
Immunological Evaluation of Recent MUC1 Glycopeptide Cancer Vaccines


Abstract
:1. Introduction
2. Different Targeting Mechanisms and Strategies
3. MUC1 Glycopeptide with Different Protein Carriers
4. Multicomponent Fully Synthetic MUC1 Vaccines
5. Use of Tn, STn and TF Analogs to Enhance Vaccine Stability
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ADCC | Antibody-dependent cellular cytotoxicity |
| APC | Antigen presenting cells |
| AuNP | Gold nanoparticles |
| BCG | Bacillus Calmette-Guérin |
| BSA | Bovine serum albumin |
| CDC | Complement-dependent cytotoxicity |
| CTB | Cholera toxin B subunit |
| CTL | Cytotoxic T lymphocyte |
| DDA | Dioctadecylammonium bromide |
| IFA | Incomplete Freund’s adjuvant |
| KLH | Keyhole limpet hemocyanin |
| MHC | Major histocompatibility complex antigen |
| MPL | Monophosphoryl lipid A |
| MUC1 | Mucin 1 |
| ODN | Oligodeoxynucleotide |
| P(HPMA) | Poly(N-(2-hydroxypropyl) methacrylamide |
| QS-21 | Quillaja saponaria extract 21 |
| Rha | Rhamnose |
| TACA | Tumor associated carbohydrate antigen |
| TF | Thomsen-Friedenreich |
| TLR | Toll-like receptor |
| Tregs | Regulatory T cells |
| TTox | Tetanus toxoid |
| VNTR | Variable number tandem repeat |
References
- Cheever, M.A.; Allison, J.P.; Ferris, A.S.; Finn, O.J.; Hastings, B.M.; Hecht, T.T.; Mellman, I.; Prindiville, S.A.; Viner, J.L.; Weiner, L.M.; et al. The prioritization of cancer antigens: A national cancer institute pilot project for the acceleration of translational research. Clin. Cancer Res. 2009, 15, 5323–5337. [Google Scholar]
- Cazet, A.; Julien, S.; Bobowski, M.; Burchell, J.; Delannoy, P. Tumour-associated carbohydrate antigens in breast cancer. Breast Cancer Res. 2010. [Google Scholar] [CrossRef]
- Ju, T.; Cummings, R.D. A unique molecular chaperone cosmc required for activity of the mammalian core 1 beta 3-galactosyltransferase. Proc. Natl. Acad. Sci. USA 2002, 99, 16613–16618. [Google Scholar]
- Sakurai, J.; Hattori, N.; Nakajima, M.; Moriya, T.; Suzuki, T.; Yokoyama, A.; Kohno, N. Differential expression of the glycosylated forms of MUC1 during lung development. Eur. J. Histochem. 2007, 51, 95–102. [Google Scholar]
- Sellers, T.A.; Huang, Y.; Cunningham, J.; Goode, E.L.; Sutphen, R.; Vierkant, R.A.; Kelemen, L.E.; Fredericksen, Z.S.; Liebow, M.; Pankratz, V.S.; et al. Association of single nucleotide polymorphisms in glycosylation genes with risk of epithelial ovarian cancer. Cancer Epidemiol. Biomark. Prev. 2008, 17, 397–404. [Google Scholar]
- Patani, N.; Jiang, W.; Mokbel, K. Prognostic utility of glycosyltransferase expression in breast cancer. Cancer Genom. Proteom. 2008, 5, 333–340. [Google Scholar]
- Meany, D.L.; Chan, D.W. Aberrant glycosylation associated with enzymes as cancer biomarkers. Clin. Proteom. 2011. [Google Scholar] [CrossRef]
- Wu, C.; Guo, X.; Wang, W.; Wang, Y.; Shan, Y.; Zhang, B.; Song, W.; Ma, S.; Ge, J.; Deng, H.; et al. N-acetylgalactosaminyltransferase-14 as a potential biomarker for breast cancer by immunohistochemistry. BMC Cancer 2010. [Google Scholar] [CrossRef]
- Gomes, J.; Marcos, N.T.; Berois, N.; Osinaga, E.; Magalhaes, A.; Pinto-de-Sousa, J.; Almeida, R.; Gartner, F.; Reis, C.A. Expression of UDP-N-acetyl-d-galactosamine: Polypeptide N-acetylgalactosaminyltransferase-6 in gastric mucosa, intestinal metaplasia, and gastric carcinoma. J. Histochem. Cytochem. 2009, 57, 79–86. [Google Scholar]
