Rapid and Broad Immune Efficacy of a Recombinant Five-Antigen Vaccine against Staphylococcus aureus Infection in Animal Models

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Bacterial Strains and Culture Methods
2.3. Animals
2.4. Cloning, Expression and Purification of Recombinant Vaccine Antigens
2.5. Mouse Immunization
2.6. ELISA
2.7. S. aureus Sepsis Mouse Model
2.8. S. aureus Pneumonia Mouse Model
2.9. Histological Analysis
2.10. Evaluation of Inflammation
2.11. Immune Response Assays
2.12. Opsonophagocytic Killing Assay
2.13. Hemolytic Activity Assay and Intestinal Toxin Activity Assay
2.14. B Cell Apoptosis Assay
2.15. Statistical Analysis
3. Results
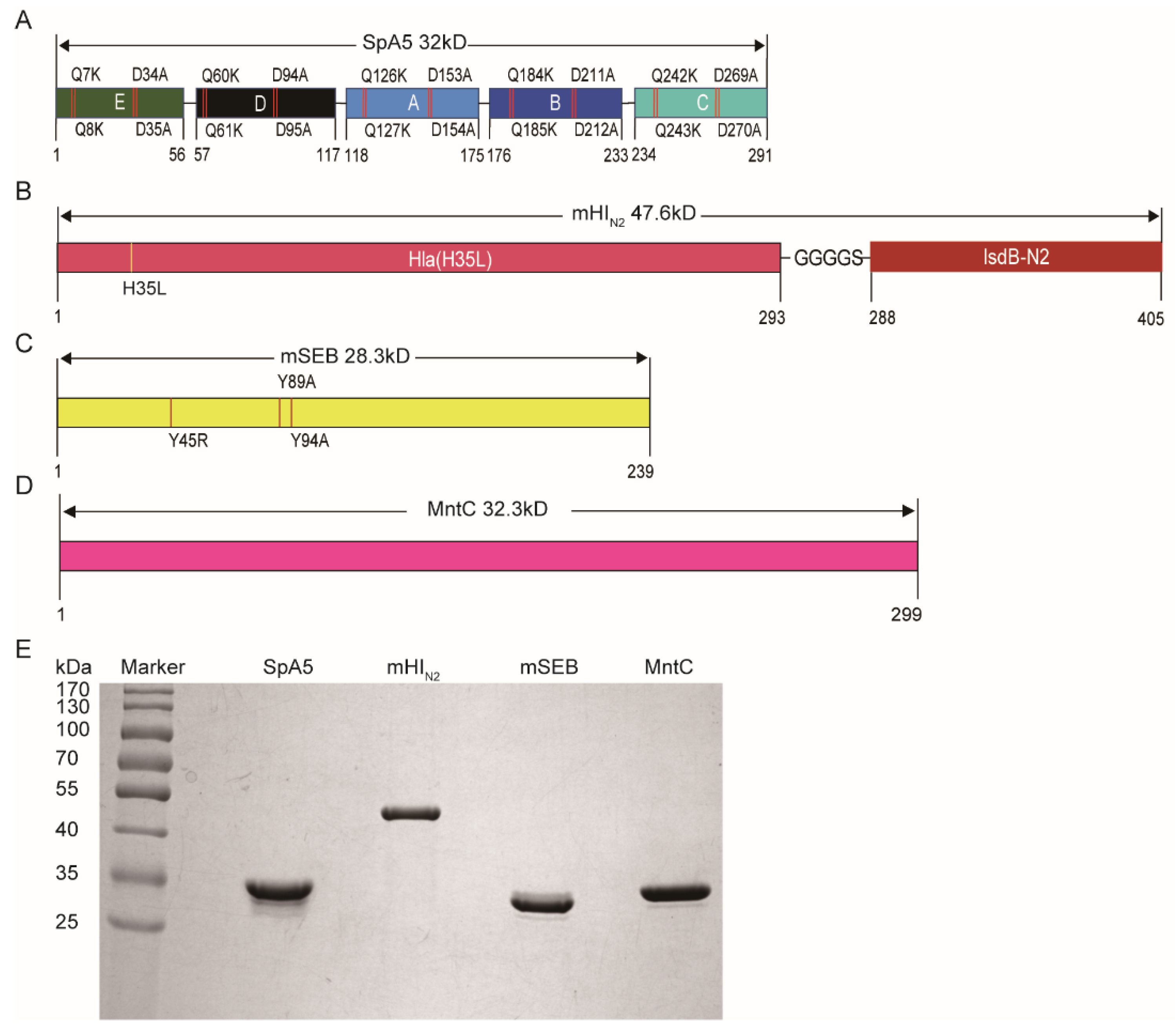
3.1. Antigen Selection and Modification
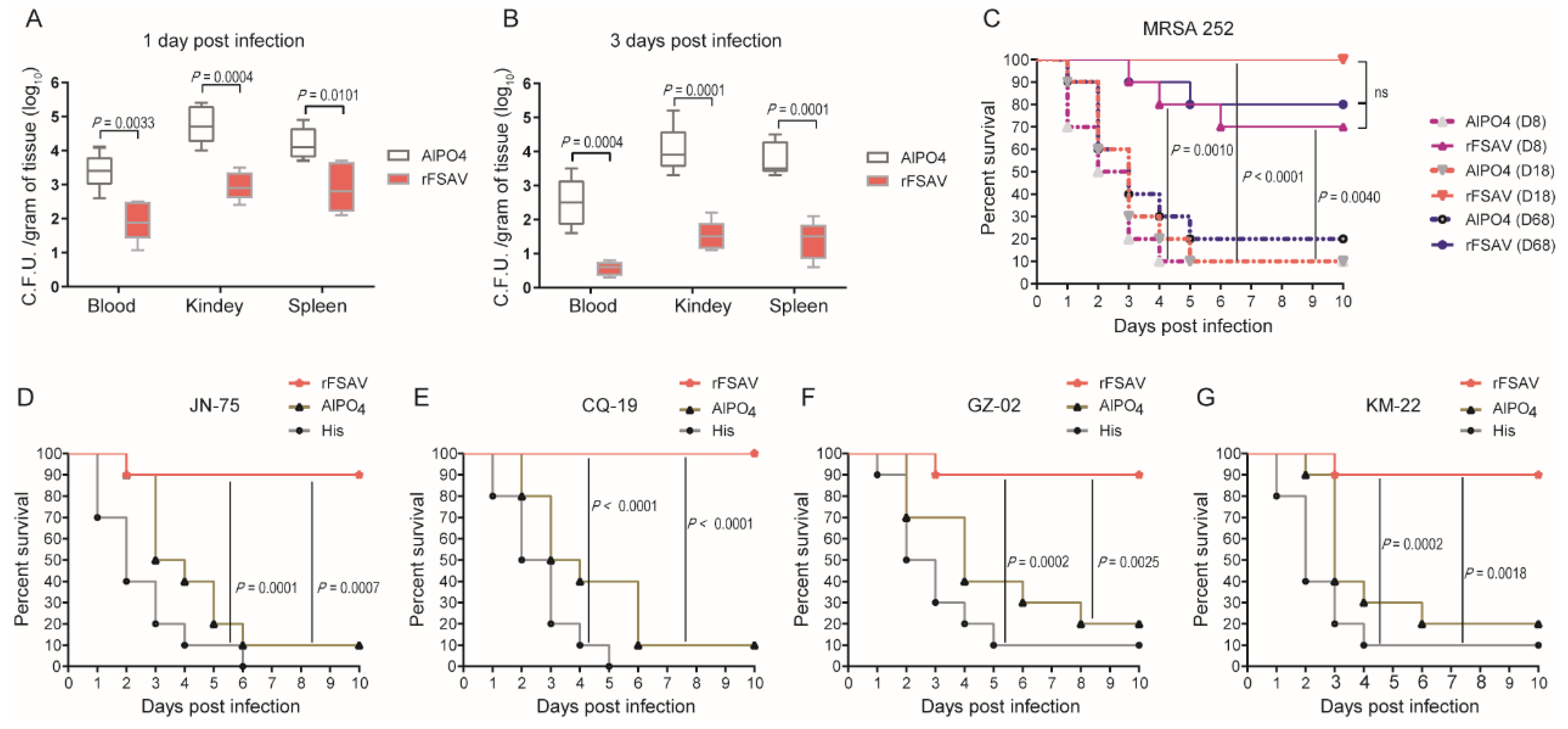
3.2. rFSAV Induces Consistent Immunoprotection against S. aureus in a Sepsis Model
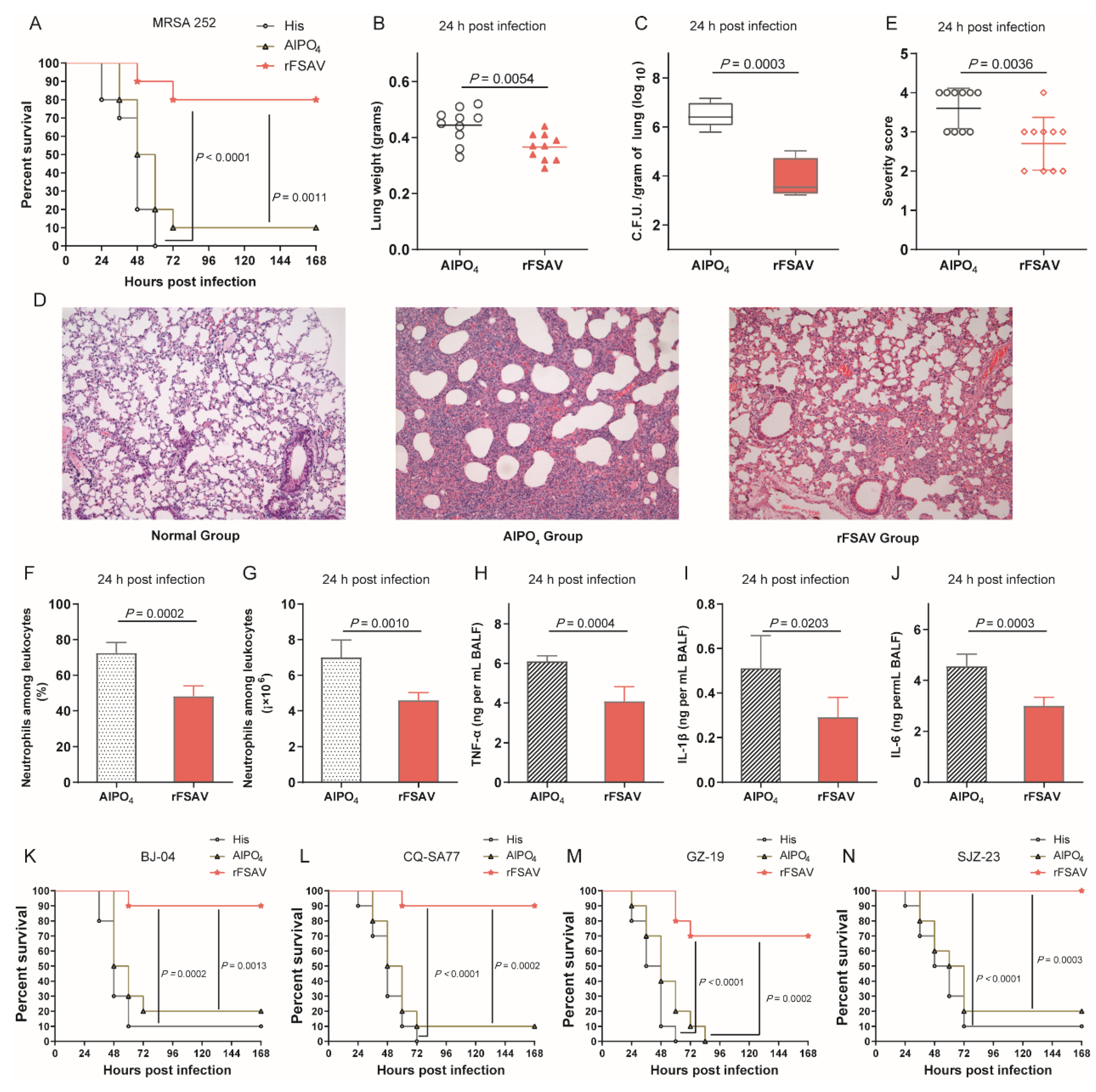
3.3. rFSAV Vaccination Protects Mice from Pneumonia by Reducing Local Bacterial Burden and Inflammation
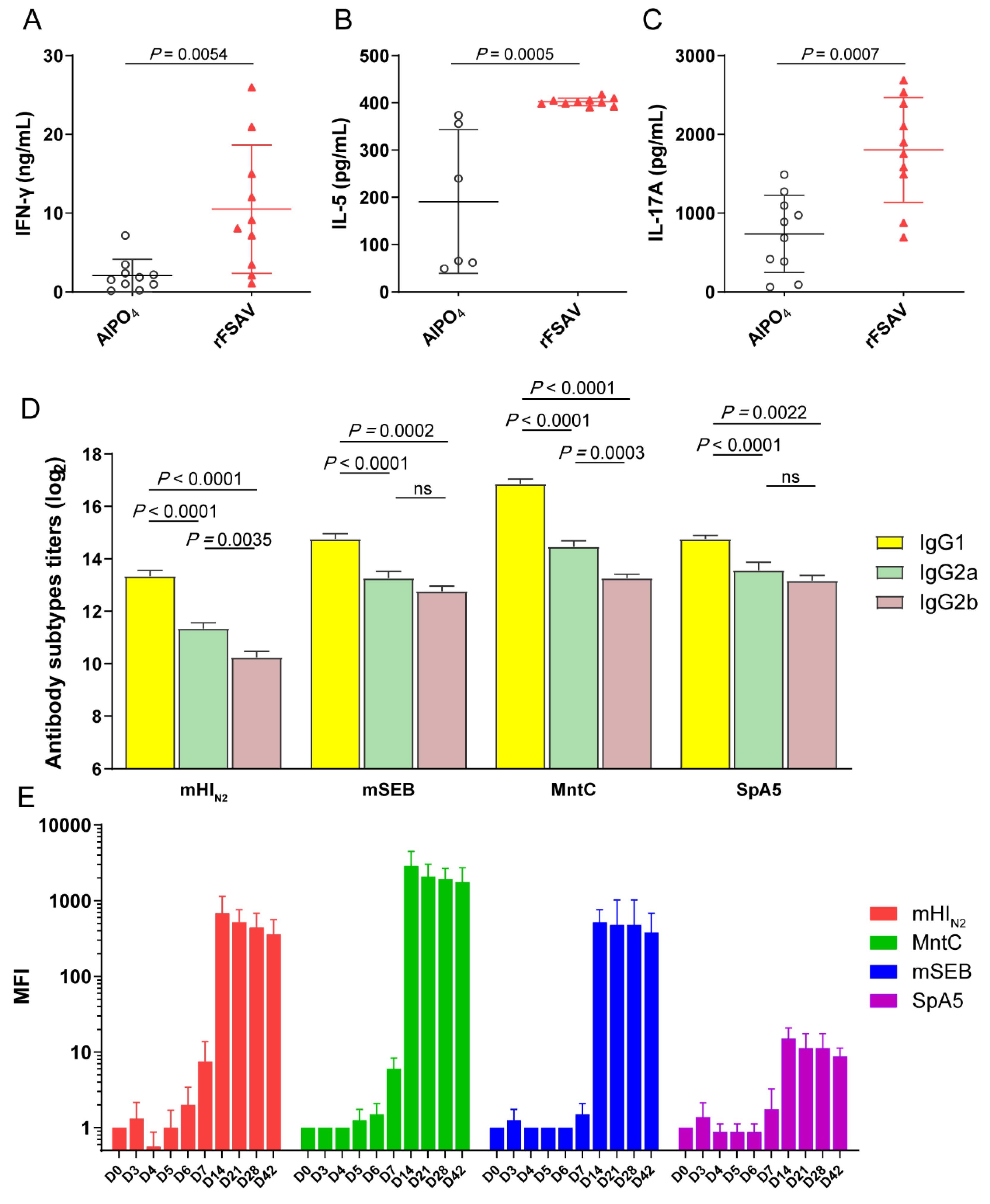
3.4. rFSAV Vaccination Elicits Specific CD4 T-Cell Responses and a Rapid Humoral Immune Response
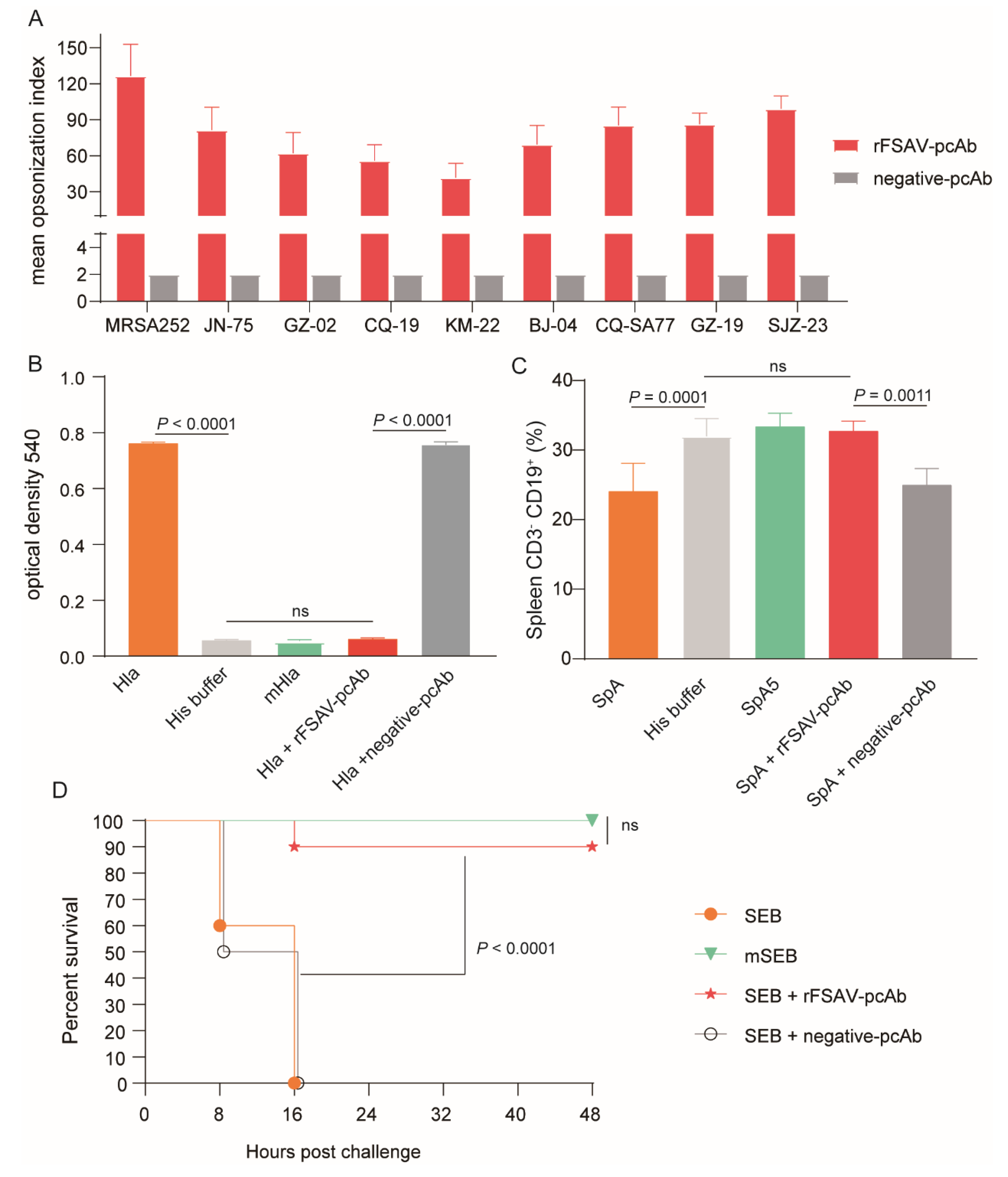
3.5. rFSAV-pcAb Is Effectivity on Promoting Opsonophagocytosis and Neutralizing Toxins
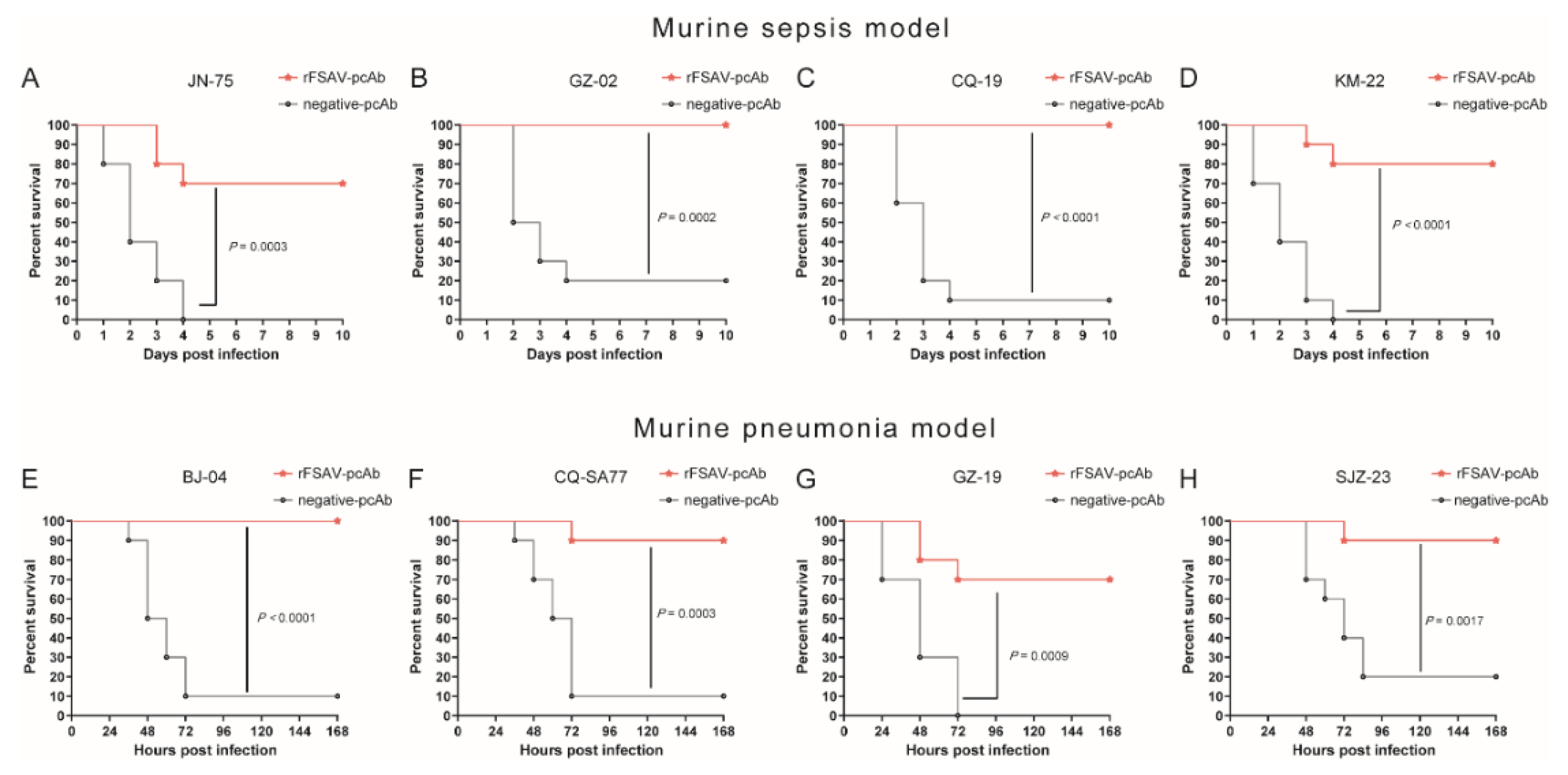
3.6. Passive Immunization Protects Mice against Clinical S. aureus Strains in Murine Sepsis and Pneumonia Models
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Oliveira, D.; Borges, A.; Simoes, M. Staphylococcus aureus Toxins and Their Molecular Activity in Infectious Diseases. Toxins 2018, 10, 252. [Google Scholar] [CrossRef] [PubMed]
- Lakhundi, S.; Zhang, K. Methicillin-Resistant Staphylococcus aureus: Molecular Characterization, Evolution, and Epidemiology. Clin. Microbiol. Rev. 2018, 31, e00018. [Google Scholar] [CrossRef] [PubMed]
