
Targeting Host Cell Surface Nucleolin for RSV Therapy: Challenges and Opportunities



Abstract
1. Introduction
2. Materials and Methods
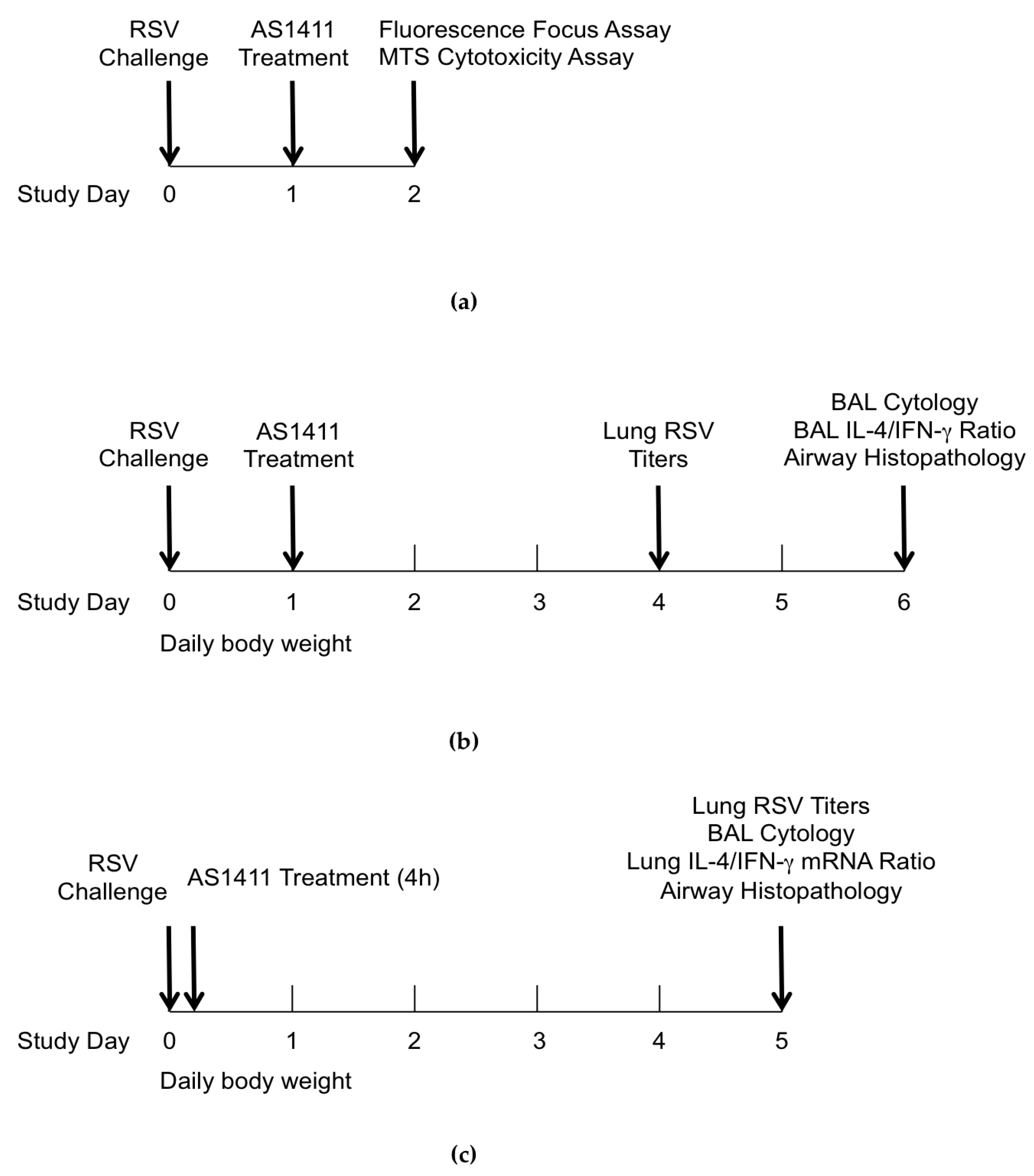
2.1. General Study Design
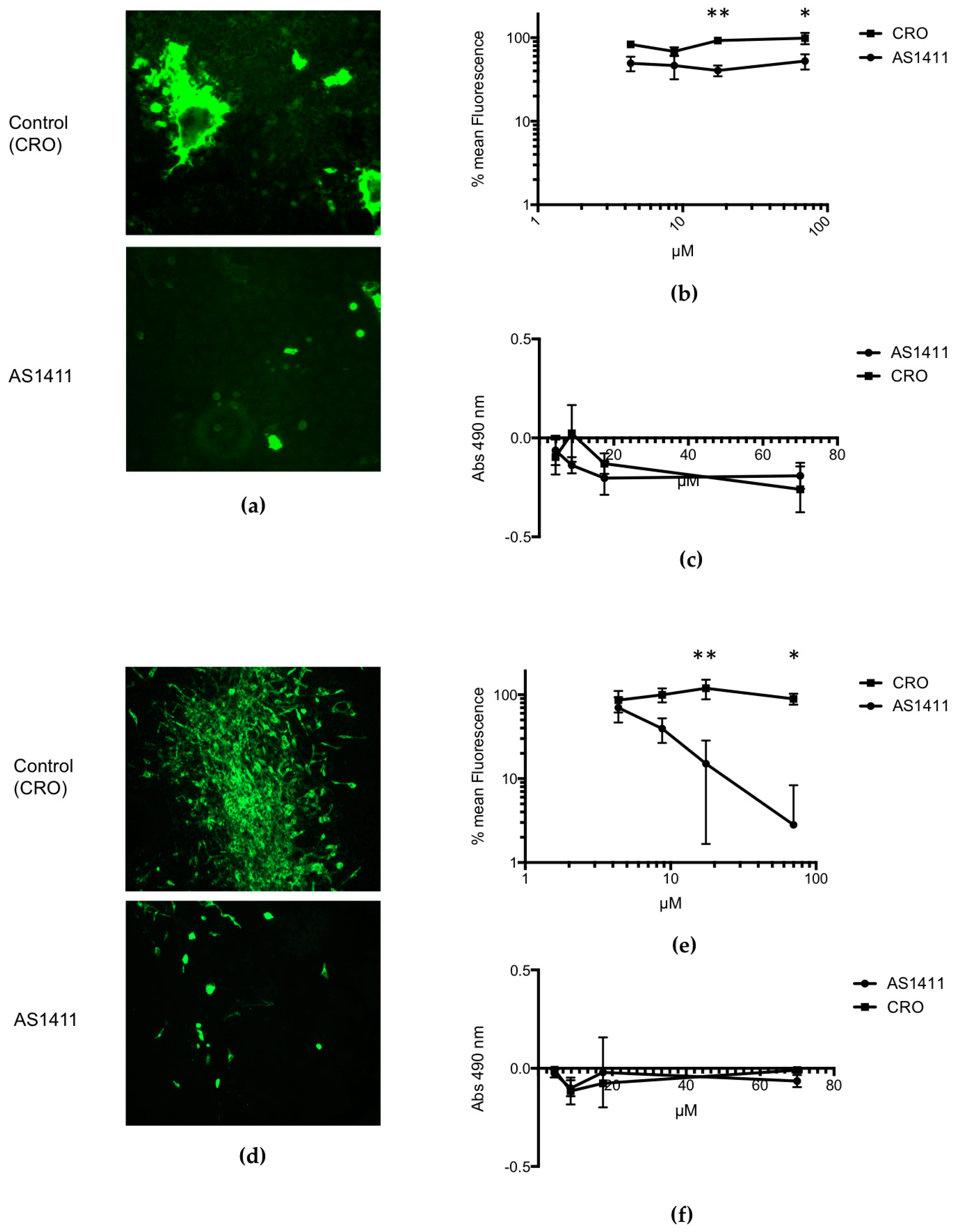
2.2. In Vitro Experiments: Administration of