- Handerson, T.; Camp, R.; Harigopal, M.; Rimm, D.; Pawelek, J. Beta1,6-branched oligosaccharides are increased in lymph node metastases and predict poor outcome in breast carcinoma. Clin. Cancer Res. 2005, 11, 2969–2973. [Google Scholar]
- Julien, S.; Adriaenssens, E.; Ottenberg, K.; Furlan, A.; Courtand, G.; Vercoutter-Edouart, A.S.; Hanisch, F.G.; Delannoy, P.; Le Bourhis, X. ST6GalNAc I expression in MDA-MB-231 breast cancer cells greatly modifies their O-glycosylation pattern and enhances their tumourigenicity. Glycobiology 2006, 16, 54–64. [Google Scholar]
- Schneider, F.; Kemmner, W.; Haensch, W.; Franke, G.; Gretschel, S.; Karsten, U.; Schlag, P.M. Overexpression of sialyltransferase CMP-sialic acid:Galbeta1,3GalNAc-R alpha6-sialyltransferase is related to poor patient survival in human colorectal carcinomas. Cancer Res. 2001, 61, 4605–4611. [Google Scholar]
- Ryan, S.O.; Turner, M.S.; Gariepy, J.; Finn, O.J. Tumor antigen epitopes interpreted by the immune system as self or abnormal-self differentially affect cancer vaccine responses. Cancer Res. 2010, 70, 5788–5796. [Google Scholar]
- Rabinovich, G.A.; van Kooyk, Y.; Cobb, B.A. Glycobiology of immune responses. Ann. N. Y. Acad. Sci. 2012, 1253, 1–15. [Google Scholar]
- Tempero, R.M.; VanLith, M.L.; Morikane, K.; Rowse, G.J.; Gendler, S.J.; Hollingsworth, M.A. CD4+ lymphocytes provide MUC1-specific tumor immunity in vivo that is undetectable in vitro and is absent in MUC1 transgenic mice. J. Immunol. 1998, 161, 5500–5506. [Google Scholar]
- Tempero, R.M.; Rowse, G.J.; Gendler, S.J.; Hollingsworth, M.A. Passively transferred anti-MUC1 antibodies cause neither autoimmune disorders nor immunity against transplanted tumors in MUC1 transgenic mice. Int. J. Cancer 1999, 80, 595–599. [Google Scholar]
- Von Mensdorff-Pouilly, S.; Petrakou, E.; Kenemans, P.; van Uffelen, K.; Verstraeten, A.A.; Snijdewint, F.G.; van Kamp, G.J.; Schol, D.J.; Reis, C.A.; Price, M.R.; et al. Reactivity of natural and induced human antibodies to MUC1 mucin with MUC1 peptides and n-acetylgalactosamine (GalNAc) peptides. Int. J. Cancer 2000, 86, 702–712. [Google Scholar]
- Soares, M.M.; Mehta, V.; Finn, O.J. Three different vaccines based on the 140-amino acid MUC1 peptide with seven tandemly repeated tumor-specific epitopes elicit distinct immune effector mechanisms in wild-type versus MUC1-transgenic mice with different potential for tumor rejection. J. Immunol. 2001, 166, 6555–6563. [Google Scholar]
- Acres, B.; Apostolopoulos, V.; Balloul, J.M.; Wreschner, D.; Xing, P.X.; Ali-Hadji, D.; Bizouarne, N.; Kieny, M.P.; McKenzie, I.F. MUC1-specific immune responses in human MUC1 transgenic mice immunized with various human MUC1 vaccines. Cancer Immunol. Immunother. 2000, 48, 588–594. [Google Scholar]
- Ninkovic, T.; Hanisch, F.G. O-glycosylated human MUC1 repeats are processed in vitro by immunoproteasomes. J. Immunol. 2007, 179, 2380–2388. [Google Scholar]
- Lakshminarayanan, V.; Supekar, N.T.; Wei, J.; McCurry, D.B.; Dueck, A.C.; Kosiorek, H.E.; Trivedi, P.P.; Bradley, J.M.; Madsen, C.S.; Pathangey, L.B.; et al. MUC1 vaccines, comprised of glycosylated or non-glycosylated peptides or tumor-derived MUC1, can circumvent immunoediting to control tumor growth in MUC1 transgenic mice. PLoS ONE 2016, 11, e0145920. [Google Scholar]