- Magill, S.S.; Edwards, J.R.; Bamberg, W.; Beldavs, Z.G.; Dumyati, G.; Kainer, M.A.; Lynfield, R.; Maloney, M.; McAllister-Hollod, L.; Nadle, J.; et al. Multistate point-prevalence survey of health care-associated infections. N. Engl. J. Med. 2014, 370, 1198–1208. [Google Scholar] [CrossRef] [PubMed]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Ansari, S.; Jha, R.K.; Mishra, S.K.; Tiwari, B.R.; Asaad, A.M. Recent advances in Staphylococcus aureus infection: Focus on vaccine development. Infect. Drug Resist. 2019, 12, 1243–1255. [Google Scholar] [CrossRef]
- Pozzi, C.; Olaniyi, R.; Liljeroos, L.; Galgani, I.; Rappuoli, R.; Bagnoli, F. Vaccines for Staphylococcus aureus and Target Populations. Curr. Top. Microbiol. Immunol. 2017, 409, 491–528. [Google Scholar] [CrossRef]
- Redi, D.; Raffaelli, C.S.; Rossetti, B.; De Luca, A.; Montagnani, F. Staphylococcus aureus vaccine preclinical and clinical development: Current state of the art. New Microbiol. 2018, 41, 208–213. [Google Scholar]
- Fowler, V.G.; Allen, K.B.; Moreira, E.D.; Moustafa, M.; Isgro, F.; Boucher, H.W.; Corey, G.R.; Carmeli, Y.; Betts, R.; Hartzel, J.S.; et al. Effect of an investigational vaccine for preventing Staphylococcus aureus infections after cardiothoracic surgery: A randomized trial. JAMA 2013, 309, 1368–1378. [Google Scholar] [CrossRef]
- Harro, C.D.; Betts, R.F.; Hartzel, J.S.; Onorato, M.T.; Lipka, J.; Smugar, S.S.; Kartsonis, N.A. The immunogenicity and safety of different formulations of a novel Staphylococcus aureus vaccine (V710): Results of two Phase I studies. Vaccine 2012, 30, 1729–1736. [Google Scholar] [CrossRef]
- Fattom, A.; Matalon, A.; Buerkert, J.; Taylor, K.; Damaso, S.; Boutriau, D. Efficacy profile of a bivalent Staphylococcus aureus glycoconjugated vaccine in adults on hemodialysis: Phase III randomized study. Hum. Vaccines Immunother. 2015, 11, 632–641. [Google Scholar] [CrossRef]
- Landrum, M.L.; Lalani, T.; Niknian, M.; Maguire, J.D.; Hospenthal, D.R.; Fattom, A.; Taylor, K.; Fraser, J.; Wilkins, K.; Ellis, M.W.; et al. Safety and immunogenicity of a recombinant Staphylococcus aureus alpha-toxoid and a recombinant Panton-Valentine leukocidin subunit, in healthy adults. Hum. Vaccines Immunother. 2017, 13, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Giersing, B.K.; Dastgheyb, S.S.; Modjarrad, K.; Moorthy, V. Status of vaccine research and development of vaccines for Staphylococcus aureus. Vaccine 2016, 34, 2962–2966. [Google Scholar] [CrossRef] [PubMed]