AS1411 to Rsv-Infected HEp-2 Cells or MDCK Cells
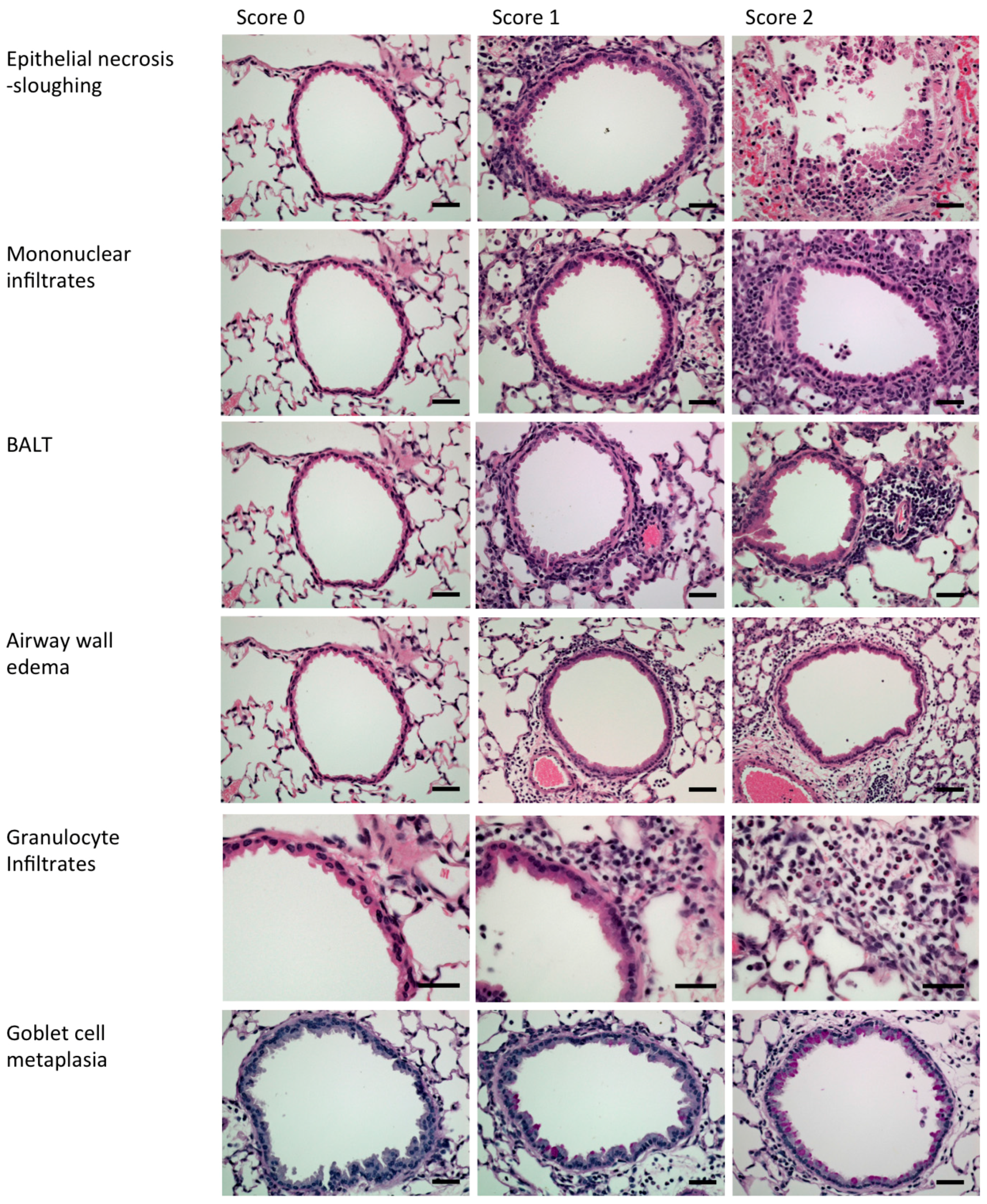
2.3. Mouse Experiments
2.4. Cotton Rat Experiments
2.5. Statistical Analyses
3. Results
3.1. In Vitro Experiments
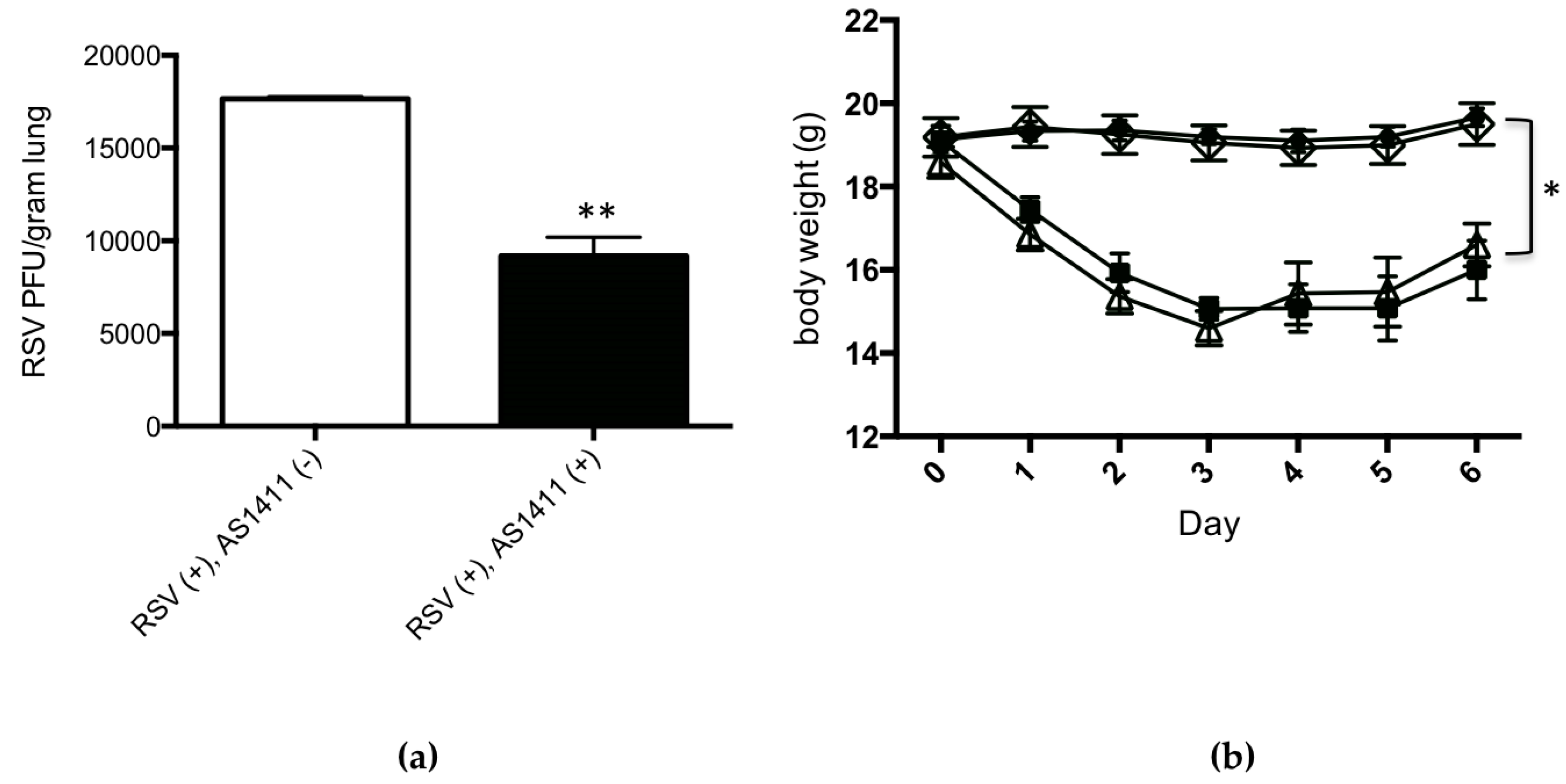
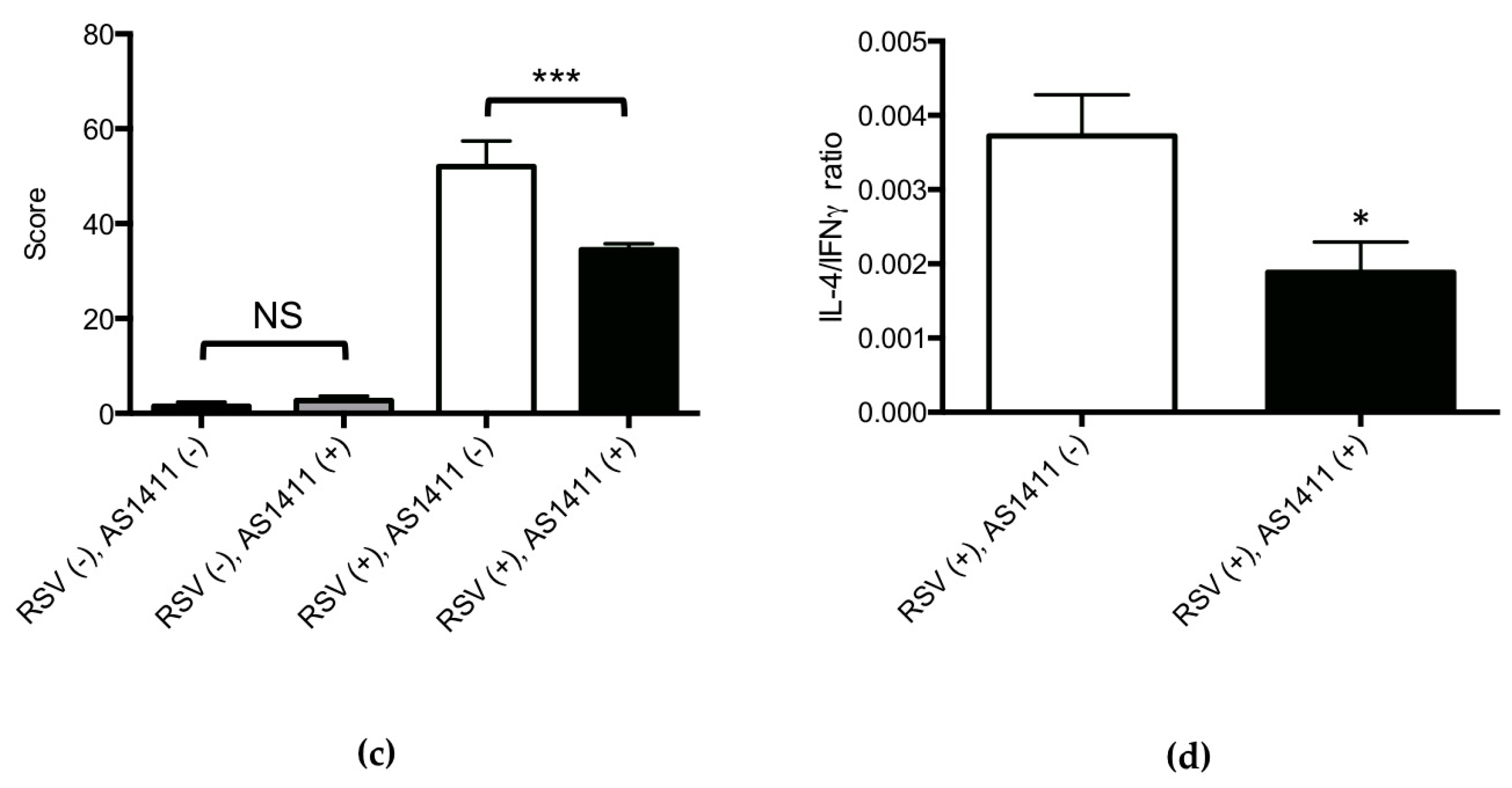
