Repurposing Plant Virus Nanoparticles


Abstract
1. Introduction
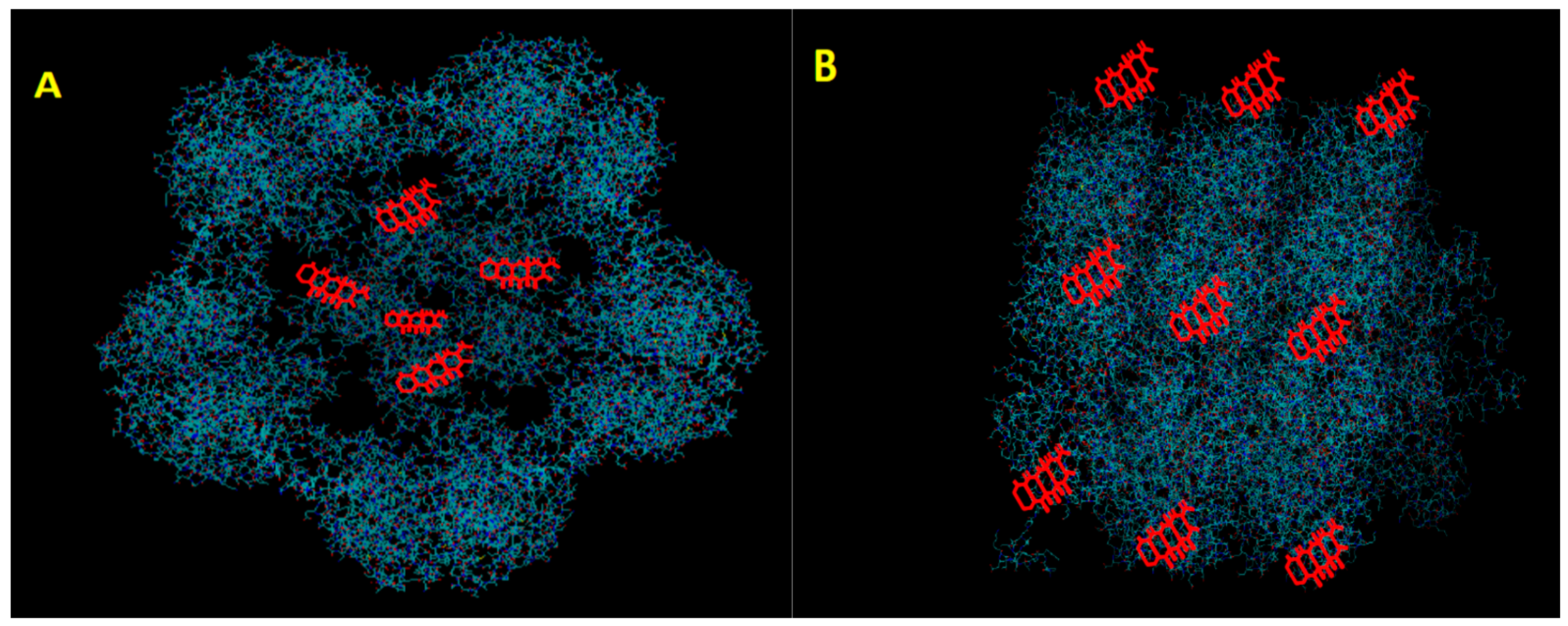
2. Icosahedral Virus Nanoparticles
3. Helical Virus Nanoparticles
4. Other Applications for Plant Virus Nanoparticles
5. Conclusions
Author Contributions
Conflicts of Interest
References
- Paul, M.; Reljic, R.; Klein, K.; Drake, P.M.; van Dolleweerd, C.; Pabst, M.; Windwarder, M.; Arcalis, E.; Stoger, E.; Altmann, F.; et al. Characterization of a plant-produced recombinant human secretory IgA with broad neutralizing activity against HIV. MAbs 2014, 6, 1585–1597. [Google Scholar] [CrossRef] [PubMed]
- Pua, T.L.; Chan, X.Y.; Loh, H.S.; Omar, A.R.; Yusibov, V.; Musiychuk, K.; Hall, A.C.; Coffin, M.V.; Shoji, Y.; Chichester, J.A.; et al. Purification and immunogenicity of hemagglutinin from highly pathogenic avian influenza virus H5N1 expressed in Nicotiana benthamiana. Hum. Vaccin. Immunother. 2017, 13, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Loh, H.S.; Green, B.J.; Yusibov, V. Using plants and modified plant viruses for the development of treatments for human diseases. Curr. Opin. Virol. 2017, 26, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Yusibov, V.; Streatfield, S.J.; Kushnir, N. Clinical development of plant-produced recombinant pharmaceuticals: Vaccines, antibodies and beyond. Hum. Vaccines 2011, 7, 313–321. [Google Scholar] [CrossRef]
- Sainsbury, F.; Lomonossoff, G.P. Transient expressions of synthetic biology in plants. Curr. Opin. Plant Biol. 2014, 19, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Aboul-Ata, A.A.; Vitti, A.; Nuzzaci, M.; El-Attar, A.K.; Piazzolla, G.; Tortorella, C.; Harandi, A.M.; Olson, O.; Wright, S.A.; Piazzolla, P. Plant-based vaccines: Novel and low-cost possible route for Mediterranean innovative vaccination strategies. Adv. Virus Res. 2014, 89, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Sun, H.; Lai, H.; Hurtado, J.; Chen, Q. Plant-produced Zika virus envelope protein elicits neutralizing immune responses that correlate with protective immunity against Zika virus in mice. Plant Biotechnol. J. 2017. [Google Scholar] [CrossRef] [PubMed]
- Mardanova, E.S.; Kotlyarov, R.Y.; Kuprianov, V.V.; Stepanova, L.A.; Tsybalova, L.M.; Lomonosoff, G.P.; Ravin, N.V. Rapid high-yield expression of a candidate influenza vaccine based on the ectodomain of M2 protein linked to flagellin in plants using viral vectors. BMC Biotechnol. 2015, 15, 42. [Google Scholar] [CrossRef] [PubMed]