- Marshall, H.; Nissen, M.; Richmond, P.; Shakib, S.; Jiang, Q.; Cooper, D.; Rill, D.; Baber, J.; Eiden, J.; Gruber, W.C.; et al. Safety and immunogenicity of a booster dose of a 3-antigen Staphylococcus aureus vaccine (SA3Ag) in healthy adults: A randomized phase 1 study. J. Infect. 2016, 73, 437–454. [Google Scholar] [CrossRef] [PubMed]
- Frenck, R.W., Jr.; Creech, C.B.; Sheldon, E.A.; Seiden, D.J.; Kankam, M.K.; Baber, J.; Zito, E.; Hubler, R.; Eiden, J.; Severs, J.M.; et al. Safety, tolerability, and immunogenicity of a 4-antigen Staphylococcus aureus vaccine (SA4Ag): Results from a first-in-human randomised, placebo-controlled phase 1/2 study. Vaccine 2017, 35, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, C.S.; White, C.J.; Ibrahim, A.S.; Filler, S.G.; Fu, Y.; Yeaman, M.R.; Edwards, J.E., Jr.; Hennessey, J.P., Jr. NDV-3, a recombinant alum-adjuvanted vaccine for Candida and Staphylococcus aureus, is safe and immunogenic in healthy adults. Vaccine 2012, 30, 7594–7600. [Google Scholar] [CrossRef] [PubMed]
- Levy, J.; Licini, L.; Haelterman, E.; Moris, P.; Lestrate, P.; Damaso, S.; Van Belle, P.; Boutriau, D. Safety and immunogenicity of an investigational 4-component Staphylococcus aureus vaccine with or without AS03B adjuvant: Results of a randomized phase I trial. Hum. Vaccines Immunother. 2015, 11, 620–631. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.H.; Pasetti, M.F.; Adhikari, R.P.; Baughman, H.; Douglas, R.; El-Khorazaty, J.; Greenberg, N.; Holtsberg, F.W.; Liao, G.C.; Reymann, M.K.; et al. Safety and Immunogenicity of a Parenterally Administered, Structure-Based Rationally Modified Recombinant Staphylococcal Enterotoxin B Protein Vaccine, STEBVax. Clin. Vaccine Immunol. 2016, 23, 918–925. [Google Scholar] [CrossRef]
- Proctor, R.A. Challenges for a universal Staphylococcus aureus vaccine. Clin. Infect. Dis. 2012, 54, 1179–1186. [Google Scholar] [CrossRef]