- Barnd, D.L.; Lan, M.S.; Metzgar, R.S.; Finn, O.J. Specific, major histocompatibility complex-unrestricted recognition of tumor-associated mucins by human cytotoxic T cells. Proc. Natl. Acad. Sci. USA 1989, 86, 7159–7163. [Google Scholar]
- Gaidzik, N.; Westerlind, U.; Kunz, H. The development of synthetic antitumour vaccines from mucin glycopeptide antigens. Chem. Soc. Rev. 2013, 42, 4421–4442. [Google Scholar]
- Palitzsch, B.; Hartmann, S.; Stergiou, N.; Glaffig, M.; Schmitt, E.; Kunz, H. A fully synthetic four-component antitumor vaccine consisting of a mucin glycopeptide antigen combined with three different T-helper-cell epitopes. Angew. Chem. Int. Ed. Engl. 2014, 53, 14245–14249. [Google Scholar]
- Rakoff-Nahoum, S.; Medzhitov, R. Toll-like receptors and cancer. Nat. Rev. Cancer 2009, 9, 57–63. [Google Scholar]
- Sakaguchi, S. Naturally arising Foxp3-expressing CD25+CD4+ regulatory T cells in immunological tolerance to self and non-self. Nat. Immunol. 2005, 6, 345–352. [Google Scholar]
- Zou, W. Regulatory T cells, tumour immunity and immunotherapy. Nat. Rev. Immunol. 2006, 6, 295–307. [Google Scholar]
- Colombo, M.P.; Piconese, S. Regulatory—Cell inhibition versus depletion: The right choice in cancer immunotherapy. Nat. Rev. Cancer 2007, 7, 880–887. [Google Scholar]
- Ghiringhelli, F.; Puig, P.E.; Roux, S.; Parcellier, A.; Schmitt, E.; Solary, E.; Kroemer, G.; Martin, F.; Chauffert, B.; Zitvogel, L. Tumor cells convert immature myeloid dendritic cells into TGF-beta-secreting cells inducing CD4+CD25+ regulatory T cell proliferation. J. Exp. Med. 2005, 202, 919–929. [Google Scholar]
- Zhang, Y.; Luo, F.; Cai, Y.; Liu, N.; Wang, L.; Xu, D.; Chu, Y. TLR1/TLR2 agonist induces tumor regression by reciprocal modulation of effector and regulatory T cells. J. Immunol. 2011, 186, 1963–1969. [Google Scholar]
- Gathuru, J.K.; Koide, F.; Ragupathi, G.; Adams, J.L.; Kerns, R.T.; Coleman, T.P.; Livingston, P.O. Identification of DHBcAg as a potent carrier protein comparable to KLH for augmenting MUC1 antigenicity. Vaccine 2005, 23, 4727–4733. [Google Scholar]
- Gilewski, T.; Adluri, S.; Ragupathi, G.; Zhang, S.; Yao, T.J.; Panageas, K.; Moynahan, M.; Houghton, A.; Norton, L.; Livingston, P.O. Vaccination of high-risk breast cancer patients with mucin-1 (MUC1) keyhole limpet hemocyanin conjugate plus QS-21. Clin. Cancer Res. 2000, 6, 1693–1701. [Google Scholar]
- Kim, S.K.; Ragupathi, G.; Cappello, S.; Kagan, E.; Livingston, P.O. Effect of immunological adjuvant combinations on the antibody and T-cell response to vaccination with MUC1-KLH and GD3-KLH conjugates. Vaccine 2000, 19, 530–537. [Google Scholar]
- Kim, S.K.; Ragupathi, G.; Musselli, C.; Choi, S.J.; Park, Y.S.; Livingston, P.O. Comparison of the effect of different immunological adjuvants on the antibody and T-cell response to immunization with MUC1-KLH and GD3-KLH conjugate cancer vaccines. Vaccine 1999, 18, 597–603. [Google Scholar]
- Ragupathi, G.; Cappello, S.; Yi, S.S.; Canter, D.; Spassova, M.; Bornmann, W.G.; Danishefsky, S.J.; Livingston, P.O. Comparison of antibody titers after immunization with monovalent or tetravalent KLH conjugate vaccines. Vaccine 2002, 20, 1030–1038. [Google Scholar]