- Gleba, Y.; Marillonnet, S.; Klimyuk, V. Design of safe and biologically contained transgenic plants: Tools and technologies for controlled transgene flow and expression. Biotechnol. Genet. Eng. Rev. 2004, 21, 325–368. [Google Scholar] [CrossRef] [PubMed]
- Gleba, Y.; Klimyuk, V.; Marillonnet, S. Magnifection—A new platform for expressing recombinant vaccines in plants. Vaccine 2005, 23, 2042–2048. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, K.B.; Han, S.S. Icosahedral plant viral nanoparticles-bioinspired synthesis of nanomaterials/nanostructures. Adv. Colloid Interface Sci. 2017, 248, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, K.B.; Han, S.S. Helical plant viral nanoparticles-bioinspired synthesis of nanomaterials and nanostructures. Bioinspir. Biomim. 2017, 12, 031001. [Google Scholar] [CrossRef] [PubMed]
- Czapar, A.E.; Steinmetz, N.F. Plant viruses and bacteriophages for drug delivery in medicine and biotechnology. Curr. Opin. Chem. Biol. 2017, 38, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Peruzzi, P.P.; Chiocca, E.A. Cancer immunotherapy: A vaccine from plant virus proteins. Nat. Nanotechnol. 2016, 11, 214–215. [Google Scholar] [CrossRef] [PubMed]
- Lico, C.; Benvenuto, E.; Baschieri, S. The Two-Faced Potato Virus X: From Plant Pathogen to Smart Nanoparticle. Front. Plant Sci. 2015, 6, 1009. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Myers, J.T.; Woods, S.E.; Gong, X.; Czapar, A.E.; Commandeur, U.; Huang, A.Y.; Levine, A.D.; Steinmetz, N.F. Plant viral nanoparticles-based HER2 vaccine: Immune response influenced by differential transport, localization and cellular interactions of particulate carriers. Biomaterials 2017, 121, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Lico, C.; Santi, L.; Twyman, R.M.; Pezzotti, M.; Avesani, L. The use of plants for the production of therapeutic human peptides. Plant Cell Rep. 2012, 31, 439–451. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, K.B.; Han, S.S. Genetic Modifications of Icosahedral Plant Virus-based Nanoparticles for Vaccine and Immunotherapy Applications. Curr. Protein Pept. Sci. 2017, 18, 1141–1151. [Google Scholar] [CrossRef] [PubMed]
- Berardi, A.; Evans, D.J.; Baldelli Bombelli, F.; Lomonossoff, G.P. Stability of plant virus-based nanocarriers in gastrointestinal fluids. Nanoscale 2017. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, M.J.; Plummer, E.M.; Rae, C.S.; Manchester, M. Interaction of Cowpea mosaic virus (CPMV) nanoparticles with antigen presenting cells in vitro and in vivo. PLoS ONE 2009, 4, e7981. [Google Scholar] [CrossRef] [PubMed]