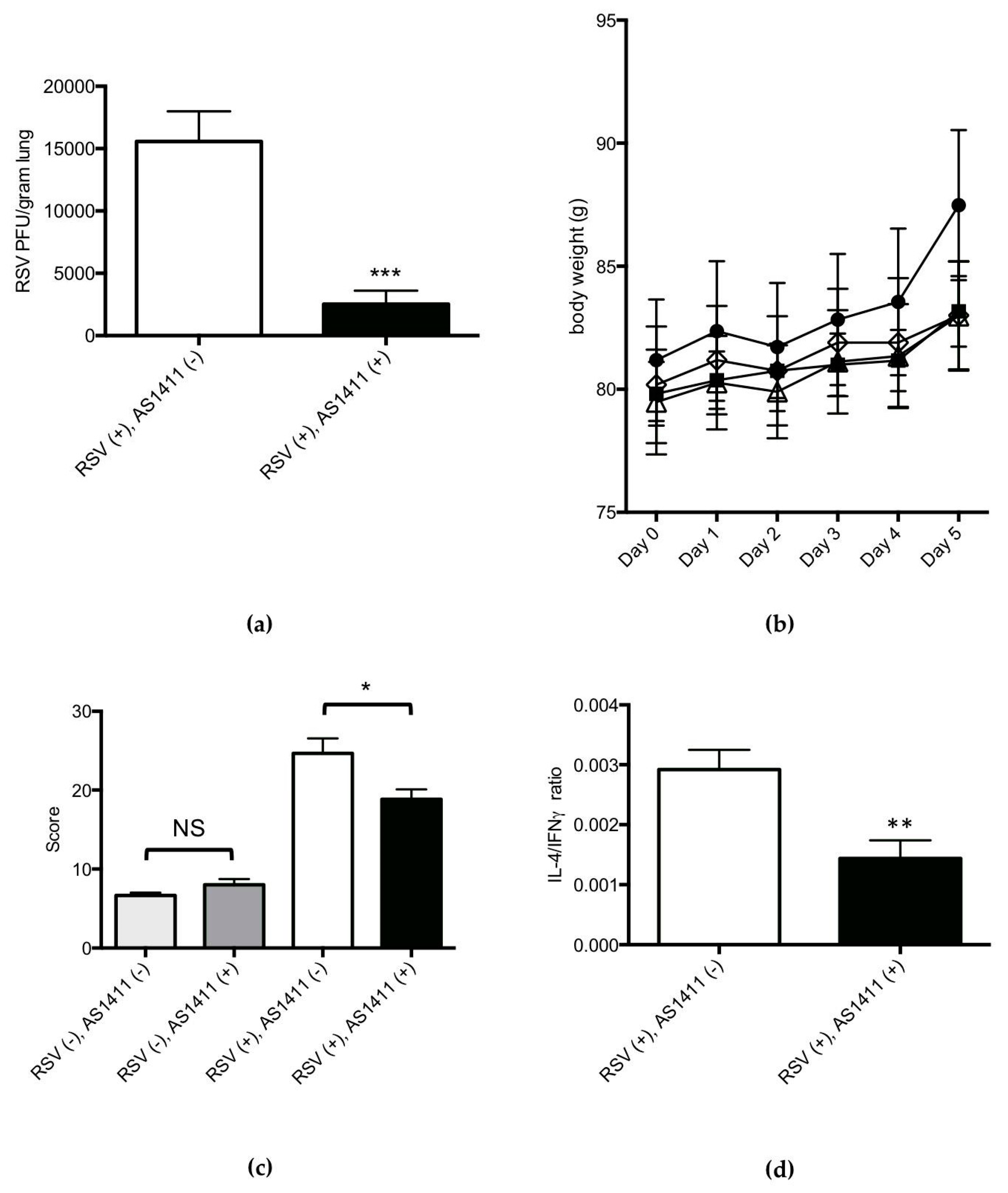
3.2. Mouse Experiments
3.3. Cotton Rat Experiments
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Adams, M.J.; Lefkowitz, E.J.; King, A.M.Q.; Harrach, B.; Harrison, R.L.; Knowles, N.J.; Kropinski, A.M.; Krupovic, M.; Kuhn, J.H.; Mushegian, A.R.; et al. Changes to taxonomy and the International Code of Virus Classification and Nomenclature ratified by the International Committee on Taxonomy of Viruses (2017). Arch. Virol. 2017, 162, 2505–2538. [Google Scholar] [CrossRef] [PubMed]
- Walsh, E.E. Respiratory Syncytial Virus Infection: An Illness for All Ages. Clin. Chest Med. 2017, 38, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Novavax RSV F Vaccine Fails Phase III Trial. Available online: http://www.genengnews.com/gen-news-highlights/novavax-rsv-f-vaccine-fails-phase-iii-trial/81253207 (accessed on 14 September 2017).