- Daum, R.S.; Spellberg, B. Progress toward a Staphylococcus aureus vaccine. Clin. Infect. Dis. 2012, 54, 560–567. [Google Scholar] [CrossRef]
- Zuo, Q.F.; Yang, L.Y.; Feng, Q.; Lu, D.S.; Dong, Y.D.; Cai, C.Z.; Wu, Y.; Guo, Y.; Gu, J.; Zeng, H.; et al. Evaluation of the protective immunity of a novel subunit fusion vaccine in a murine model of systemic MRSA infection. PLoS ONE 2013, 8, e81212. [Google Scholar] [CrossRef]
- Jinyong, Z.; Xiaoli, Z.; Weijun, Z.; Ying, G.; Gang, G.; Xuhu, M.; Quanming, Z. Fusion expression and immunogenicity of Bordetella pertussis PTS1-FHA protein: Implications for the vaccine development. Mol. Biol. Rep. 2011, 38, 1957–1963. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.M. The bicinchoninic acid (BCA) assay for protein quantitation. Methods Mol. Biol. 1994, 32, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Gu, J.; Zou, J.; Lei, L.; Jing, H.; Zhang, J.; Zeng, H.; Zou, Q.; Lv, F.; Zhang, J. PA0833 Is an OmpA C-Like Protein That Confers Protection Against Pseudomonas aeruginosa Infection. Front. Microbiol. 2018, 9, 1062. [Google Scholar] [CrossRef] [PubMed]
- Kuklin, N.A.; Clark, D.J.; Secore, S.; Cook, J.; Cope, L.D.; McNeely, T.; Noble, L.; Brown, M.J.; Zorman, J.K.; Wang, X.M.; et al. A novel Staphylococcus aureus vaccine: Iron surface determinant B induces rapid antibody responses in rhesus macaques and specific increased survival in a murine S. aureus sepsis model. Infect. Immun. 2006, 74, 2215–2223. [Google Scholar] [CrossRef] [PubMed]
- Rauch, S.; Gough, P.; Kim, H.K.; Schneewind, O.; Missiakas, D. Vaccine protection of leukopenic mice against Staphylococcus aureus bloodstream infection. Infect. Immun. 2014, 82, 4889–4898. [Google Scholar] [CrossRef]
- Bubeck Wardenburg, J.; Schneewind, O. Vaccine protection against Staphylococcus aureus pneumonia. J. Exp. Med. 2008, 205, 287–294. [Google Scholar] [CrossRef]