- Saunders, K.; Sainsbury, F.; Lomonossoff, G.P. Efficient generation of cowpea mosaic virus empty virus-like particles by the proteolytic processing of precursors in insect cells and plants. Virology 2009, 393, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Lizotte, P.H.; Wen, A.M.; Sheen, M.R.; Fields, J.; Rojanasopondist, P.; Steinmetz, N.F.; Fiering, S. In situ vaccination with cowpea mosaic virus nanoparticles suppresses metastatic cancer. Nat. Nanotechnol. 2016, 11, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Madden, A.J.; Oberhardt, B.; Lockney, D.; Santos, C.; Vennam, P.; Arney, D.; Franzen, S.; Lommel, S.A.; Miller, C.R.; Gehrig, P.; et al. Pharmacokinetics and efficacy of doxorubicin-loaded plant virus nanoparticles in preclinical models of cancer. Nanomedicine 2017, 12, 2519–2532. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Wong, S.M.; Lim, L.Y. Folic acid-conjugated protein cages of a plant virus: A novel delivery platform for doxorubicin. Bioconjug. Chem. 2007, 18, 836–843. [Google Scholar] [CrossRef] [PubMed]
- Alemzadeh, E.; Izadpanah, K.; Ahmadi, F. Generation of recombinant protein shells of Johnson grass chlorotic stripe mosaic virus in tobacco plants and their use as drug carrier. J. Virol. Methods 2017, 248, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Masarapu, H.; Patel, B.K.; Chariou, P.L.; Hu, H.; Gulati, N.M.; Carpenter, B.L.; Ghiladi, R.A.; Shukla, S.; Steinmetz, N.F. Physalis Mottle Virus-Like Particles as Nanocarriers for Imaging Reagents and Drugs. Biomacromolecules 2017, 18, 4141–4153. [Google Scholar] [CrossRef] [PubMed]
- Tusé, D. Safety of plant-made pharmaceuticals: Product development and regulatory considerations based on case studies of two autologous human cancer vaccines. Hum. Vaccines 2011, 7, 322–330. [Google Scholar] [CrossRef]
- Tusé, D.; Ku, N.; Bendandi, M.; Becerra, C.; Collins, R., Jr.; Langford, N.; Sancho, S.I.; López-Díaz de Cerio, A.; Pastor, F.; Kandzia, R.; et al. Clinical Safety and Immunogenicity of Tumor-Targeted, Plant-Made Id-KLH Conjugate Vaccines for Follicular Lymphoma. Biomed. Res. Int. 2015, 2015, 648143. [Google Scholar] [CrossRef]
- Kernan, D.L.; Wen, A.M.; Pitek, A.S.; Steinmetz, N.F. Featured Article: Delivery of chemotherapeutic vcMMAE using tobacco mosaic virus nanoparticles. Exp. Biol. Med. 2017, 242, 1405–1411. [Google Scholar] [CrossRef] [PubMed]
- Komarova, T.V.; Kosorukov, V.S.; Frolova, O.Y.; Petrunia, I.V.; Skrypnik, K.A.; Gleba, Y.Y.; Dorokhov, Y.L. Plant-made trastuzumab (herceptin) inhibits HER2/Neu+ cell proliferation and retards tumor growth. PLoS ONE 2011, 6, e17541. [Google Scholar] [CrossRef] [PubMed]