- Cai, H.; Huang, Z.H.; Shi, L.; Sun, Z.Y.; Zhao, Y.F.; Kunz, H.; Li, Y.M. Variation of the glycosylation pattern in MUC1 glycopeptide BSA vaccines and its influence on the immune response. Angew. Chem. Int. Ed. Engl. 2012, 51, 1719–1723. [Google Scholar]
- Hoffmann-Roder, A.; Johannes, M. Synthesis of a MUC1-glycopeptide-BSA conjugate vaccine bearing the 3′-deoxy-3′-fluoro-Thomsen-Friedenreich antigen. Chem. Commun. (Camb.) 2011, 47, 9903–9905. [Google Scholar]
- Hoffmann-Roder, A.; Kaiser, A.; Wagner, S.; Gaidzik, N.; Kowalczyk, D.; Westerlind, U.; Gerlitzki, B.; Schmitt, E.; Kunz, H. Synthetic antitumor vaccines from tetanus toxoid conjugates of MUC1 glycopeptides with the Thomsen-Friedenreich antigen and a fluorine-substituted analogue. Angew. Chem. Int. Ed. Engl. 2010, 49, 8498–8503. [Google Scholar]
- Fang, F.; Ma, J.; Ni, W.; Wang, F.; Sun, X.; Li, Y.; Li, Q.; Xie, F.; Wang, J.; Zhai, R.; et al. MUC1 and maltosebinding protein recombinant fusion protein combined with Bacillus Calmette-Guerin induces MUC1 specific and nonspecific antitumor immunity in mice. Mol. Med. Rep. 2014, 10, 1056–1064. [Google Scholar]
- Yuan, S.; Shi, C.; Ling, R.; Wang, T.; Wang, H.; Han, W. Immunization with two recombinant Bacillus Calmette-Guerin vaccines that combine the expression of multiple tandem repeats of mucin-1 and colony stimulating-factor suppress breast tumor growth in mice. J. Cancer Res. Clin. Oncol. 2010, 136, 1359–1367. [Google Scholar]
- Yuan, S.; Shi, C.; Liu, L.; Han, W. MUC1-based recombinant Bacillus Calmette-Guerin vaccines as candidates for breast cancer immunotherapy. Expert Opin. Biol. Ther. 2010, 10, 1037–1048. [Google Scholar]
- Wang, Y.; Liu, C.; Xia, Q.; Wang, P.; Li, B.; Lu, Z.; Sun, J.; Wu, H.; Yu, B.; Wu, J.; et al. Antitumor effect of adenoviral vector prime protein boost immunity targeting the MUC1 VNTRs. Oncol. Rep. 2014, 31, 1437–1444. [Google Scholar]
- Lu, W.; Qiu, L.; Yan, Z.; Lin, Z.; Cao, M.; Hu, C.; Wang, Z.; Wang, J.; Yu, Y.; Cheng, X.; et al. Cytotoxic T cell responses are enhanced by antigen design involving the presentation of MUC1 peptide on cholera toxin B subunit. Oncotarget 2015, 6, 34537–34548. [Google Scholar]
- Kimura, T.; Finn, O.J. MUC1 immunotherapy is here to stay. Expert Opin. Biol. Ther. 2013, 13, 35–49. [Google Scholar]
- Cai, H.; Huang, Z.-H.; Shi, L.; Zou, P.; Zhao, Y.-F.; Kunz, H.; Li, Y.-M. Synthesis of Tn/T antigen MUC1 glycopeptide BSA conjugates and their evaluation as vaccines. Eur. J. Org. Chem. 2011, 2011, 3685–3689. [Google Scholar]
- Dziadek, S.; Kowalczyk, D.; Kunz, H. Synthetic vaccines consisting of tumor-associated MUC1 glycopeptide antigens and bovine serum albumin. Angew. Chem. Int. Ed. Engl. 2005, 44, 7624–7630. [Google Scholar]
- Cai, H.; Chen, M.S.; Sun, Z.Y.; Zhao, Y.F.; Kunz, H.; Li, Y.M. Self-adjuvanting synthetic antitumor vaccines from MUC1 glycopeptides conjugated to T-cell epitopes from tetanus toxoid. Angew. Chem. Int. Ed. Engl. 2013, 52, 6106–6110. [Google Scholar]
- Broderson, J.R. A retrospective review of lesions associated with the use of Freund’s adjuvant. Lab. Anim. Sci. 1989, 39, 400–405. [Google Scholar]
