFIHN VFIHNFKRKGGI | IFN-γ < TNF-α | *13:01 *13:02 | *13:01 | KTAVQMAVFIHNFKR | DR supermotif | ||
| HC176 | Pol (974–988) | VVPRRKAKIIRDYGK VVPRRKAKIIR VVPRRKAKII | IFN-γ < TNF-α | *03:01 *11:01 | *11:01 | SDIKVVPRRKAKIIR | human | ||
| VPRRKAKIIRDYGK RKAKIIRDYGK KAKIIRDYGK AKIIRDYGK | |||||||||
| HC177 | Pol (978–992) | RKAKIIRDYGKQMAG RKAKIIRDYGKQMA RKAKIIRDYGKQM RKAKIIRDYG | IFN-γ ≈ TNF-α | *03:01 *15:01 | *03:01 | IIRDYGKQM | *03:01 | ||
| KAKIIRDYGKQMAG AKIIRDYGKQMAG KIIRDYGKQMAG IIRDYGKQMAG IRDYGKQMAG RDYGKQMAG | |||||||||
| HC178 | Pol (982–996) | IIRDYGKQMAGADCV IIRDYGKQMAG | IFN-γ < TNF-α | *07:01 *16:01 | |||||
| HC190 | gp160 (745–760) | LLRAIEAQQHLLQLT LLRAIEAQQHLL LLRAIEAQQHL LLRAIEAQQHLLQLT IEAQQHLLQLT | *01:02 *04:01 | *01:02 | LLRAIEAQQ | *04:01 |
| Protein | Total Region Length (AA Residues) | Stimulatory CD4 T-Cell Epitopes (No.) |
|---|---|---|
| Gag | 134 (17%) | 2 (11%) |
| Pol | 526 (68%) | 13 (68%) |
| Vif | 28 (4%) | 2 (11%) |
| Env | 91 (12%) | 2 (11%) |
| HIVconsv | 776 (100%) | 19 (100%) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borthwick, N.; Silva-Arrieta, S.; Llano, A.; Takiguchi, M.; Brander, C.; Hanke, T. Novel Nested Peptide Epitopes Recognized by CD4+ T Cells Induced by HIV-1 Conserved-Region Vaccines. Vaccines 2020, 8, 28. https://doi.org/10.3390/vaccines8010028
Borthwick N, Silva-Arrieta S, Llano A, Takiguchi M, Brander C, Hanke T. Novel Nested Peptide Epitopes Recognized by CD4+ T Cells Induced by HIV-1 Conserved-Region Vaccines. Vaccines. 2020; 8(1):28. https://doi.org/10.3390/vaccines8010028
Chicago/Turabian StyleBorthwick, Nicola, Sandra Silva-Arrieta, Anuska Llano, Masafumi Takiguchi, Christian Brander, and Tomáš Hanke. 2020. "Novel Nested Peptide Epitopes Recognized by CD4+ T Cells Induced by HIV-1 Conserved-Region Vaccines" Vaccines 8, no. 1: 28. https://doi.org/10.3390/vaccines8010028
APA StyleBorthwick, N., Silva-Arrieta, S., Llano, A., Takiguchi, M., Brander, C., & Hanke, T. (2020). Novel Nested Peptide Epitopes Recognized by CD4+ T Cells Induced by HIV-1 Conserved-Region Vaccines. Vaccines, 8(1), 28. https://doi.org/10.3390/vaccines8010028