- Bruckman, M.A.; Czapar, A.E.; VanMeter, A.; Randolph, L.N.; Steinmetz, N.F. Tobacco mosaic virus-based protein nanoparticles and nanorods for chemotherapy delivery targeting breast cancer. J. Control. Release 2016, 231, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Franke, C.E.; Czapar, A.E.; Patel, R.B.; Steinmetz, N.F. Tobacco Mosaic Virus-Delivered Cisplatin Restores Efficacy in Platinum-Resistant Ovarian Cancer Cells. Mol. Pharm. 2017. [Google Scholar] [CrossRef] [PubMed]
- Jobsri, J.; Allen, A.; Rajagopal, D.; Shipton, M.; Kanyuka, K.; Lomonossoff, G.P.; Ottensmeier, C.; Diebold, S.S.; Stevenson, F.K.; Savelyeva, N. Plant virus particles carrying tumour antigen activate TLR7 and Induce high levels of protective antibody. PLoS ONE 2015, 10, e0118096. [Google Scholar] [CrossRef] [PubMed]
- Babin, C.; Majeau, N.; Leclerc, D. Engineering of papaya mosaic virus (PapMV) nanoparticles with a CTL epitope derived from influenza NP. J. Nanobiotechnol. 2013, 11, 10. [Google Scholar] [CrossRef] [PubMed]
- Lacasse, P.; Denis, J.; Lapointe, R.; Leclerc, D.; Lamarre, A. Novel plant virus-based vaccine induces protective cytotoxic T-lymphocyte-mediated antiviral immunity through dendritic cell maturation. J. Virol. 2008, 82, 785–794. [Google Scholar] [CrossRef] [PubMed]
- Hanafi, L.A.; Bolduc, M.; Gagné, M.E.; Dufour, F.; Langelier, Y.; Boulassel, M.R.; Routy, J.P.; Leclerc, D.; Lapointe, R. Two distinct chimeric potexviruses share antigenic cross-presentation properties of MHC class I epitopes. Vaccine 2010, 28, 5617–5626. [Google Scholar] [CrossRef] [PubMed]
- Lebel, M.È.; Chartrand, K.; Tarrab, E.; Savard, P.; Leclerc, D.; Lamarre, A. Potentiating Cancer Immunotherapy Using Papaya Mosaic Virus-Derived Nanoparticles. Nano Lett. 2016, 16, 1826–1832. [Google Scholar] [CrossRef] [PubMed]
- Lebel, M.È.; Daudelin, J.F.; Chartrand, K.; Tarrab, E.; Kalinke, U.; Savard, P.; Labrecque, N.; Leclerc, D.; Lamarre, A. Nanoparticle adjuvant sensing by TLR7 enhances CD8+ T cell-mediated protection from Listeria monocytogenes infection. J. Immunol. 2014, 192, 1071–1078. [Google Scholar] [CrossRef] [PubMed]
- Steinmetz, N.F.; Cho, C.-F.; Ablack, A.; Lewis, J.D.; Manchester, M. Cowpea mosaic virus nanoparticles target surface vimentin on cancer cells. Nanomedicine 2011, 6, 351–364. [Google Scholar] [CrossRef] [PubMed]
- PDQ Adult Treatment Editorial Board. Adult Non-Hodgkin Lymphoma Treatment (PDQ®): Health Professional Version; PDQ Cancer Information Summaries Bethesda (MD); National Cancer Institute: Bethesda, MD, USA, 2002–2017.