- Bagnoli, F.; Fontana, M.R.; Soldaini, E.; Mishra, R.P.; Fiaschi, L.; Cartocci, E.; Nardi-Dei, V.; Ruggiero, P.; Nosari, S.; De Falco, M.G.; et al. Vaccine composition formulated with a novel TLR7-dependent adjuvant induces high and broad protection against Staphylococcus aureus. Proc. Natl. Acad. Sci. USA 2015, 112, 3680–3685. [Google Scholar] [CrossRef]
- Behrouz, B.; Mahdavi, M.; Amirmozafari, N.; Fatemi, M.J.; Irajian, G.; Bahroudi, M.; Hashemi, F.B. Immunogenicity of Pseudomonas aeruginosa recombinant b-type fagellin as a vaccine candidate: Protective efficacy in a murine burn wound sepsis model. Burns 2016. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, F.; Zhang, X.; Jing, H.; Ren, C.; Cai, C.; Dong, Y.; Zhang, Y.; Zou, Q.; Zeng, H. Protective Efficacy and Mechanism of Passive Immunization with Polyclonal Antibodies in a Sepsis Model of Staphylococcus aureus Infection. Sci. Rep. 2015, 5, 15553. [Google Scholar] [CrossRef]
- Kim, H.K.; Cheng, A.G.; Kim, H.Y.; Missiakas, D.M.; Schneewind, O. Nontoxigenic protein A vaccine for methicillin-resistant Staphylococcus aureus infections in mice. J. Exp. Med. 2010, 207, 1863–1870. [Google Scholar] [CrossRef]
- Goodyear, C.S.; Silverman, G.J. Death by a B cell superantigen: In vivo VH-targeted apoptotic supraclonal B cell deletion by a Staphylococcal Toxin. J. Exp. Med. 2003, 197, 1125–1139. [Google Scholar] [CrossRef] [PubMed]
- Thammavongsa, V.; Rauch, S.; Kim, H.K.; Missiakas, D.M.; Schneewind, O. Protein A-neutralizing monoclonal antibody protects neonatal mice against Staphylococcus aureus. Vaccine 2015, 33, 523–526. [Google Scholar] [CrossRef] [PubMed]
- Bhakdi, S.; Jursch, R.; Broker, M.; Ronneberger, H.; Hungerer, K.D. Functionally inactive S. aureus alpha-toxin containing a single amino acid substitution: Potential usefulness as a vaccine. Behring Inst. Mitt. 1994, 95, 80–84. [Google Scholar]
- Menzies, B.E.; Kernodle, D.S. Site-directed mutagenesis of the alpha-toxin gene of Staphylococcus aureus: Role of histidines in toxin activity in vitro and in a murine model. Infect. Immun. 1994, 62, 1843–1847. [Google Scholar] [CrossRef]