- Johannes, M.; Reindl, M.; Gerlitzki, B.; Schmitt, E.; Hoffmann-Roder, A. Synthesis and biological evaluation of a novel MUC1 glycopeptide conjugate vaccine candidate comprising a 4’-deoxy-4’-fluoro-Thomsen-Friedenreich epitope. Beilstein J. Org. Chem. 2015, 11, 155–161. [Google Scholar]
- Ragupathi, G.; Koide, F.; Livingston, P.O.; Cho, Y.S.; Endo, A.; Wan, Q.; Spassova, M.K.; Keding, S.J.; Allen, J.; Ouerfelli, O.; et al. Preparation and evaluation of unimolecular pentavalent and hexavalent antigenic constructs targeting prostate and breast cancer: A synthetic route to anticancer vaccine candidates. J. Am. Chem. Soc. 2006, 128, 2715–2725. [Google Scholar]
- Zhu, J.; Wan, Q.; Lee, D.; Yang, G.; Spassova, M.K.; Ouerfelli, O.; Ragupathi, G.; Damani, P.; Livingston, P.O.; Danishefsky, S.J. From synthesis to biologics: Preclinical data on a chemistry derived anticancer vaccine. J. Am. Chem. Soc. 2009, 131, 9298–9303. [Google Scholar]
- Kaiser, A.; Gaidzik, N.; Westerlind, U.; Kowalczyk, D.; Hobel, A.; Schmitt, E.; Kunz, H. A synthetic vaccine consisting of a tumor-associated sialyl-T(n)-MUC1 tandem-repeat glycopeptide and tetanus toxoid: Induction of a strong and highly selective immune response. Angew. Chem. Int. Ed. Engl. 2009, 48, 7551–7555. [Google Scholar]
- Palitzsch, B.; Gaidzik, N.; Stergiou, N.; Stahn, S.; Hartmann, S.; Gerlitzki, B.; Teusch, N.; Flemming, P.; Schmitt, E.; Kunz, H. A synthetic glycopeptide vaccine for the induction of a monoclonal antibody that differentiates between normal and tumor mammary cells and enables the diagnosis of human pancreatic cancer. Angew. Chem. Int. Ed. Engl. 2016, 55, 2894–2898. [Google Scholar]
- Buskas, T.; Li, Y.; Boons, G.J. The immunogenicity of the tumor-associated antigen Lewis(y) may be suppressed by a bifunctional cross-linker required for coupling to a carrier protein. Chemistry 2004, 10, 3517–3524. [Google Scholar]
- Herzenberg, L.A.; Tokuhisa, T.; Herzenberg, L.A. Carrier-priming leads to hapten-specific suppression. Nature 1980, 285, 664–667. [Google Scholar]
- Kagan, E.; Ragupathi, G.; Yi, S.S.; Reis, C.A.; Gildersleeve, J.; Kahne, D.; Clausen, H.; Danishefsky, S.J.; Livingston, P.O. Comparison of antigen constructs and carrier molecules for augmenting the immunogenicity of the monosaccharide epithelial cancer antigen Tn. Cancer Immunol. Immunother. 2005, 54, 424–430. [Google Scholar]
- Schutze, M.P.; Leclerc, C.; Jolivet, M.; Audibert, F.; Chedid, L. Carrier-induced epitopic suppression, a major issue for future synthetic vaccines. J. Immunol. 1985, 135, 2319–2322. [Google Scholar]
- Cai, H.; Sun, Z.Y.; Chen, M.S.; Zhao, Y.F.; Kunz, H.; Li, Y.M. Synthetic multivalent glycopeptide-lipopeptide antitumor vaccines: Impact of the cluster effect on the killing of tumor cells. Angew. Chem. Int. Ed. Engl. 2014, 53, 1699–1703. [Google Scholar]
- Geraci, C.; Consoli, G.M.; Granata, G.; Galante, E.; Palmigiano, A.; Pappalardo, M.; Di Puma, S.D.; Spadaro, A. First self-adjuvant multicomponent potential vaccine candidates by tethering of four or eight MUC1 antigenic immunodominant PDTRP units on a calixarene platform: Synthesis and biological evaluation. Bioconjug. Chem. 2013, 24, 1710–1720. [Google Scholar]
- McDonald, D.M.; Wilkinson, B.L.; Corcilius, L.; Thaysen-Andersen, M.; Byrne, S.N.; Payne, R.J. Synthesis and immunological evaluation of self-adjuvanting MUC1-macrophage activating lipopeptide 2 conjugate vaccine candidates. Chem. Commun. (Camb.) 2014, 50, 10273–10276. [Google Scholar]