- Zhu, Q.; Patel, N.K.; McAuliffe, J.M.; Zhu, W.; Wachter, L.; McCarthy, M.P.; Suzich, J.A. Natural polymorphisms and resistance-associated mutations in the fusion protein of respiratory syncytial virus (RSV): Effects on RSV susceptibility to palivizumab. J. Infect. Dis. 2012, 205, 635–638. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; McAuliffe, J.M.; Patel, N.K.; Palmer-Hill, F.J.; Yang, C.F.; Liang, B.; Su, L.; Zhu, W.; Wachter, L.; Wilson, S.; et al. Analysis of respiratory syncytial virus preclinical and clinical variants resistant to neutralization by monoclonal antibodies palivizumab and/or motavizumab. J. Infect. Dis. 2011, 203, 674–682. [Google Scholar] [CrossRef] [PubMed]
- Papenburg, J.; Carbonneau, J.; Hamelin, M.E.; Isabel, S.; Bouhy, X.; Ohoumanne, N.; Dery, P.; Paes, B.A.; Corbeil, J.; Bergeron, M.G.; et al. Molecular evolution of respiratory syncytial virus fusion gene, Canada, 2006–2010. Emerg. Infect. Dis. 2012, 18, 120–124. [Google Scholar] [CrossRef] [PubMed]
- DeVincenzo, J.P.; Hall, C.B.; Kimberlin, D.W.; Sanchez, P.J.; Rodriguez, W.J.; Jantausch, B.A.; Corey, L.; Kahn, J.S.; Englund, J.A.; Suzich, J.A.; et al. Surveillance of clinical isolates of respiratory syncytial virus for palivizumab (Synagis)-resistant mutants. J. Infect. Dis. 2004, 190, 975–978. [Google Scholar] [CrossRef] [PubMed]
- Graham, B.S.; Modjarrad, K.; McLellan, J.S. Novel antigens for RSV vaccines. Curr. Opin. Immunol. 2015, 35, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Fearns, R.; Deval, J. New antiviral approaches for respiratory syncytial virus and other mononegaviruses: Inhibiting the RNA polymerase. Antiviral. Res. 2016, 134, 63–76. [Google Scholar] [CrossRef] [PubMed]
- Battles, M.B.; Langedijk, J.P.; Furmanova-Hollenstein, P.; Chaiwatpongsakorn, S.; Costello, H.M.; Kwanten, L.; Vranckx, L.; Vink, P.; Jaensch, S.; Jonckers, T.H.; et al. Molecular mechanism of respiratory syncytial virus fusion inhibitors. Nat. Chem. Biol. 2016, 12, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Heylen, E.; Neyts, J.; Jochmans, D. Drug candidates and model systems in respiratory syncytial virus antiviral drug discovery. Biochem. Pharmacol. 2017, 127, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.; MacCarthy, T. Potential for evolution of complex defense strategies in a multi-scale model of virus-host coevolution. BMC Evol. Biol. 2016, 16, 233. [Google Scholar] [CrossRef] [PubMed]
- Keener, A.B. Host with the most: Targeting host cells instead of pathogens to fight infectious disease. Nat. Med. 2017, 23, 528–531. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Yen, H.L. Targeting the host or the virus: Current and novel concepts for antiviral approaches against influenza virus infection. Antiviral. Res. 2012, 96, 391–404. [Google Scholar] [CrossRef] [PubMed]
- Tayyari, F.; Marchant, D.; Moraes, T.J.; Duan, W.; Mastrangelo, P.; Hegele, R.G. Identification of nucleolin as a cellular receptor for human respiratory syncytial virus. Nat. Med. 2011, 17, 1132–1135. [Google Scholar] [CrossRef] [PubMed]
- Soundararajan, S.; Chen, W.; Spicer, E.K.; Courtenay-Luck, N.; Fernandes, D.J. The nucleolin targeting aptamer AS1411 destabilizes Bcl-2 messenger RNA in human breast cancer cells. Cancer Res. 2008, 68, 2358–2365. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Song, C.; Jiang, C.; Shen, X.; Qiao, Q.; Hu, Y. Nucleolin targeting AS1411 modified protein nanoparticle for antitumor drugs delivery. Mol. Pharm. 2013, 10, 3555–3563. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, J.E.; Bambury, R.M.; Van Allen, E.M.; Drabkin, H.A.; Lara, P.N., Jr.; Harzstark, A.L.; Wagle, N.; Figlin, R.A.; Smith, G.W.; Garraway, L.A.; et al. A phase II trial of AS1411 (a novel nucleolin-targeted DNA aptamer) in metastatic renal cell carcinoma. Investig. New Drugs 2014, 32, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Balcarova-Stander, J.; Pfeiffer, S.E.; Fuller, S.D.; Simons, K. Development of cell surface polarity in the epithelial Madin-Darby canine kidney (MDCK) cell line. EMBO J. 1984, 3, 2687–2694. [Google Scholar] [PubMed]
- Shakeri, A.; Mastrangelo, P.; Griffin, J.K.; Moraes, T.J.; Hegele, R.G. Respiratory syncytial virus receptor expression in the mouse and viral tropism. Histol. Histopathol. 2015, 30, 401–411. [Google Scholar] [PubMed]
- Mastrangelo, P.; Hegele, R.G. RSV fusion: Time for a new model. Viruses 2013, 5, 873–885. [Google Scholar] [CrossRef] [PubMed]