- Gaudin, C.F.; Grigg, J.C.; Arrieta, A.L.; Murphy, M.E. Unique heme-iron coordination by the hemoglobin receptor IsdB of Staphylococcus aureus. Biochemistry 2011, 50, 5443–5452. [Google Scholar] [CrossRef] [PubMed]
- Varshney, A.K.; Wang, X.; Macintyre, J.; Zollner, R.S.; Kelleher, K.; Kovalenko, O.V.; Pechuan, X.; Byrne, F.R.; Fries, B.C. Humanized Staphylococcal Enterotoxin B (SEB)-Specific Monoclonal Antibodies Protect From SEB Intoxication and Staphylococcus aureus Infections Alone or as Adjunctive Therapy with Vancomycin. J. Infect. Dis. 2014, 210, 973–981. [Google Scholar] [CrossRef]
- Boles, J.W.; Pitt, M.L.; LeClaire, R.D.; Gibbs, P.H.; Torres, E.; Dyas, B.; Ulrich, R.G.; Bavari, S. Generation of protective immunity by inactivated recombinant staphylococcal enterotoxin B vaccine in nonhuman primates and identification of correlates of immunity. Clin. Immunol. 2003, 108, 51–59. [Google Scholar] [CrossRef]
- Inskeep, T.K.; Stahl, C.; Odle, J.; Oakes, J.; Hudson, L.; Bost, K.L.; Piller, K.J. Oral vaccine formulations stimulate mucosal and systemic antibody responses against staphylococcal enterotoxin B in a piglet model. Clin. Vaccine Immunol. 2010, 17, 1163–1169. [Google Scholar] [CrossRef]
- Kehl-Fie, T.E.; Zhang, Y.; Moore, J.L.; Farrand, A.J.; Hood, M.I.; Rathi, S.; Chazin, W.J.; Caprioli, R.M.; Skaar, E.P. MntABC and MntH contribute to systemic Staphylococcus aureus infection by competing with calprotectin for nutrient manganese. Infect. Immun. 2013, 81, 3395–3405. [Google Scholar] [CrossRef]
- Salazar, N.; Castiblanco-Valencia, M.M.; da Silva, L.B.; de Castro, Í.A.; Monaris, D.; Masuda, H.P.; Barbosa, A.S.; Arêas, A.P.M. Staphylococcus aureus Manganese Transport Protein C (MntC) Is an Extracellular Matrix-and Plasminogen-Binding Protein. PLoS ONE 2014, 9, e112730. [Google Scholar] [CrossRef]
- Anderson, A.S.; Scully, I.L.; Timofeyeva, Y.; Murphy, E.; McNeil, L.K.; Mininni, T.; Nunez, L.; Carriere, M.; Singer, C.; Dilts, D.A.; et al. Staphylococcus aureus manganese transport protein C is a highly conserved cell surface protein that elicits protective immunity against S. aureus and Staphylococcus epidermidis. J. Infect. Dis. 2012, 205, 1688–1696. [Google Scholar] [CrossRef]