- Borsutzky, S.; Kretschmer, K.; Becker, P.D.; Muhlradt, P.F.; Kirschning, C.J.; Weiss, S.; Guzman, C.A. The mucosal adjuvant macrophage-activating lipopeptide-2 directly stimulates B lymphocytes via the TLR2 without the need of accessory cells. J. Immunol. 2005, 174, 6308–6313. [Google Scholar]
- Nicholls, E.F.; Madera, L.; Hancock, R.E. Immunomodulators as adjuvants for vaccines and antimicrobial therapy. Ann. N. Y. Acad. Sci. 2010, 1213, 46–61. [Google Scholar]
- Korsholm, K.S.; Agger, E.M.; Foged, C.; Christensen, D.; Dietrich, J.; Andersen, C.S.; Geisler, C.; Andersen, P. The adjuvant mechanism of cationic dimethyldioctadecylammonium liposomes. Immunology 2007, 121, 216–226. [Google Scholar]
- Persing, D.H.; Coler, R.N.; Lacy, M.J.; Johnson, D.A.; Baldridge, J.R.; Hershberg, R.M.; Reed, S.G. Taking toll: Lipid A mimetics as adjuvants and immunomodulators. Trends Microbiol. 2002, 10, S32–S37. [Google Scholar]
- Takeuchi, O.; Hoshino, K.; Kawai, T.; Sanjo, H.; Takada, H.; Ogawa, T.; Takeda, K.; Akira, S. Differential roles of TLR2 and TLR4 in recognition of Gram-negative and Gram-positive bacterial cell wall components. Immunity 1999, 11, 443–451. [Google Scholar]
- Yang, M.; Yan, Y.; Fang, M.; Wan, M.; Wu, X.; Zhang, X.; Zhao, T.; Wei, H.; Song, D.; Wang, L.; et al. MF59 formulated with CpG ODN as a potent adjuvant of recombinant hsp65-MUC1 for inducing anti-MUC1+ tumor immunity in mice. Int. Immunopharmacol. 2012, 13, 408–416. [Google Scholar]
- Lakshminarayanan, V.; Thompson, P.; Wolfert, M.A.; Buskas, T.; Bradley, J.M.; Pathangey, L.B.; Madsen, C.S.; Cohen, P.A.; Gendler, S.J.; Boons, G.J. Immune recognition of tumor-associated mucin MUC1 is achieved by a fully synthetic aberrantly glycosylated MUC1 tripartite vaccine. Proc. Natl. Acad. Sci. USA 2012, 109, 261–266. [Google Scholar]
- Abdel-Aal, A.B.; Lakshminarayanan, V.; Thompson, P.; Supekar, N.; Bradley, J.M.; Wolfert, M.A.; Cohen, P.A.; Gendler, S.J.; Boons, G.J. Immune and anticancer responses elicited by fully synthetic aberrantly glycosylated MUC1 tripartite vaccines modified by a TLR2 or TLR9 agonist. Chembiochem 2014, 15, 1508–1513. [Google Scholar]
- Thompson, P.; Lakshminarayanan, V.; Supekar, N.T.; Bradley, J.M.; Cohen, P.A.; Wolfert, M.A.; Gendler, S.J.; Boons, G.J. Linear synthesis and immunological properties of a fully synthetic vaccine candidate containing a sialylated MUC1 glycopeptide. Chem. Commun. (Camb.) 2015, 51, 10214–10217. [Google Scholar]
- Cai, H.; Degliangeli, F.; Palitzsch, B.; Gerlitzki, B.; Kunz, H.; Schmitt, E.; Fiammengo, R.; Westerlind, U. Glycopeptide-functionalized gold nanoparticles for antibody induction against the tumor associated mucin-1 glycoprotein. Bioorg. Med. Chem. 2016, 24, 1132–1135. [Google Scholar]
- Sarkar, S.; Lombardo, S.A.; Herner, D.N.; Talan, R.S.; Wall, K.A.; Sucheck, S.J. Synthesis of a single-molecule L-rhamnose-containing three-component vaccine and evaluation of antigenicity in the presence of anti-L-rhamnose antibodies. J. Am. Chem. Soc. 2010, 132, 17236–17246. [Google Scholar]
- Sarkar, S.; Salyer, A.C.; Wall, K.A.; Sucheck, S.J. Synthesis and immunological evaluation of a MUC1 glycopeptide incorporated into L-rhamnose displaying liposomes. Bioconjug. Chem. 2013, 24, 363–375. [Google Scholar]
