
Recent Advances in Subunit Vaccine Carriers


Abstract
:
1. Introduction
2. Types of Vaccines
3. Immunology of Vaccines
4. Minimal Subunit Vaccine Development
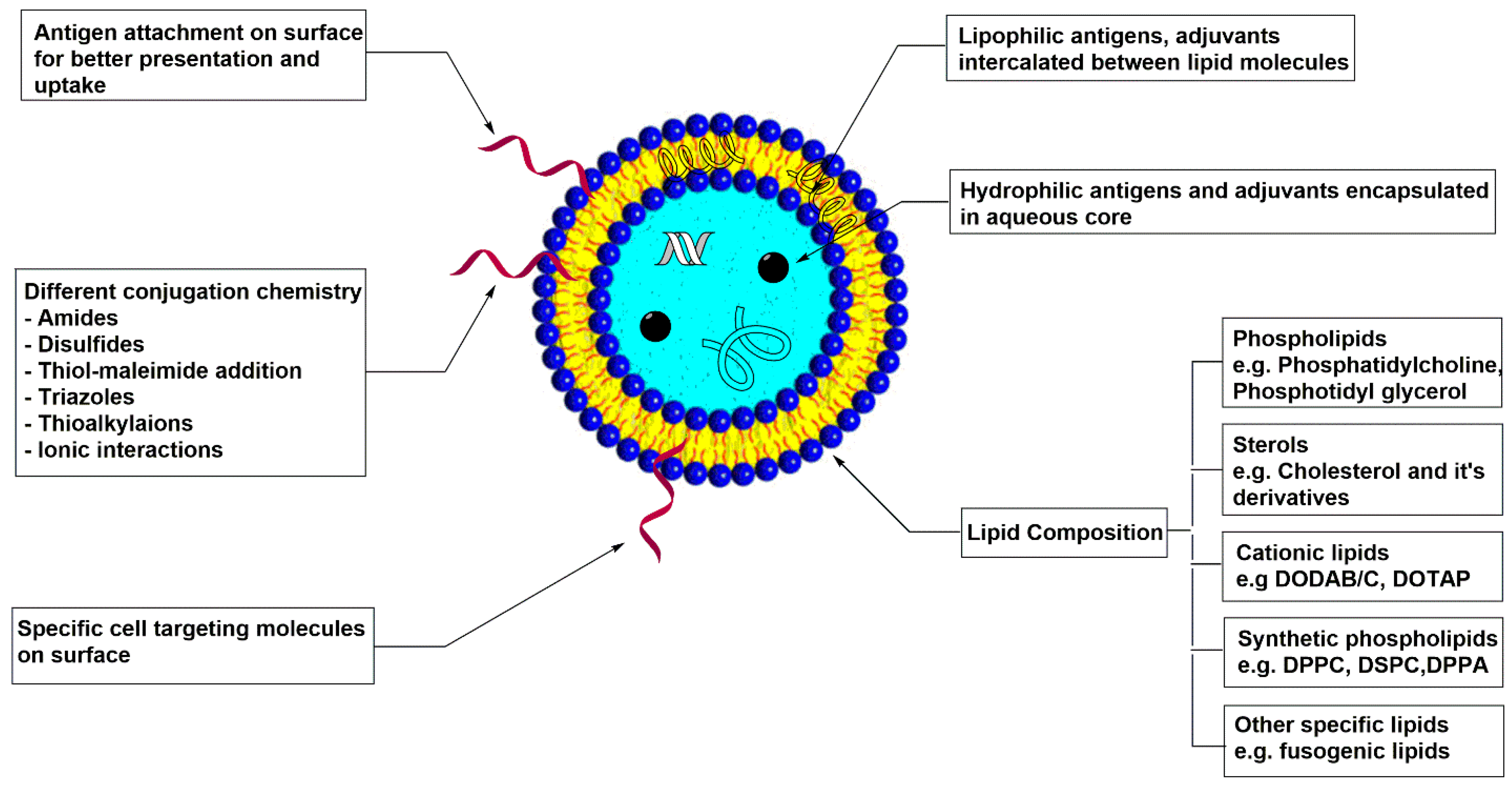
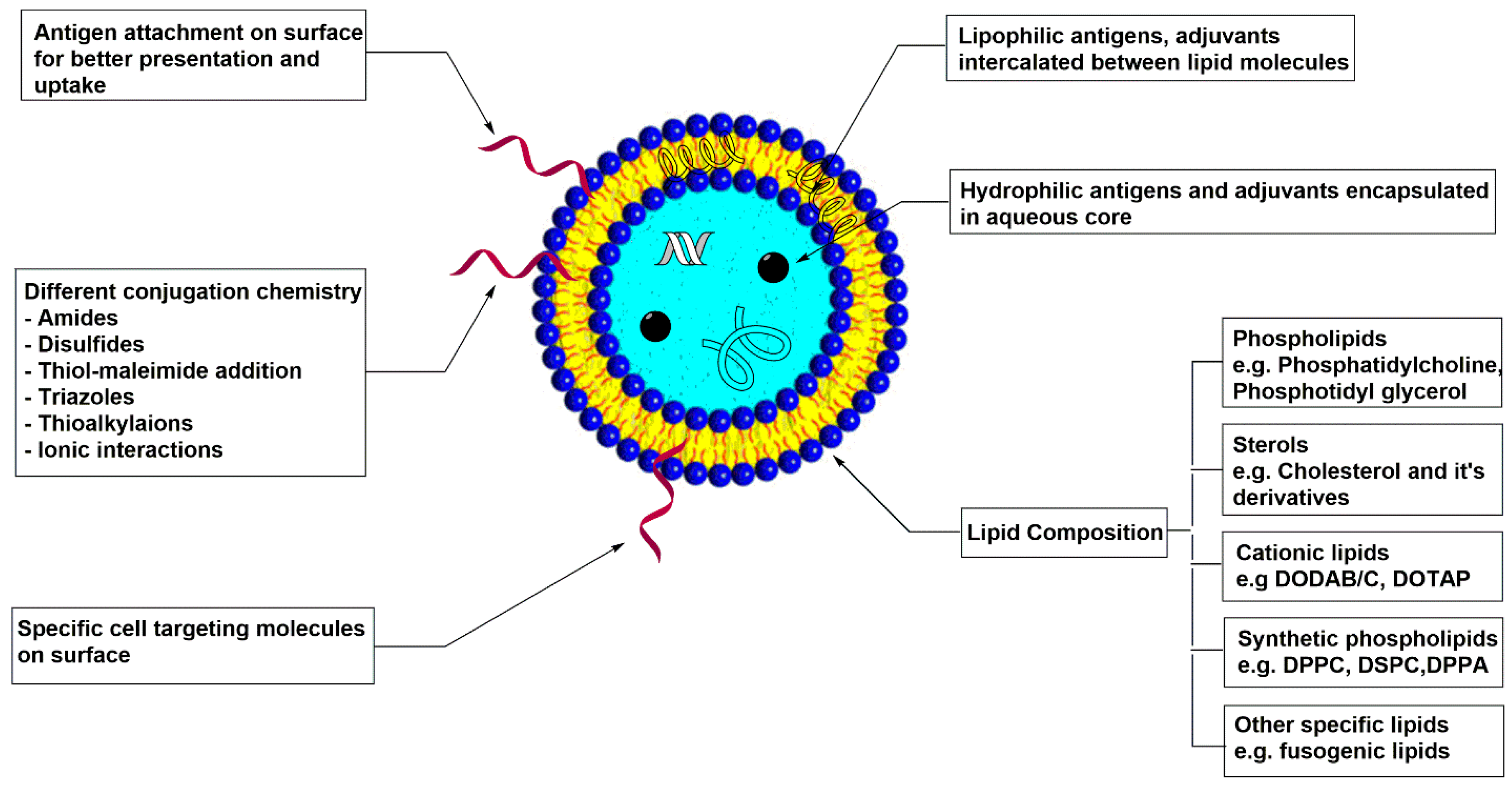
5. Liposomes
5.1. Physiochemical Properties
5.2. Liposome-Polymer Hybrid Vaccine Carrier
5.3. Dendritic Cells (DCs) Targeting Liposomal Vaccine Carrier
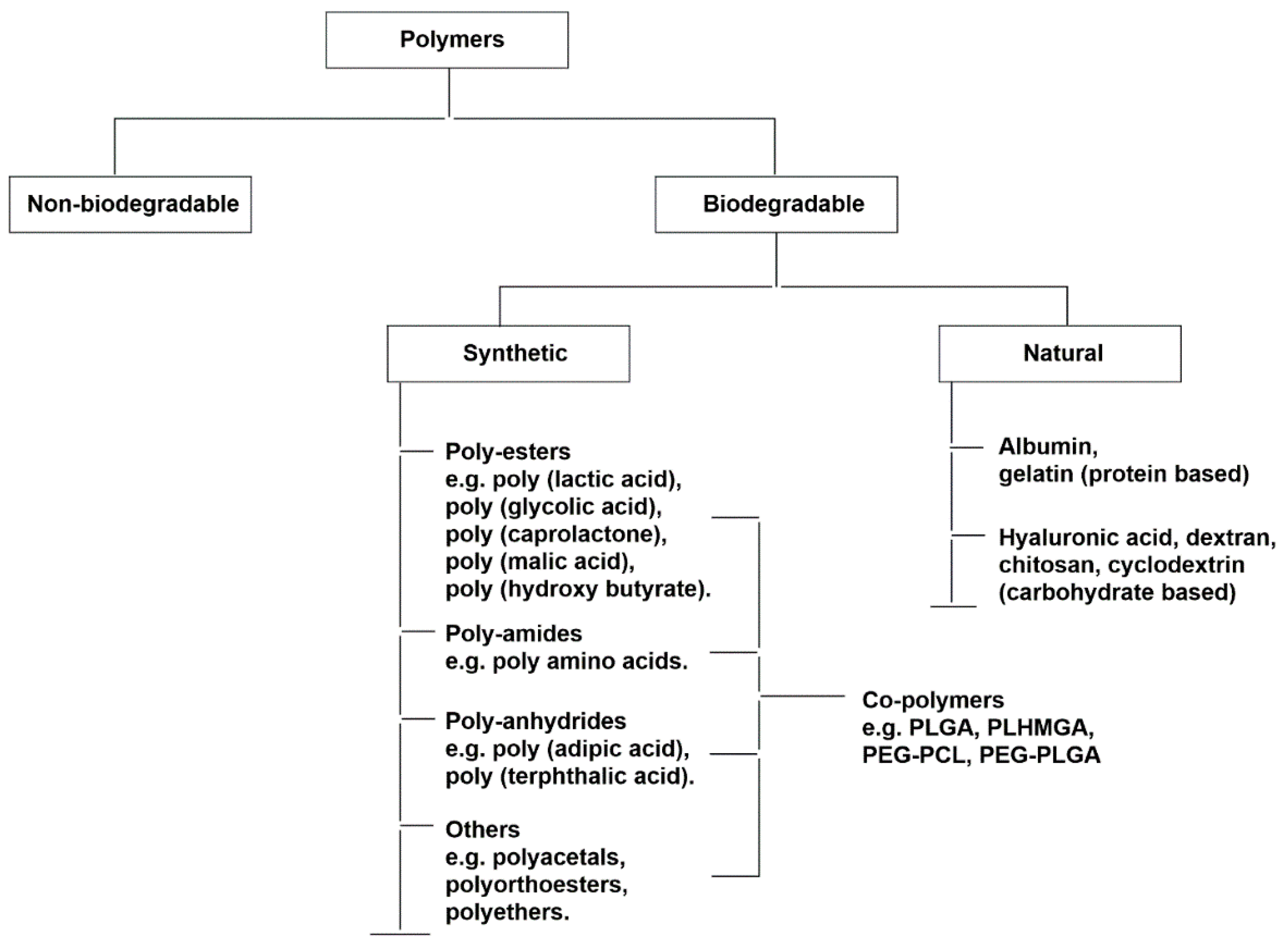
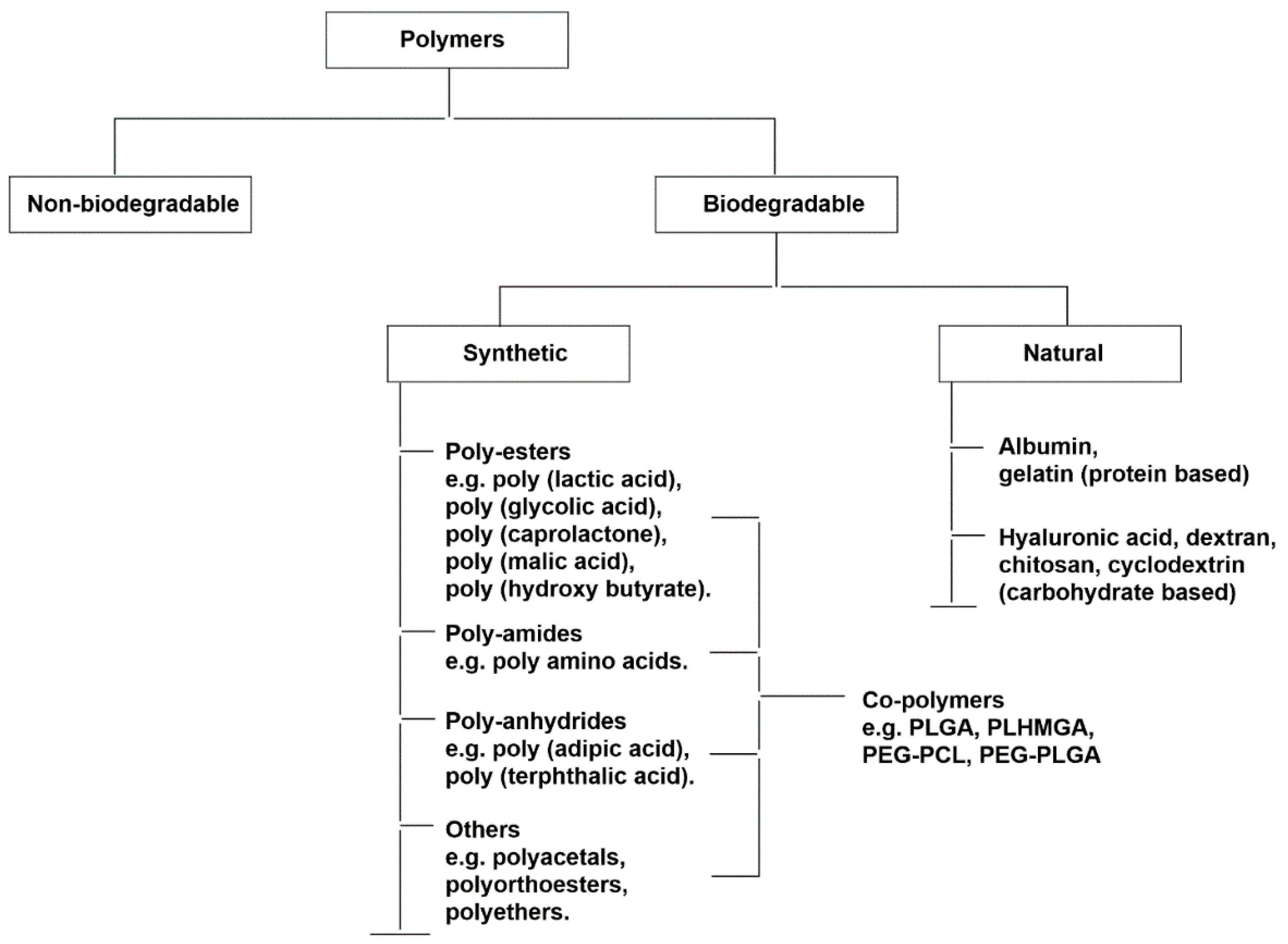
6. Polymeric Nanoparticles
Physiochemical Properties
7. Inorganic Nanoparticles
8. Conclusions and Future Directions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ALF | Army Liposome Formulation |
| CD 80 | Cluster of Differentiation 80 |
| CFA | Complete Freund’s adjuvant |
| DDA | Dimethyl dioctadecyl-ammonium bromide |
| DLPE | 1,2-dilauroyl-sn-glycero-3-phosphoethanolamine |
| DMG | 1,2-Dimyristoyl-sn-glycerol |
| DMPC | Dimyristoyl phosphatidylcholine |
| DMPG | Dimyristoyl phosphatidylglycerol |
| DOPC | 1,2-dioleoyl-sn-glycero-3-phosphocholine |
| DOPE | 1,2-dioleoyl-sn-glycero-3-phosphoethanolamine |
| DOPG | 1,2-dioleoyl-sn-glycero-3-phoshpo-(1’-rac-gylcerol) |
| DOTAP | 1,2-dioleoyl-3-trimethylammonium propane |
| DPPC | Dipalmitoyl phosphatidylcholine |
| DSPE-PEG | 1,2-distearoyl-sn-glycero-3-phosphoethanolamine-N-[methoxy (polyethyleneglycol)] |
| EPC | Egg phosphatidylcholine |
| HBsAg | Recombinant human hepatitis B virus surface antigen |
| IFA | Incomplete Freund’s adjuvant |
| INF-γ | Interferon γ |
| IVTT | in vitro transcription and translation |
| MART-1 | Melanoma antigen recognized by T-cells 1 |
| MHC | Major Histocompatibility Complex |
| MPC | Mannose-PEG1000-cholesterol |
| MPER | Membrane proximal external region |
| MPLA | Monophosphoryl lipid A |
| NK cells | Natural Killer cells |
| PC | Phosphatidylcholine |
| PEG | Polyethylene Glycol |