- McCormick, A.A. Tobacco derived cancer vaccines for non-Hodgkin’s lymphoma: Perspectives and progress. Hum. Vaccin. 2011, 7, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Bendandi, M.; Marillonnet, S.; Kandzia, R.; Thieme, F.; Nickstadt, A.; Herz, S.; Fröde, R.; Inogés, S.; Lòpez-Dìaz de Cerio, A.; Soria, E.; et al. Rapid, high-yield production in plants of individualized idiotype vaccines for non-Hodgkin’s lymphoma. Ann. Oncol. 2010, 21, 2420–2427. [Google Scholar] [CrossRef] [PubMed]
- Chotprakaikiat, W.; Allen, A.; Bui-Minh, D.; Harden, E.; Jobsri, J.; Cavallo, F.; Gleba, Y.; Stevenson, F.K.; Ottensmeier, C.; Klimyuk, V.; et al. A plant-expressed conjugate vaccine breaks CD4+ tolerance and induces potent immunity against metastatic Her2+ breast cancer. Oncoimmunology 2016, 5, e1166323. [Google Scholar] [CrossRef] [PubMed]
- Le, D.H.T.; Hu, H.; Commandeur, U.; Steinmetz, N.F. Chemical addressability of potato virus X for its applications in bio/nanotechnology. J. Struct. Biol. 2017, 200, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.L.; Murray, A.A.; Le, D.H.T.; Sheen, M.R.; Shukla, S.; Commandeur, U.; Fiering, S.; Steinmetz, N.F. Combination of Plant Virus Nanoparticle-Based in Situ Vaccination with Chemotherapy Potentiates Antitumor Response. Nano Lett. 2017, 17, 4019–4028. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Ablack, A.L.; Wen, A.M.; Lee, K.L.; Lewis, J.D.; Steinmetz, N.F. Increased tumor homing and tissue penetration of the filamentous plant viral nanoparticle Potato virus X. Mol. Pharm. 2012, 10, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Lebel, M.È.; Chartrand, K.; Leclerc, D.; Lamarre, A. Plant Viruses as Nanoparticle-Based Vaccines and Adjuvants. Vaccines (Basel) 2015, 3, 620–637. [Google Scholar] [CrossRef] [PubMed]
- Carignan, D.; Herblot, S.; Laliberté-Gagné, M.È.; Bolduc, M.; Duval, M.; Savard, P.; Leclerc, D. Activation of innate immunity in primary human cells using a plant virus derived nanoparticle TLR7/8 agonist. Nanomedicine 2017. [Google Scholar] [CrossRef] [PubMed]
- González-Gamboa, I.; Manrique, P.; Sánchez, F.; Ponz, F. Plant-made potyvirus-like particles used for log-increasing antibody sensing capacity. J. Biotechnol. 2017, 254, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.L.; Carpenter, B.L.; Wen, A.M.; Ghiladi, R.A.; Steinmetz, N.F. High aspect ratio nanotubes formed by tobacco mosaic virus for delivery of photodynamic agents targeting melanoma. ACS Biomater. Sci. Eng. 2016, 2, 838–844. [Google Scholar] [CrossRef] [PubMed]
- Pitek, A.S.; Wang, Y.; Gulati, S.; Gao, H.; Stewart, P.L.; Simon, D.I.; Steinmetz, N.F. Elongated Plant Virus-Based Nanoparticles for Enhanced Delivery of Thrombolytic Therapies. Mol. Pharm. 2017, 14, 3815–3823. [Google Scholar] [CrossRef] [PubMed]
- Lauria, I.; Dickmeis, C.; Röder, J.; Beckers, M.; Rütten, S.; Lin, Y.Y.; Commandeur, U.; Fischer, H. Engineered Potato virus X nanoparticles support hydroxyapatite nucleation for improved bone tissue replacement. Acta Biomater. 2017, 62, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Wen, A.M.; Lee, K.L.; Yildiz, I.; Bruuckman, M.A.; Shukla, S.; Steinmetz, N.F. Vrial nanoparticles for in vivo tumor inmaging. J. Vis. Exp. 2012, 16, e4352. [Google Scholar]