- Brock, S.C.; Goldenring, J.R.; Crowe, J.E., Jr. Apical recycling systems regulate directional budding of respiratory syncytial virus from polarized epithelial cells. Proc. Natl. Acad. Sci. USA 2003, 100, 15143–15148. [Google Scholar] [CrossRef] [PubMed]
- Roberts, S.R.; Compans, R.W.; Wertz, G.W. Respiratory syncytial virus matures at the apical surfaces of polarized epithelial cells. J. Virol. 1995, 69, 2667–2673. [Google Scholar] [PubMed]
- Openshaw, P.J. The mouse model of respiratory syncytial virus disease. Curr. Top. Microbiol. Immunol. 2013, 372, 359–369. [Google Scholar] [PubMed]
- Boukhvalova, M.S.; Prince, G.A.; Blanco, J.C. The cotton rat model of respiratory viral infections. Biologicals 2009, 37, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Rivera, C.A.; Gomez, R.S.; Diaz, R.A.; Cespedes, P.F.; Espinoza, J.A.; Gonzalez, P.A.; Riedel, C.A.; Bueno, S.M.; Kalergis, A.M. Novel therapies and vaccines against the human respiratory syncytial virus. Expert. Opin. Investig. Drugs 2015, 24, 1613–1630. [Google Scholar] [CrossRef] [PubMed]
- Feyzi, R.; Boskabady, M.H.; Seyedhosseini Tamijani, S.M.; Rafatpanah, H.; Rezaei, S.A. The Effect of Safranal on Th1/Th2 Cytokine Balance. Iran. J. Immunol. 2016, 13, 263–273. [Google Scholar] [PubMed]
- Zahzeh, M.R.; Loukidi, B.; Meziane, W.; Haddouche, M.; Mesli, N.; Zouaoui, Z.; Aribi, M. Relationship between NADPH and Th1/Th2 ratio in patients with non- Hodgkin lymphoma who have been exposed to pesticides. J. Blood Med. 2015, 6, 99–107. [Google Scholar] [PubMed]
- Kaan, P.M.; Hegele, R.G. Interaction between respiratory syncytial virus and particulate matter in guinea pig alveolar macrophages. Am. J. Respir. Cell Mol. Biol. 2003, 28, 697–704. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, R.; Ward, M.; Bright, H.; Priest, R.; Foster, M.R.; Hurle, M.; Blair, E.; Bird, M. Isolation and characterisation of potential respiratory syncytial virus receptor(s) on epithelial cells. Microbes Infect. 2003, 5, 123–133. [Google Scholar] [CrossRef]
- Ueba, O. Respiratory syncytial virus. I. Concentration and purification of the infectious virus. Acta Med. Okayama 1978, 32, 265–272. [Google Scholar] [PubMed]
- Hegele, R.G.; Robinson, P.J.; Gonzalez, S.; Hogg, J.C. Production of acute bronchiolitis in guinea-pigs by human respiratory syncytial virus. Eur. Respir. J. 1993, 6, 1324–1331. [Google Scholar] [PubMed]
- Aherne, W.; Bird, T.; Court, S.D.; Gardner, P.S.; McQuillin, J. Pathological changes in virus infections of the lower respiratory tract in children. J. Clin. Pathol. 1970, 23, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Blanco, J.C.; Richardson, J.Y.; Darnell, M.E.; Rowzee, A.; Pletneva, L.; Porter, D.D.; Prince, G.A. Cytokine and chemokine gene expression after primary and secondary respiratory syncytial virus infection in cotton rats. J. Infect. Dis. 2002, 185, 1780–1785. [Google Scholar] [CrossRef] [PubMed]
- Blanco, J.C.; Pletneva, L.; Boukhvalova, M.; Richardson, J.Y.; Harris, K.A.; Prince, G.A. The cotton rat: An underutilized animal model for human infectious diseases can now be exploited using specific reagents to cytokines, chemokines, and interferons. J. Interferon Cytokine Res. 2004, 24, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Becker, Y. Respiratory syncytial virus (RSV) evades the human adaptive immune system by skewing the Th1/Th2 cytokine balance toward increased levels of Th2 cytokines and IgE, markers of allergy—A review. Virus Genes 2006, 33, 235–252. [Google Scholar] [PubMed]
- Boyoglu-Barnum, S.; Chirkova, T.; Todd, S.O.; Barnum, T.R.; Gaston, K.A.; Jorquera, P.; Haynes, L.M.; Tripp, R.A.; Moore, M.L.; Anderson, L.J. Prophylaxis with a respiratory syncytial virus (RSV) anti-G protein monoclonal antibody shifts the adaptive immune response to RSV rA2-line19F infection from Th2 to Th1 in BALB/c mice. J. Virol. 2014, 88, 10569–10583. [Google Scholar] [CrossRef] [PubMed]
- Vaux-Peretz, F.; Meignier, B. Comparison of lung histopathology and bronchoalveolar lavage cytology in mice and cotton rats infected with respiratory syncytial virus. Vaccine 1990, 8, 543–548. [Google Scholar] [CrossRef]
- Reyes-Reyes, E.M.; Teng, Y.; Bates, P.J. A new paradigm for aptamer therapeutic AS1411 action: Uptake by macropinocytosis and its stimulation by a nucleolin-dependent mechanism. Cancer Res. 2010, 70, 8617–8629. [Google Scholar] [CrossRef] [PubMed]
- Ireson, C.R.; Kelland, L.R. Discovery and development of anticancer aptamers. Mol. Cancer Ther. 2006, 5, 2957–2962. [Google Scholar] [CrossRef] [PubMed]