| POPE | 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphoethanolamine |
| POPG | 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphatidylglycerol |
| SA | Stearyl amine |
| S1P | Sphingosine-1-phosphate |
| SPC | Soy phosphatidylcholine |
| TDB | d-(+)-trehalose 6,6-dibehenate |
| VMP | Vivax malaria protein |
References
- Ragupathi, G.; Damani, P.; Srivastava, G.; Srivastava, O.; Sucheck, S.J.; Ichikawa, Y.; Livingston, P.O. Synthesis of sialyl Lewisa (sLea, CA19-9) and construction of an immunogenic sLea vaccine. Cancer Immunol. Immunother. 2009, 58, 1397–1405. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Warren, J.D.; Danishefsky, S.J. Synthetic carbohydrate-based anticancer vaccines: The memorial sloan-kettering experience. Expert Rev. Vaccines 2009, 8, 1399–1413. [Google Scholar] [CrossRef] [PubMed]
- da Costa, C.; Walker, B.; Bonavia, A. Tuberculosis vaccines—State of the art, and novel approaches to vaccine development. Int. J. Infect. Dis. 2015, 32, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, H.A. Profiling the host immune response to tuberculosis vaccines. Vaccine 2015, 33, 5313–5315. [Google Scholar] [CrossRef] [PubMed]
- Tam, P.H.; Lowary, T.L. Mycobacterial lipoarabinomannan fragments as haptens for potential anti-tuberculosis vaccines. In Carbohydrate Chemistry: Chemical and Biological Approaches; Royal Society of Chemistry: London, UK, 2010; pp. 38–63. [Google Scholar]
- Baxter, D. Active and passive immunity, vaccine types, excipients and licensing. Occup. Med. 2007, 57, 552–556. [Google Scholar] [CrossRef] [PubMed]
- Scott, C. Classifying vaccines: From cowpox to the cutting edge. BioProcess Int. 2004, 2, 14–23. [Google Scholar]
- Datta, J.; Terhune, J.H.; Lowenfeld, L.; Cintolo, J.A.; Xu, S.; Roses, R.E.; Czerniecki, B.J. Optimizing dendritic cell-based approaches for cancer immunotherapy. Yale J. Biol. Med. 2014, 87, 491–518. [Google Scholar] [PubMed]