- O’Brien, E.C.; McLoughlin, R.M. Considering the ’Alternatives’ for Next-Generation Anti-Staphylococcus aureus Vaccine Development. Trends Mol. Med. 2019, 25, 171–184. [Google Scholar] [CrossRef]
- Mohamed, N.; Wang, M.Y.; Le Huec, J.C.; Liljenqvist, U.; Scully, I.L.; Baber, J.; Begier, E.; Jansen, K.U.; Gurtman, A.; Anderson, A.S. Vaccine development to prevent Staphylococcus aureus surgical-site infections. Br. J. Surg. 2017, 104, e41–e54. [Google Scholar] [CrossRef] [PubMed]
- Salgado-Pabon, W.; Schlievert, P.M. Models matter: The search for an effective Staphylococcus aureus vaccine. Nat. Rev. Microbiol. 2014, 12, 585–591. [Google Scholar] [CrossRef] [PubMed]
- McKee, A.S.; Munks, M.W.; MacLeod, M.K.; Fleenor, C.J.; Van Rooijen, N.; Kappler, J.W.; Marrack, P. Alum induces innate immune responses through macrophage and mast cell sensors, but these sensors are not required for alum to act as an adjuvant for specific immunity. J. Immunol. 2009, 183, 4403–4414. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Zhu, H.; Xia, X.; Liang, Z.; Ma, X.; Sun, B. Vaccine adjuvants: Understanding the structure and mechanism of adjuvanticity. Vaccine 2019, 37, 3167–3178. [Google Scholar] [CrossRef]
- McNeely, T.B.; Shah, N.A.; Fridman, A.; Joshi, A.; Hartzel, J.S.; Keshari, R.S.; Lupu, F.; DiNubile, M.J. Mortality among recipients of the Merck V710 Staphylococcus aureus vaccine after postoperative S. aureus infections: An analysis of possible contributing host factors. Hum. Vaccines Immunother. 2014, 10, 3513–3516. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Survival Rate (10 days) | |||||||
|---|---|---|---|---|---|---|---|---|
| V1 | V2 | V3 | V4 | V5 | V6 | Mean | ||
| Control | His buffer | 0 | 10% | 10% | 20% | 10% | 30% | 16% |
| AlPO4 | 30% | 25% | 20% | 30% | 20% | 30% | 25% | |
| Monovalent | mHIN2 | 10% | 30% | 40% | 30% | 25% | 30% | 27% |
| mSEB | 30% | 11% | 30% | 30% | 30% | 38% | 24% | |
| SpA5 | 70% | 20% | 30% | 35% | 40% | 35% | 40% | |
| MntC | 56% | 0% | 30% | 30% | 35% | 27% | 29% | |