- Perron, M.; Stray, K.; Kinkade, A.; Theodore, D.; Lee, G.; Eisenberg, E.; Sangi, M.; Gilbert, B.E.; Jordan, R.; Piedra, P.A.; et al. GS-5806 Inhibits a Broad Range of Respiratory Syncytial Virus Clinical Isolates by Blocking the Virus-Cell Fusion Process. Antimicrob. Agents Chemother. 2015, 60, 1264–1273. [Google Scholar] [CrossRef] [PubMed]
- Su, P.Y.; Wang, Y.F.; Huang, S.W.; Lo, Y.C.; Wang, Y.H.; Wu, S.R.; Shieh, D.B.; Chen, S.H.; Wang, J.R.; Lai, M.D.; et al. Cell surface nucleolin facilitates enterovirus 71 binding and infection. J. Virol. 2015, 89, 4527–4538. [Google Scholar] [CrossRef] [PubMed]
- Hovanessian, A.G. Midkine, a cytokine that inhibits HIV infection by binding to the cell surface expressed nucleolin. Cell Res. 2006, 16, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Thongtan, T.; Wikan, N.; Wintachai, P.; Rattanarungsan, C.; Srisomsap, C.; Cheepsunthorn, P.; Smith, D.R. Characterization of putative Japanese encephalitis virus receptor molecules on microglial cells. J. Med. Virol. 2012, 84, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.; Shan, X.; Lapeyre, B.; Melese, T. The amino terminus of mammalian nucleolin specifically recognizes SV40 T-antigen type nuclear localization sequences. Eur. J. Cell Biol. 1993, 62, 13–21. [Google Scholar] [PubMed]
- Perrone, R.; Butovskaya, E.; Lago, S.; Garzino-Demo, A.; Pannecouque, C.; Palu, G.; Richter, S.N. The G-quadruplex-forming aptamer AS1411 potently inhibits HIV-1 attachment to the host cell. Int. J. Antimicrob. Agents 2016, 47, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Stobart, C.C.; Rostad, C.A.; Ke, Z.; Dillard, R.S.; Hampton, C.M.; Strauss, J.D.; Yi, H.; Hotard, A.L.; Meng, J.; Pickles, R.J.; et al. A live RSV vaccine with engineered thermostability is immunogenic in cotton rats despite high attenuation. Nat. Commun. 2016, 7, 13916. [Google Scholar] [CrossRef] [PubMed]
- Mbuagbaw, L.; Mursleen, S.; Irlam, J.H.; Spaulding, A.B.; Rutherford, G.W.; Siegfried, N. Efavirenz or nevirapine in three-drug combination therapy with two nucleoside or nucleotide-reverse transcriptase inhibitors for initial treatment of HIV infection in antiretroviral-naive individuals. Cochrane Database Syst. Rev. 2016, 12, CD004246. [Google Scholar] [PubMed]
- Kohli, A.; Shaffer, A.; Sherman, A.; Kottilil, S. Treatment of hepatitis C: A systematic review. JAMA 2014, 312, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Simoes, E.A.; DeVincenzo, J.P.; Boeckh, M.; Bont, L.; Crowe, J.E., Jr.; Griffiths, P.; Hayden, F.G.; Hodinka, R.L.; Smyth, R.L.; Spencer, K.; et al. Challenges and opportunities in developing respiratory syncytial virus therapeutics. J. Infect. Dis. 2015, 211, S1–S20. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RSV (−), AS1411 (−) | RSV (−), AS1411 (+) | RSV (+) AS1411 (−) | RSV (+), AS1411 (+) | Statistical Analysis b | ||
|---|---|---|---|---|---|---|
| Mice | Total cell count (×104/mL) | 4.47 ± 0.95 | 5.57 ± 1.18 | 30.60 ± 11.95 | 14.48 ± 3.97 | S |
| % granulocytes | 1.45 ± 0.66 | 2.69 ± 0.63 | 20.32 ± 7.84 | 23.31 ± 9.26 | S | |
| % lymphocytes | 9.79 ± 2.36 | 9.28 ± 2.63 | 44.99 ± 0.18 | 37.66 ± 9.33 | S | |
| % macrophages | 88.72 ± 0.20 | 88.05 ± 0.19 | 34.90 ± 0.14 | 39.00 ± 10.49 | S | |
| Cotton rats | Total cell count (×105/mL) | 2.96 ± 0.73 | 4.56 ± 2.17 | 2.03 ± 1.18 | 3.63 ± 0.98 | NS |
| % granulocytes | 26.6 ± 12.7 | 25.14 ± 13.11 | 21.37 ± 10.80 | 29.95 ± 13.94 | NS | |
| % lymphocytes | 1.48 ± 1.01 | 0.77 ± 0.56 | 1.12 ± 0.60 | 2.60 ± 2.06 | NS | |
| % macrophages | 71.88 ± 13.22 | 74.09 ± 13.57 | 77.51 ± 10.99 | 67.45 ± 12.26 | NS |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mastrangelo, P.; Norris, M.J.; Duan, W.; Barrett, E.G.; Moraes, T.J.; Hegele, R.G. Targeting Host Cell Surface Nucleolin for RSV Therapy: Challenges and Opportunities. Vaccines 2017, 5, 27. https://doi.org/10.3390/vaccines5030027
Mastrangelo P, Norris MJ, Duan W, Barrett EG, Moraes TJ, Hegele RG. Targeting Host Cell Surface Nucleolin for RSV Therapy: Challenges and Opportunities. Vaccines. 2017; 5(3):27. https://doi.org/10.3390/vaccines5030027
Chicago/Turabian StyleMastrangelo, Peter, Michael J. Norris, Wenming Duan, Edward G. Barrett, Theo J. Moraes, and Richard G. Hegele. 2017. "Targeting Host Cell Surface Nucleolin for RSV Therapy: Challenges and Opportunities" Vaccines 5, no. 3: 27. https://doi.org/10.3390/vaccines5030027
APA StyleMastrangelo, P., Norris, M. J., Duan, W., Barrett, E. G., Moraes, T. J., & Hegele, R. G. (2017). Targeting Host Cell Surface Nucleolin for RSV Therapy: Challenges and Opportunities. Vaccines, 5(3), 27. https://doi.org/10.3390/vaccines5030027