- Amigorena, S.; Savina, A. Intracellular mechanisms of antigen cross presentation in dendritic cells. Curr. Opin. Immunol. 2010, 22, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Alexopoulou, L.; Holt, A.C.; Medzhitov, R.; Flavell, R.A. Recognition of double-stranded RNA and activation of NF-κB by Toll-like receptor 3. Nature 2001, 413, 732–738. [Google Scholar] [CrossRef] [PubMed]
- Janeway, C.A., Jr.; Medzhitov, R. Innate immune recognition. Annu. Rev. Immunol. 2002, 20, 197–216. [Google Scholar] [CrossRef] [PubMed]
- Danishefsky, S.J.; Allen, J.R. From the laboratory to the clinic: A retrospective on fully synthetic carbohydrate-based anticancer vaccines. Angew. Chem. Int. Ed. 2000, 39, 836–863. [Google Scholar] [CrossRef]
- Ouerfelli, O.; Warren, J.D.; Wilson, R.M.; Danishefsky, S.J. Synthetic carbohydrate-based antitumor vaccines: Challenges and opportunities. Expert Rev. Vaccines 2005, 4, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Glunz, P.W.; Hintermann, S.; Schwarz, J.B.; Kuduk, S.D.; Chen, X.T.; Williams, L.J.; Sames, D.; Danishefsky, S.J.; Kudryashov, V.; Lloyd, K.O. Probing cell surface “glyco-architecture” through total synthesis. Immunological consequences of a human blood group determinant in a clustered mucin-like context. J. Am. Chem. Soc. 1999, 121, 10636–10637. [Google Scholar] [CrossRef]
- Glunz, P.W.; Hintermann, S.; Williams, L.J.; Schwarz, J.B.; Kuduk, S.D.; Kudryashov, V.; Lloyd, K.O.; Danishefsky, S.J. Design and synthesis of Ley-bearing glycopeptides that mimic cell surface Ley mucin glycoprotein architecture. J. Am. Chem. Soc. 2000, 122, 7273–7279. [Google Scholar] [CrossRef]