- Röder, J.; Fischer, R.; Commandeur, U. Adoption of the 2A Ribosomal Skip Principle to Tobacco Mosaic Virusfor Peptide Display. Front. Plant Sci. 2017, 8, 1125. [Google Scholar] [CrossRef] [PubMed]
- Yildiz, I.; Lee, K.L.; Chen, K.; Shukla, S.; Steinmetz, N.F. Infusion of imaging and therapeutic molecules into the plant virus-based carrier cowpea mosaic virus: Cargo-loading and delivery. J. Control. Release 2013, 172, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Cho, C.F.; Shukla, S.; Simpson, E.J.; Steinmetz, N.F.; Luyt, L.G.; Lewis, J.D. Molecular targeted viral nanoparticles as tools for imaging cancer. Methods Mol. Biol. 2014, 1108, 211–230. [Google Scholar] [PubMed]
- Shukla, S.; DiFranco, N.A.; Wen, A.M.; Commandeur, U.; Steinmetz, N.F. To Target or Not to Target: Active vs. Passive Tumor Homing of Filamentous Nanoparticles Based on Potato virus X. Cell. Mol. Bioeng. 2015, 8, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Bruckman, M.A.; Lauren, N. Biodistribution, pharmacokinetics, and blood compatibility of native and PEGylated tobacco mosaic virus nano-rods and -spheres in mice. Virology 2014, 449, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Pitek, A.S.; Jameson, S.A.; Veliz, F.A.; Shukla, S.; Steinmetz, N.F. Serum albumin ‘camouflage’ of plant virus based nanoparticles prevents their antibody recognition and enhances pharmacokinetics. Biomaterials 2016, 89, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Esfandiari, N.; Arzanani, M.K.; Soleimani, M.; Kohi-Habibi, M.; Svendsen, W.E. A new application of plant virus nanoparticles as drug delivery in breast cancer. Tumor Biol. 2016, 37, 1229–1236. [Google Scholar] [CrossRef] [PubMed]
- Lico, C.; Schoubben, A.; Baschieri, S.; Blasi, P.; Santi, L. Nanoparticles in biomedicine: New insights from plant viruses. Curr. Med. Chem. 2013, 20, 3471–3487. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| A. Icosahedral Viruses | ||
|---|---|---|
| Plant Virus Nanoparticle | Application Tested | Reference |
| Cowpea mosaic virus (CPMV) | Solid tumors | [20,21,22] |
| Red clover necrotic mosaic virus (RCNMV) encapsulating DOX | Ovarian cancer, melanoma | [23] |
| Hibiscus chlorotic ringspot virus (HCRSV) encapsulating DOX, PSA and PAA | Ovarian cancer | [24] |
| Johnson grass chlorotic stripe mosaic virus (JgCSMV) encapsulating DOX | ||
| Physalis mottle virus (PhMV) encasiulating DOX and MTX | Untested | [25] |
| B. Helical Viruses | ||
| Tobacco mosaic virus (TMV) immunoglobulins, also nanoparticle carrying vcMMAE, Her2, cisplatin, porphyrin dynamic sensors | Breast, ovarian, prostate cancers | [26] |
| Potato virus X (PVX) | ||
| Papaya mosaic vírus (PapMV) | Non Hodgkins Lymphoma, Ovarian cancer cells, melanoma cells | [27,28,29,30,31,32] |
| Murine B cell lymphoma, Breast cancer cells | [30,33] | |
| Ovarian cancer cells | [34,35,36,37] | |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hefferon, K.L. Repurposing Plant Virus Nanoparticles. Vaccines 2018, 6, 11. https://doi.org/10.3390/vaccines6010011
Hefferon KL. Repurposing Plant Virus Nanoparticles. Vaccines. 2018; 6(1):11. https://doi.org/10.3390/vaccines6010011
Chicago/Turabian StyleHefferon, Kathleen L. 2018. "Repurposing Plant Virus Nanoparticles" Vaccines 6, no. 1: 11. https://doi.org/10.3390/vaccines6010011
APA StyleHefferon, K. L. (2018). Repurposing Plant Virus Nanoparticles. Vaccines, 6(1), 11. https://doi.org/10.3390/vaccines6010011