- Karmakar, P.; Lee, K.; Sarkar, S.; Wall, K.A.; Sucheck, S.J. Synthesis of a liposomal MUC1 glycopeptide-based immunotherapeutic and evaluation of the effect of L-rhamnose targeting on cellular immune responses. Bioconjug. Chem. 2016, 27, 110–120. [Google Scholar]
- Hartmann, S.; Nuhn, L.; Palitzsch, B.; Glaffig, M.; Stergiou, N.; Gerlitzki, B.; Schmitt, E.; Kunz, H.; Zentel, R. CpG-loaded multifunctional cationic nanohydrogel particles as self-adjuvanting glycopeptide antitumor vaccines. Adv. Healthc. Mater. 2015, 4, 522–527. [Google Scholar]
- Nuhn, L.; Hartmann, S.; Palitzsch, B.; Gerlitzki, B.; Schmitt, E.; Zentel, R.; Kunz, H. Water-soluble polymers coupled with glycopeptide antigens and T-cell epitopes as potential antitumor vaccines. Angew. Chem. Int. Ed. Engl. 2013, 52, 10652–10656. [Google Scholar]
- Glaffig, M.; Palitzsch, B.; Stergiou, N.; Schull, C.; Strassburger, D.; Schmitt, E.; Frey, H.; Kunz, H. Enhanced immunogenicity of multivalent MUC1 glycopeptide antitumour vaccines based on hyperbranched polymers. Org. Biomol. Chem. 2015, 13, 10150–10154. [Google Scholar]
- Huang, Z.H.; Shi, L.; Ma, J.W.; Sun, Z.Y.; Cai, H.; Chen, Y.X.; Zhao, Y.F.; Li, Y.M. A totally synthetic, self-assembling, adjuvant-free MUC1 glycopeptide vaccine for cancer therapy. J. Am. Chem. Soc. 2012, 134, 8730–8733. [Google Scholar]
- Mond, J.J.; Lees, A.; Snapper, C.M. T cell-independent antigens type 2. Annu. Rev. Immunol. 1995, 13, 655–692. [Google Scholar]
- Liu, Y.F.; Sun, Z.Y.; Chen, P.G.; Huang, Z.H.; Gao, Y.; Shi, L.; Zhao, Y.F.; Chen, Y.X.; Li, Y.M. Glycopeptide nanoconjugates based on multilayer self-assembly as an antitumor vaccine. Bioconjug. Chem. 2015, 26, 1439–1442. [Google Scholar]
- Pillai, K.; Pourgholami, M.H.; Chua, T.C.; Morris, D.L. MUC1 as a potential target in anticancer therapies. Am. J. Clin. Oncol. 2015, 38, 108–118. [Google Scholar]
- Weng, Y.; Shao, L.; Ouyang, H.; Liu, Y.; Yao, J.; Yang, H.; Luo, Y.; Wang, H.; Zhao, Z.; Mou, H.; et al. A unique MUC1–2-VNTR DNA vaccine suppresses tumor growth and prolongs survival in a murine multiple myeloma model. Oncol. Rep. 2012, 27, 1815–1822. [Google Scholar]
- Richichi, B.; Thomas, B.; Fiore, M.; Bosco, R.; Qureshi, H.; Nativi, C.; Renaudet, O.; BenMohamed, L. A cancer therapeutic vaccine based on clustered Tn-antigen mimetics induces strong antibody-mediated protective immunity. Angew. Chem. Int. Ed. Engl. 2014, 53, 11917–11920. [Google Scholar]
- Nativi, C.; Renaudet, O. Recent progress in antitumoral synthetic vaccines. ACS Med. Chem. Lett. 2014, 5, 1176–1178. [Google Scholar]
- Yang, F.; Zheng, X.J.; Huo, C.X.; Wang, Y.; Zhang, Y.; Ye, X.S. Enhancement of the immunogenicity of synthetic carbohydrate vaccines by chemical modifications of STn antigen. ACS Chem. Biol. 2011, 6, 252–259. [Google Scholar]
- Yin, Z.; Huang, X. Recent development in carbohydrate based anti-cancer vaccines. J. Carbohydr. Chem. 2012, 31, 143–186. [Google Scholar]
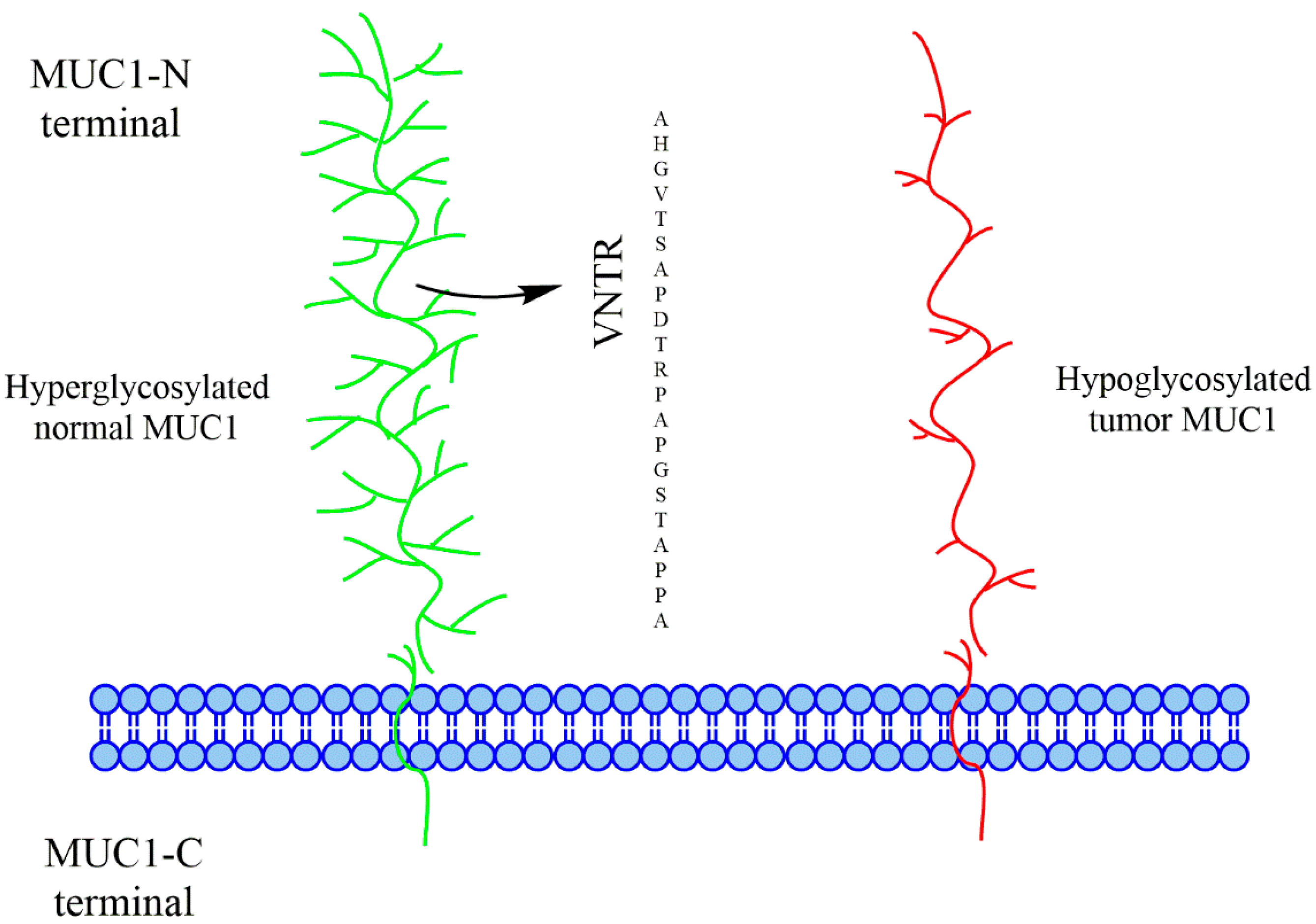


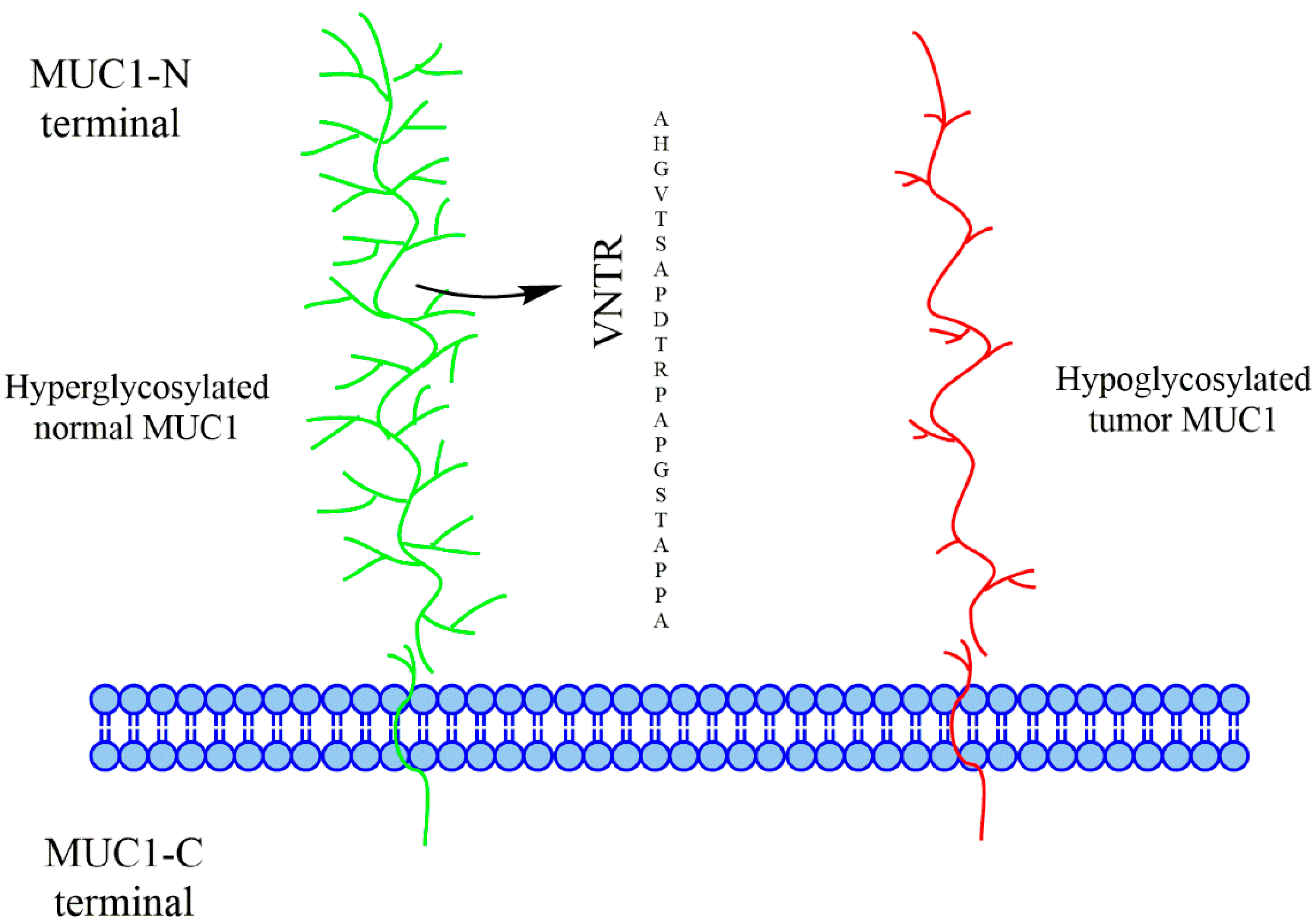{kind=link}
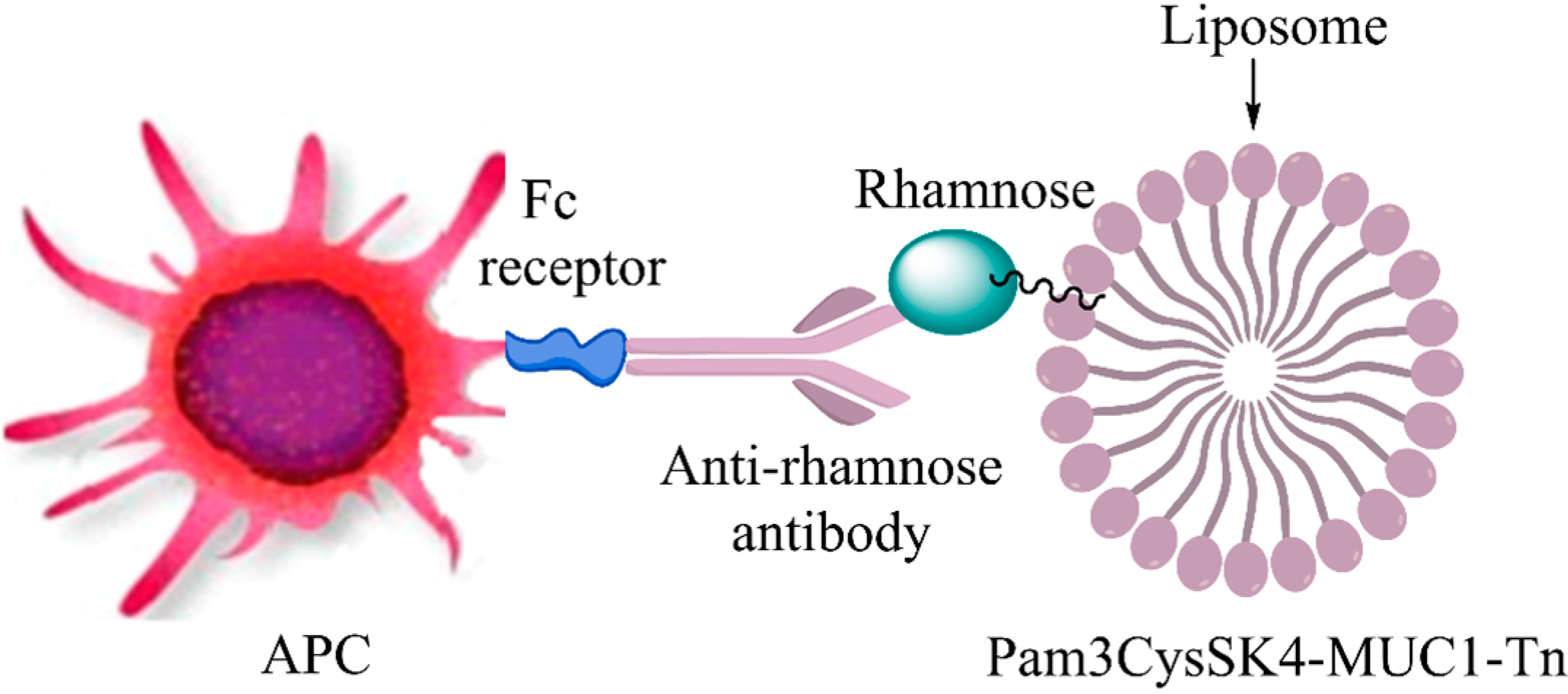{kind=link}
| Authors | Year | Vaccine Preparations | Cell Lines & Animals | Response |
|---|---|---|---|---|