- Kudryashov, V.; Glunz, P.W.; Williams, L.J.; Hintermann, S.; Danishefsky, S.J.; Lloyd, K.O. Toward optimized carbohydrate-based anticancer vaccines: Epitope clustering, carrier structure, and adjuvant all influence antibody responses to Lewisy conjugates in mice. Proc. Natl. Acad. Sci. USA 2001, 98, 3264–3269. [Google Scholar] [CrossRef] [PubMed]
- Toyokuni, T.; Dean, B.; Cai, S.; Boivin, D.; Hakomori, S.; Singhal, A.K. Synthetic vaccines: Synthesis of a dimeric Tn antigen-lipopeptide conjugate that elicits immune responses against Tn-expressing glycoproteins. J. Am. Chem. Soc. 1994, 116, 395–396. [Google Scholar] [CrossRef]
- Toyokuni, T.; Hakomori, S.I.; Singhal, A.K. Synthetic carbohydrate vaccines: Synthesis and immunogenicity of Tn antigen conjugates. Bioorg. Med. Chem. 1994, 2, 1119–1132. [Google Scholar] [CrossRef]
- Buskas, T.; Ingale, S.; Boons, G.J. Towards a fully synthetic carbohydrate-based anticancer vaccine: Synthesis and immunological evaluation of a lipidated glycopeptide containing the tumor-associated Tn antigen. Angew. Chem. Int. Ed. 2005, 44, 5985–5988. [Google Scholar] [CrossRef] [PubMed]
- Krikorian, D.; Panou-Pomonis, E.; Voitharou, C.; Sakarellos, C.; Sakarellos-Daitsiotis, M. A peptide carrier with a built-in vaccine adjuvant: Construction of immunogenic conjugates. Bioconjugate Chem. 2005, 16, 812–819. [Google Scholar] [CrossRef] [PubMed]
- Brocke, C.; Kunz, H. Synthesis of tumor-associated glycopeptide antigens. Bioorg. Med. Chem. 2002, 10, 3085–3112. [Google Scholar] [CrossRef]
- Buskas, T.; Thompson, P.; Boons, G.J. Immunotherapy for cancer: Synthetic carbohydrate-based vaccines. Chem. Commun. 2009, 36, 5335–5349. [Google Scholar] [CrossRef] [PubMed]
- Hanisch, F.G. Immunology of MUC1 and Advancements in the Development of MUC1 Glycopeptide Tumor Vaccines: An Update. Available online: http://www.eurekaselect.com/117190/chapter/immunology-of-muc1-and-advancements-in-the-development-of-muc1-glycopeptide-tumor-vaccines%3A-an-updat (accessed on 7 November 2015).