| Bivalent | mHIN2 + mSEB | 45% | 38% | 30% | 60% | 80% | 90% | 60% |
| mHIN2 + SpA5 | 58% | 40% | 50% | 60% | 80% | 80% | 62% | |
| mHIN2 + MntC | 60% | 50% | 25% | 80% | 90% | 100% | 69% | |
| mSEB + SpA5 | 76% | 90% | 60% | 80% | 100% | 60% | 78% | |
| mSEB + MntC | 65% | 70% | 70% | 78% | 60% | 89% | 73% | |
| SpA5 + MntC | 73% | 70% | 80% | 80% | 80% | 90% | 80% | |
| Trivalent | mHIN2 + mSEB + SpA5 | 76% | 100% | 78% | 67% | 67% | 80% | 78% |
| mHIN2 + mSEB + MntC | 71% | 60% | 50% | 80% | 60% | 100% | 70% | |
| mHIN2 + SpA5 + MntC | 70% | 80% | 89% | 67% | 70% | 60% | 73% | |
| mSEB + SpA5 + MntC | 80% | 70% | 90% | 80% | 89% | 90% | 84% | |
| rFSAV | mHIN2 + mSEB + SpA5 + MntC | 90% | 80% | 90% | 85% | 85% | 90% | 87% |
| V710 [24] | IsdB | 70% | 55% | 50% | - | - | - | 58% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, H.; Yang, F.; Feng, Q.; Zhang, J.; Gu, J.; Jing, H.; Cai, C.; Xu, L.; Yang, X.; Xia, X.; et al. Rapid and Broad Immune Efficacy of a Recombinant Five-Antigen Vaccine against Staphylococcus aureus Infection in Animal Models. Vaccines 2020, 8, 134. https://doi.org/10.3390/vaccines8010134
Zeng H, Yang F, Feng Q, Zhang J, Gu J, Jing H, Cai C, Xu L, Yang X, Xia X, et al. Rapid and Broad Immune Efficacy of a Recombinant Five-Antigen Vaccine against Staphylococcus aureus Infection in Animal Models. Vaccines. 2020; 8(1):134. https://doi.org/10.3390/vaccines8010134
Chicago/Turabian StyleZeng, Hao, Feng Yang, Qiang Feng, Jinyong Zhang, Jiang Gu, Haiming Jing, Changzhi Cai, Liming Xu, Xi Yang, Xin Xia, and et al. 2020. "Rapid and Broad Immune Efficacy of a Recombinant Five-Antigen Vaccine against Staphylococcus aureus Infection in Animal Models" Vaccines 8, no. 1: 134. https://doi.org/10.3390/vaccines8010134
APA StyleZeng, H., Yang, F., Feng, Q., Zhang, J., Gu, J., Jing, H., Cai, C., Xu, L., Yang, X., Xia, X., Zeng, N., Fan, S., & Zou, Q. (2020). Rapid and Broad Immune Efficacy of a Recombinant Five-Antigen Vaccine against Staphylococcus aureus Infection in Animal Models. Vaccines, 8(1), 134. https://doi.org/10.3390/vaccines8010134