| Karmakar et al. [73] | 2016 | Rhamnose containg liposomal Pam3Cys-MUC1-Tn | EL4, C57BL/6 | CTL, IFN-γ |
| Lakshminarayanan et al. [21] | 2016 | Glycosylates, no-glycosylates or tumor derived MUC1 vaccines | C57mg, MC38, B16, EL4, Panc02, MUC1 Tg mice (C57BL/6) | IFN-γ producing CD4+ & CD8+ cells |
| Cai et al. [70] | 2016 | Glycopeptide-functionalized gold nanoparticles | MCF7, BALB/c | IgG1, IgG2a, IgG2b |
| Lu et al. [43] | 2015 | MUC1 peptide on cholera toxin B (CTB) subunit | B16, C57BL/6 | CTL, Th1 |
| Glaffig et al. [76] | 2015 | MUC1-P2 conjugate on hyperbranched polymers | T47D, BALB/c | IgG1, IgG2a, IgG2b, IgM |
| Thompson et al. [69] | 2015 | MUC1-STn glycopeptide with T cell epitope & TLR2 ligand | C57mg, B16, MUC1 Tg mice (C57BL/6) | CTL, IFN-γ, IgG1, IgG2a, IgG2b, IgG3 |
| Hartmann et al. [74] | 2015 | CpG loaded nanohydrogel particles | T47D, BALB/c | Upregulation of CD40, CD80, CD86 |
| Johannes et al. [49] | 2015 | Fluorinated MUC1-BSA/TTox conjugate vaccine | MCF7, BALB/cj | IgG1, IgG2a, IgG2b but no IgM |
| Wang et al. [42] | 2014 | MUC1 VNTR and (MUC1-VPP) with DDA/MPL as adjuvant | B16, C2C12, C57BL/6 | CTL, IFN-γ |
| McDonald et al. [60] | 2014 | MUC1-MALP2 conjugate vaccine | C57BL/6 | IgM, IgG1, IgG2b, IgG3 |
| Cai et al. [58] | 2014 | Multivalent glycopeptide-lipopeptide vaccine | MCF7, BALB/c | IgG, IgM |
| Palitzsch et al. [24] | 2014 | MUC1 glycopeptide with three different Th cell epitopes | T47D, BALB/c | IgG, IgM |
| Abdel-Aal et al. [68] | 2014 | MUC1 tripartite vaccine modified by TLR 2 or TLR 4 | C57mg, EL4, MUC1 Tg mice (C57BL/6) | IgG, IgM, CTL |
| Geraci et al. [59] | 2013 | PDTRP MUC1 conjugated calixarene containing TLR2 ligand | MCF7, BALB/c | IgG |
| Sarkar et al. [72] | 2013 | MUC1 glycopeptide into L-Rhamnose displaying liposomes | BALB/c | IgG1, IgG2a, IgG2b, IgM |
| Cai et al. [47] | 2013 | MUC1 glycopeptide with T cell epitope from TTox | MCF7, T47D, | IgG1, IgG2a, IgG2b, IgG3, IgM |
| Nuhn et al. [75] | 2013 | MUC1-VNTR with T helper epitope and P(HPMA) | MCF7, BALB/c | IgG |
| Yang et al. [66] | 2012 | Recombinant HSP65-MUC1 and MF59-YW002 as adjuvant | B16, C57BL/6 | Th1, CTL |
| Huang et al. [77] | 2012 | MUC1 glycopeptide with a B cell epitope | MCF7 | IgG1, IgG2a, IgG2b, IgG3, IgM |
| Cai et al. [36] | 2012 | MUC1 Glycopeptide-BSA vaccine | MCF7, BALB/c | IgG1, IgG2a, IgG2b, IgG3, IgA, IgM |
| Weng et al. [81] | 2012 | MUC1-2-VNTR DNA vaccine | BALB/c | CTL |
| Lakshminarayanan et al. [67] | 2012 | MUC1 tripartite vaccine | Yac.MUC1, C57mg, MUC1 Tg mice (C57BL/6) | IgG, CTL, TNF-α, RANTES, IL-6, 12, IL-1β |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hossain, M.K.; Wall, K.A. Immunological Evaluation of Recent MUC1 Glycopeptide Cancer Vaccines. Vaccines 2016, 4, 25. https://doi.org/10.3390/vaccines4030025
Hossain MK, Wall KA. Immunological Evaluation of Recent MUC1 Glycopeptide Cancer Vaccines. Vaccines. 2016; 4(3):25. https://doi.org/10.3390/vaccines4030025
Chicago/Turabian StyleHossain, Md Kamal, and Katherine A. Wall. 2016. "Immunological Evaluation of Recent MUC1 Glycopeptide Cancer Vaccines" Vaccines 4, no. 3: 25. https://doi.org/10.3390/vaccines4030025
APA StyleHossain, M. K., & Wall, K. A. (2016). Immunological Evaluation of Recent MUC1 Glycopeptide Cancer Vaccines. Vaccines, 4(3), 25. https://doi.org/10.3390/vaccines4030025