- Heimburg-Molinaro, J.; Lum, M.; Vijay, G.; Jain, M.; Almogren, A.; Rittenhouse-Olson, K. Cancer vaccines and carbohydrate epitopes. Vaccine 2011, 29, 8802–8826. [Google Scholar] [CrossRef] [PubMed]
- Morelli, L.; Poletti, L.; Lay, L. Carbohydrates and immunology: Synthetic oligosaccharide antigens for vaccine formulation. Eur. J. Org. Chem. 2011, 29, 5723–5777. [Google Scholar] [CrossRef]
- Wang, L.X. Synthetic carbohydrate antigens for HIV vaccine design. Curr. Opin. Chem. Biol. 2013, 17, 997–1005. [Google Scholar] [CrossRef] [PubMed]
- Bernardi, A.; Jiménez-Barbero, J.; Casnati, A.; De Castro, C.; Darbre, T.; Fieschi, F.; Finne, J.; Funken, H.; Jaeger, K.E.; Lahmann, M.; et al. Multivalent glycoconjugates as anti-pathogenic agents. Chem. Soc. Rev. 2013, 42, 4709–4727. [Google Scholar] [CrossRef] [PubMed]
- Azuma, I.; Seya, T. Development of immunoadjuvants for immunotherapy of cancer. Int. Immunopharmacol. 2001, 1, 1249–1259. [Google Scholar] [CrossRef]
- Dredge, K.; Marriott, J.B.; Todryk, S.M.; Dalgleish, A.G. Adjuvants and the promotion of Th1-type cytokines in tumour immunotherapy. Cancer Immunol. Immunother. 2002, 51, 521–531. [Google Scholar] [CrossRef] [PubMed]
- Tsan, M.F. Toll-like receptors, inflammation and cancer. Semin. Cancer Biol. 2006, 16, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Vogel, F.R. Immunologic adjuvants for modern vaccine formulations. Ann. N. Y. Acad. Sci. 1995, 754, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Gregory, A.E.; Titball, R.; Williamson, D. Vaccine delivery using nanoparticles. Front. Cell. Infect. Microbiol. 2013. [Google Scholar] [CrossRef] [PubMed]
- Krishnamachari, Y.; Geary, S.M.; Lemke, C.D.; Salem, A.K. Nanoparticle delivery systems in cancer vaccines. Pharmaceut. Res. 2011, 28, 215–236. [Google Scholar] [CrossRef] [PubMed]
- Sahdev, P.; Ochyl, L.J.; Moon, J.J. Biomaterials for nanoparticle vaccine delivery systems. Pharmaceut. Res. 2014, 31, 2563–2582. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Seth, A.; Wibowo, N.; Zhao, C.X.; Mitter, N.; Yu, C.; Middelberg, A.P. Nanoparticle vaccines. Vaccine 2014, 32, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Henriksen-Lacey, M.; Korsholm, K.S.; Andersen, P.; Perrie, Y.; Christensen, D. Liposomal vaccine delivery systems. Expert Opin. Drug Deliv. 2011, 8, 505–519. [Google Scholar] [CrossRef] [PubMed]
- Perrie, Y.; Crofts, F.; Devitt, A.; Griffiths, H.R.; Kastner, E.; Nadella, V. Designing liposomal adjuvants for the next generation of vaccines. Adv. Drug Deliv. Rev. 2016, 99, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Schwendener, R.A. Liposomes as vaccine delivery systems: A review of the recent advances. Ther. Adv. Vaccines 2014, 2, 159–182. [Google Scholar] [CrossRef] [PubMed]
- Shashi, K.; Satinder, K.; Bharat, P. A complete review on: Liposomes. Int. J. Pharm. 2012, 3, 10–16. [Google Scholar]
- Watson, D.S.; Endsley, A.N.; Huang, L. Design considerations for liposomal vaccines: Influence of formulation parameters on antibody and cell-mediated immune responses to liposome associated antigens. Vaccine 2012, 30, 2256–2272. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Sharma, U.S. Liposomes in drug delivery: Progress and limitations. Int. J. Pharm. 1997, 154, 123–140. [Google Scholar] [CrossRef]
- Shahum, E.; Thérien, H.M. Correlation between in vitro and in vivo behaviour of liposomal antigens. Vaccine 1994, 12, 1125–1131. [Google Scholar] [CrossRef]
- Tan, L.; Weisslg, V.; Gregorladls, G. Comparison of the immune response against polio peptides covalently-surface-linked to and internally-entrapped in liposomes. Asian Pac. J. Allergy Immunol. 2011, 9, 25–30. [Google Scholar]
- Thérien, H.M.; Lair, D.; Shahum, E. Liposomal vaccine: Influence of antigen association on the kinetics of the humoral response. Vaccine 1990, 8, 558–562. [Google Scholar] [CrossRef]
- Vannier, W.; Snyder, S. Antibody responses to liposome-associated antigen. Immunol. Lett. 1988, 19, 59–64. [Google Scholar] [CrossRef]
- White, W.I.; Cassatt, D.R.; Madsen, J.; Burke, S.J.; Woods, R.M.; Wassef, N.M.; Alving, C.R.; Koenig, S. Antibody and cytotoxic T-lymphocyte responses to a single liposome-associated peptide antigen. Vaccine 1995, 13, 1111–1122. [Google Scholar] [CrossRef]
- Conwell, C.C.; Huang, L. Recent advances in non-viral gene delivery. Adv. Genet. 2005, 53, 1–18. [Google Scholar]
- Gao, X.; Huang, L. Cationic liposome-mediated gene transfer. Gene Ther. 1995, 2, 710–722. [Google Scholar] [PubMed]
- Li, W.; Szoka, F., Jr. Lipid-based nanoparticles for nucleic acid delivery. Pharm. Res. 2007, 24, 438–449. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Szoka, F.C. Mechanism of DNA release from cationic liposome/DNA complexes used in cell transfection. Biochemistry 1996, 35, 5616–5623. [Google Scholar] [CrossRef] [PubMed]
- Akbarzadeh, A.; Rezaei-Sadabady, R.; Davaran, S.; Joo, S.W.; Zarghami, N.; Hanifehpour, Y.; Samiei, M.; Kouhi, M.; Nejati-Koshki, K. Liposome: Classification, preparation, and applications. Nanoscale Res. Lett. 2013. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Li, X.; Zhang, T.; Song, Y.; She, Z.; Li, J.; Deng, Y. Progress involving new techniques for liposome preparation. Asian J. Pharm. Sci. 2014, 9, 176–182. [Google Scholar] [CrossRef]
- Patil, Y.P.; Jadhav, S. Novel methods for liposome preparation. Chem. Phys. Lipids 2014, 177, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Szoka, F., Jr.; Papahadjopoulos, D. Comparative properties and methods of preparation of lipid vesicles (liposomes). Annu. Rev. Biophys. Bioeng. 1980, 9, 467–508. [Google Scholar] [CrossRef] [PubMed]
- Manolova, V.; Flace, A.; Bauer, M.; Schwarz, K.; Saudan, P.; Bachmann, M.F. Nanoparticles target distinct dendritic cell populations according to their size. Eur. J. Immunol. 2008, 38, 1404–1413. [Google Scholar] [CrossRef] [PubMed]
- Mann, J.F.; Shakir, E.; Carter, K.C.; Mullen, A.B.; Alexander, J.; Ferro, V.A. Lipid vesicle size of an oral influenza vaccine delivery vehicle influences the Th1/Th2 bias in the immune response and protection against infection. Vaccine 2009, 27, 3643–3649. [Google Scholar] [CrossRef] [PubMed]
- Miyabe, H.; Hyodo, M.; Nakamura, T.; Sato, Y.; Hayakawa, Y.; Harashima, H. A new adjuvant delivery system ‘cyclic di-GMP/YSK05 liposome’ for cancer immunotherapy. J. Control. Release 2014, 184, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Badiee, A.; Jaafari, M.R.; Khamesipour, A.; Samiei, A.; Soroush, D.; Kheiri, M.T.; Barkhordari, F.; McMaster, W.R.; Mahboudi, F. Enhancement of immune response and protection in BALB/c mice immunized with liposomal recombinant major surface glycoprotein of Leishmania (rgp63): The role of bilayer composition. Colloids Surf. B. Biointerfaces 2009, 74, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Garnier, F.; Forquet, F.; Bertolino, P.; Gerlier, D. Enhancement of in vivo and in vitro T cell response against measles virus haemagglutinin after its incorporation into liposomes: Effect of the phospholipid composition. Vaccine 1991, 9, 340–345. [Google Scholar] [CrossRef]
- Kahl, L.; Scott, C.A.; Lelchuk, R.; Gregoriadis, G.; Liew, F.Y. Vaccination against murine cutaneous leishmaniasis by using Leishmania major antigen/liposomes. Optimization and assessment of the requirement for intravenous immunization. J. Immunol. 1989, 142, 4441–4449. [Google Scholar] [PubMed]
- Kersten, G.; van de Put, A.M.; Teerlink, T.; Beuvery, E.C.; Crommelin, D.J. Immunogenicity of liposomes and iscoms containing the major outer membrane protein of Neisseria gonorrhoeae: Influence of protein content and liposomal bilayer composition. Infect Immun. 1988, 56, 1661–1664. [Google Scholar] [PubMed]
- Mazumdar, T.; Anam, K.; Ali, N. Influence of phospholipid composition on the adjuvanticity and protective efficacy of liposome-encapsulated Leishmania donovani antigens. J. Parasitol. 2005, 91, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Christensen, D.; Korsholm, K.S.; Andersen, P.; Agger, E.M. Cationic liposomes as vaccine adjuvants. Expert Rev. Vaccines 2011, 10, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Soema, P.C.; Willems, G.J.; Jiskoot, W.; Amorij, J.P.; Kersten, G.F. Predicting the influence of liposomal lipid composition on liposome size, zeta potential and liposome-induced dendritic cell maturation using a design of experiments approach. Eur. J. Pharm. Biopharm. 2015, 94, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Hafez, I.; Maurer, N.; Cullis, P. On the mechanism whereby cationic lipids promote intracellular delivery of polynucleic acids. Gene Ther. 2001, 8, 1188–1196. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Sahdev, P.; Ochyl, L.J.; Akerberg, J.J.; Moon, J.J. Cationic liposome–hyaluronic acid hybrid nanoparticles for intranasal vaccination with subunit antigens. J. Control. Release 2015, 208, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Karmakar, P.; Lee, K.; Sarkar, S.; Wall, K.A.; Sucheck, S.J. Synthesis of a liposomal MUC1 glycopeptide-based immunotherapeutic and evaluation of the effect of l-Rhamnose targeting on cellular immune responses. Bioconjugate Chem. 2016, 27, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Salyer, A.C.; Wall, K.A.; Sucheck, S.J. Synthesis and immunological evaluation of a MUC1 glycopeptide incorporated into l-rhamnose displaying liposomes. Bioconjugate Chem. 2013, 24, 363–375. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Lombardo, S.A.; Herner, D.N.; Talan, R.S.; Wall, K.A.; Sucheck, S.J. Synthesis of a single-molecule L-rhamnose-containing three-component vaccine and evaluation of antigenicity in the presence of anti-L-rhamnose antibodies. J. Am. Chem. Soc. 2010, 132, 17236–17246. [Google Scholar] [CrossRef] [PubMed]
- Oyelaran, O.; McShane, L.M.; Dodd, L.; Gildersleeve, J.C. Profiling human serum antibodies with a carbohydrate antigen microarray. J. Proteome Res. 2009, 8, 4301–4310. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.L.; Lin, H.J.; Wang, H.W.; Tsai, W.Y.; Lin, S.F.; Chien, M.Y.; Liang, P.H.; Huang, Y.Y.; Liu, D.Z. Galactosylated liposome as a dendritic cell-targeted mucosal vaccine for inducing protective anti-tumor immunity. Acta Biomater. 2015, 11, 356–367. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Zhen, Y.; Ma, X.; Wei, B.; Li, S.; Wang, N. Mannosylated and lipid A-incorporating cationic liposomes constituting microneedle arrays as an effective oral mucosal HBV vaccine applicable in the controlled temperature chain. Colloids Surf. B Biointerfaces 2015, 126, 520–530. [Google Scholar] [CrossRef] [PubMed]
- Derrick, S.C.; Yang, A.; Parra, M.; Kolibab, K.; Morris, S.L. Effect of cationic liposomes on BCG trafficking and vaccine-induced immune responses following a subcutaneous immunization in mice. Vaccine 2015, 33, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, K.F.; Amir, M.; Khan, N.; Rama Krishna, G.; Sheikh, J.A.; Rajagopal, K.; Agrewala, J.N. Prime-boost vaccination strategy with bacillus Calmette-Guerin (BCG) and liposomized alpha-crystalline protein 1 reinvigorates BCG potency. Clin. Exp. Immunol. 2015, 181, 286–296. [Google Scholar] [CrossRef] [PubMed]
- Beck, Z.; Matyas, G.R.; Jalah, R.; Rao, M.; Polonis, V.R.; Alving, C.R. Differential immune responses to HIV-1 envelope protein induced by liposomal adjuvant formulations containing monophosphoryl lipid A with or without QS21. Vaccine 2015, 33, 5578–5587. [Google Scholar] [CrossRef] [PubMed]
- Hanson, M.C.; Abraham, W.; Crespo, M.P.; Chen, S.H.; Liu, H.; Szeto, G.L.; Kim, M.; Reinherz, E.L.; Irvine, D.J. Liposomal vaccines incorporating molecular adjuvants and intrastructural T-cell help promote the immunogenicity of HIV membrane-proximal external region peptides. Vaccine 2015, 33, 861–868. [Google Scholar] [CrossRef] [PubMed]
- Neumann, S.; Young, K.; Compton, B.; Anderson, R.; Painter, G.; Hook, S. Synthetic TRP2 long-peptide and alpha-galactosylceramide formulated into cationic liposomes elicit CD8(+) T-cell responses and prevent tumour progression. Vaccine 2015, 33, 5838–5844. [Google Scholar] [CrossRef] [PubMed]
- Yuba, E.; Kanda, Y.; Yoshizaki, Y.; Teranishi, R.; Harada, A.; Sugiura, K.; Izawa, T.; Yamate, J.; Sakaguchi, N.; Koiwai, K.; et al. pH-sensitive polymer-liposome-based antigen delivery systems potentiated with interferon-gamma gene lipoplex for efficient cancer immunotherapy. Biomaterials 2015, 67, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Yamazaki, D.; Yamauchi, J.; Harashima, H. The nanoparticulation by octaarginine-modified liposome improves α-galactosylceramide-mediated antitumor therapy via systemic administration. J. Control. Release 2013, 171, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Lanzi, A.; Fehres, C.M.; de Gruijl, T.D.; van Kooyk, Y.; Mastrobattista, E. Effects of antigen-expressing immunostimulatory liposomes on chemotaxis and maturation of dendritic cells in Vitro and in human skin explants. Pharmaceut. Res. 2014, 31, 516–526. [Google Scholar] [CrossRef] [PubMed]
- Carrera, I.; Etcheverría, I.; Fernández-Novoa, L.; Lombardi, V.R.; Lakshmana, M.K.; Cacabelos, R.; Vigo, C. A Comparative Evaluation of a Novel Vaccine in APP/PS1 Mouse Models of Alzheimer’s Disease. BioMed Res. Int. 2015. [Google Scholar] [CrossRef] [PubMed]
- Saravanan, P.; Sreenivasa, B.P.; Selvan, R.P.; Basagoudanavar, S.H.; Hosamani, M.; Reddy, N.D.; Nathanielsz, J.; Derozier, C.; Venkataramanan, R. Protective immune response to liposome adjuvanted high potency foot-and-mouth disease vaccine in Indian cattle. Vaccine 2015, 33, 670–677. [Google Scholar] [CrossRef] [PubMed]
- Bolhassani, A.; Javanzad, S.; Saleh, T.; Hashemi, M.; Aghasadeghi, M.R.; Sadat, S.M. Polymeric nanoparticles: Potent vectors for vaccine delivery targeting cancer and infectious diseases. Hum. Vaccines Immunother. 2014, 10, 321–332. [Google Scholar] [CrossRef] [PubMed]
- De Souza Rebouças, J.; Esparza, I.; Ferrer, M.; Sanz, M.L.; Irache, J.M.; Gamazo, C. Nanoparticulate adjuvants and delivery systems for allergen immunotherapy. BioMed Res. Int. 2012. [Google Scholar] [CrossRef] [PubMed]
- Hadinoto, K.; Sundaresan, A.; Cheow, W.S. Lipid–polymer hybrid nanoparticles as a new generation therapeutic delivery platform: A review. Eur. J. Pharm. Biopharm. 2013, 85, 427–443. [Google Scholar] [CrossRef] [PubMed]
- Kunugi, S.; Yamaoka, T. Polymers in Nanomedicine; Springer Science & Business Media: Berlin, Germany, 2012. [Google Scholar]
- Morachis, J.M.; Mahmoud, E.A.; Almutairi, A. Physical and chemical strategies for therapeutic delivery by using polymeric nanoparticles. Pharmacol. Rev. 2012, 64, 505–519. [Google Scholar] [CrossRef] [PubMed]
- Yue, H.; Ma, G. Polymeric micro/nanoparticles: Particle design and potential vaccine delivery applications. Vaccine 2015, 33, 5927–5936. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.L.; Rosalia, R.A.; Varypataki, E.; Sibuea, S.; Ossendorp, F.; Jiskoot, W. Poly-(lactic-co-glycolic-acid)-based particulate vaccines: Particle uptake by dendritic cells is a key parameter for immune activation. Vaccine 2015, 33, 847–854. [Google Scholar] [CrossRef] [PubMed]
- Rahimian, S.; Fransen, M.F.; Kleinovink, J.W.; Christensen, J.R.; Amidi, M.; Hennink, W.E.; Ossendorp, F. Polymeric nanoparticles for co-delivery of synthetic long peptide antigen and poly IC as therapeutic cancer vaccine formulation. J. Control. Release 2015, 203, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Anselmo, A.C.; Banerjee, A.; Zakrewsky, M.; Mitragotri, S. Shape and size-dependent immune response to antigen-carrying nanoparticles. J. Control. Release 2015, 220, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Mueller, S.N.; Tian, S.; DeSimone, J.M. Rapid and persistent delivery of antigen by lymph node targeting PRINT nanoparticle vaccine carrier to promote humoral immunity. Mol. Pharmacol. 2015, 12, 1356–1365. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Shah, S.; Thomas, A.; Ou-Yang, H.D.; Liu, Y. The influence of size, shape and vessel geometry on nanoparticle distribution. Microfluid. Nanofluid. 2013, 14, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Anselmo, A.C.; Zhang, M.; Kumar, S.; Vogus, D.R.; Menegatti, S.; Helgeson, M.E.; Mitragotri, S. Elasticity of nanoparticles influences their blood circulation, phagocytosis, endocytosis, and targeting. ACS Nano 2015, 9, 3169–3177. [Google Scholar] [CrossRef] [PubMed]
- Shima, F.; Akagi, T.; Akashi, M. The role of hydrophobicity in the disruption of erythrocyte membrane by nanoparticles composed of hydrophobically modified poly(gamma-glutamic acid). J. Biomater. Sci. Polym. Ed. 2014, 25, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.M.; Zupancic, E.; Vandermeulen, G.; Oliveira, V.G.; Salgado, A.; Videira, M.; Gaspar, M.; Graca, L.; Préat, V.; Florindo, H.F. In vivo delivery of peptides and Toll-like receptor ligands by mannose-functionalized polymeric nanoparticles induces prophylactic and therapeutic anti-tumor immune responses in a melanoma model. J. Control. Release 2015, 198, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Saluja, S.S.; Hanlon, D.J.; Sharp, F.A.; Hong, E.; Khalil, D.; Robinson, E.; Tigelaar, R.; Fahmy, T.M.; Edelson, R.L. Targeting human dendritic cells via DEC-205 using PLGA nanoparticles leads to enhanced cross-presentation of a melanoma-associated antigen. Int. J. Nanomedicine 2014, 9, 5231–5346. [Google Scholar] [PubMed]
- Rosalia, R.A.; Cruz, L.J.; van Duikeren, S.; Tromp, A.T.; Silva, A.L.; Jiskoot, W.; de Gruijl, T.; Löwik, C.; Oostendorp, J.; van der Burg, S.H.; et al. CD40-targeted dendritic cell delivery of PLGA-nanoparticle vaccines induce potent anti-tumor responses. Biomaterials 2015, 40, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Fang, R.H.; Hu, C.M.; Luk, B.T.; Gao, W.; Copp, J.A.; Tai, Y.; O’Connor, D.E.; Zhang, L. Cancer cell membrane-coated nanoparticles for anticancer vaccination and drug delivery. Nano Lett. 2014, 14, 2181–2188. [Google Scholar] [CrossRef] [PubMed]
- Heo, M.B.; Cho, M.Y.; Lim, Y.T. Polymer nanoparticles for enhanced immune response: Combined delivery of tumor antigen and small interference RNA for immunosuppressive gene to dendritic cells. Acta Biomater. 2014, 10, 2169–2176. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Hoerle, R.; Ehrich, M.; Zhang, C. Engineering the lipid layer of lipid-PLGA hybrid nanoparticles for enhanced in vitro cellular uptake and improved stability. Acta Biomater. 2015, 28, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Ledet, G.; Graves, R.; Datta, D.; Robinson, S.; Bansal, G.P.; Mandal, T.; Kumar, N. Potent functional immunogenicity of plasmodium falciparum transmission-blocking antigen (Pfs25) delivered with nanoemulsion and porous polymeric nanoparticles. Pharmaceut. Res. 2015, 32, 3827–3836. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.J.; Suh, H.; Polhemus, M.E.; Ockenhouse, C.F.; Yadava, A.; Irvine, D.J. Antigen-displaying lipid-enveloped PLGA nanoparticles as delivery agents for a Plasmodium vivax malaria vaccine. PLoS ONE 2012, 7, e31472. [Google Scholar] [CrossRef] [PubMed]
- Orozco, V.H.; Palacio, J.; Sierra, J.; López, B.L. Increased covalent conjugation of a model antigen to poly(lactic acid)-g-maleic anhydride nanoparticles compared to bare poly(lactic acid) nanoparticles. Colloid Polym. Sci. 2013, 291, 2775–2781. [Google Scholar] [CrossRef]
- Rahimian, S.; Kleinovink, J.W.; Fransen, M.F.; Mezzanotte, L.; Gold, H.; Wisse, P.; Overkleeft, H.; Amidi, M.; Jiskoot, W.; Löwik, C.W.; et al. Near-infrared labeled, ovalbumin loaded polymeric nanoparticles based on a hydrophilic polyester as model vaccine: In vivo tracking and evaluation of antigen-specific CD8(+) T cell immune response. Biomaterials 2015, 37, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Kunda, N.K.; Alfagih, I.M.; Miyaji, E.N.; Figueiredo, D.B.; Gonçalves, V.M.; Ferreira, D.M.; Dennison, S.R.; Somavarapu, S.; Hutcheon, G.A.; Saleem, I.Y. Pulmonary dry powder vaccine of pneumococcal antigen loaded nanoparticles. Int. J. Pharm. 2015, 495, 903–912. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Chiu, Y.C.; Tostanoski, L.H.; Jewell, C.M. Polyelectrolyte multilayers assembled entirely from immune signals on gold nanoparticle templates promote antigen-specific T cell response. ACS Nano 2015, 9, 6465–6477. [Google Scholar] [CrossRef] [PubMed]
- Chiu, Y.C.; Gammon, J.M.; Andorko, J.I.; Tostanoski, L.H.; Jewell, C.M. Modular vaccine design using carrier-free capsules assembled from polyionic immune signals. ACS Biomater. Sci. Eng. 2015, 1, 1200–1205. [Google Scholar] [CrossRef] [PubMed]
- Chiodo, F.; Enríquez-Navas, P.M.; Angulo, J.; Marradi, M.; Penadés, S. Assembling different antennas of the gp120 high mannose-type glycans on gold nanoparticles provides superior binding to the anti-HIV antibody 2G12 than the individual antennas. Carbohydr. Res. 2015, 405, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Almeida, J.P.; Lin, A.Y.; Figueroa, E.R.; Foster, A.E.; Drezek, R.A. In vivo gold nanoparticle delivery of peptide vaccine induces anti-tumor immune response in prophylactic and therapeutic tumor models. Small 2015, 11, 1453–1459. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Medina, S.H.; Barchi, J.J. Synthesis and cell-selective antitumor properties of amino acid conjugated tumor-associated carbohydrate antigen-coated gold nanoparticles. Carbohydr. Res. 2015, 405, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Mocan, T.; Matea, C.; Tabaran, F.; Iancu, C.; Orasan, R.; Mocan, L. In vitro administration of gold nanoparticles functionalized with MUC-1 protein fragment generates anticancer vaccine response via macrophage activation and polarization mechanism. J. Cancer 2015, 6, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Tavernaro, I.; Hartmann, S.; Sommer, L.; Hausmann, H.; Rohner, C.; Ruehl, M.; Hoffmann-Roeder, A.; Schlecht, S. Synthesis of tumor-associated MUC1-glycopeptides and their multivalent presentation by functionalized gold colloids. Org. Biomol. Chem. 2015, 13, 81–97. [Google Scholar] [CrossRef] [PubMed]
- Tao, W.; Gill, H.S. M2e-immobilized gold nanoparticles as influenza a vaccine: Role of soluble M2e and longevity of protection. Vaccine 2015, 33, 2307–2315. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Ray, P.C.; Datta, D.; Bansal, G.P.; Angov, E.; Kumar, N. Nanovaccines for malaria using plasmodium falciparum antigen Pfs25 attached gold nanoparticles. Vaccine 2015, 33, 5064–5071. [Google Scholar] [CrossRef] [PubMed]
- Dykman, L.A.; Staroverov, S.A.; Mezhenny, P.V.; Fomin, A.S.; Kozlov, S.V.; Volkov, A.A.; Laskavy, V.N.; Shchyogolev, S.Y. Use of a synthetic foot-and-mouth disease virus peptide conjugated to gold nanoparticles for enhancing immunological response. Gold Bull. 2015, 48, 93–101. [Google Scholar] [CrossRef]
- Gianvincenzo, P.D.; Calvo, J.; Perez, S.; Álvarez, A.; Bedoya, L.M.; Alcamí, J.; Penadés, S. Negatively charged glyconanoparticles modulate and stabilize the secondary structures of a gp120 V3 loop peptide: Toward fully synthetic HIV vaccine candidates. Bioconjugate Chem. 2015, 26, 755–765. [Google Scholar] [CrossRef] [PubMed]
- Sungsuwan, S.; Yin, Z.; Huang, X. Lipopeptide-coated iron oxide nanoparticles as potential glycoconjugate-based synthetic anticancer vaccines. ACS Appl. Mater. Interfaces 2015, 7, 17535–17544. [Google Scholar] [CrossRef] [PubMed]
- Ramella, D.; Polito, L.; Mazzini, S.; Ronchi, S.; Scaglioni, L.; Marelli, M.; Lay, L. A strategy for multivalent presentation of carba analogues from N. meningitidis a capsular polysaccharide. Eur. J. Org. Chem. 2014, 27, 5915–5924. [Google Scholar] [CrossRef]
- den Brok, M.H.; Sutmuller, R.P.; van der Voort, R.; Bennink, E.J.; Figdor, C.G.; Ruers, T.J.; Adema, G.J. In situ tumor ablation creates an antigen source for the generation of antitumor immunity. Cancer Res. 2004, 64, 4024–4029. [Google Scholar] [CrossRef] [PubMed]
- O’Neal, D.P.; Hirsch, L.R.; Halas, N.J.; Payne, J.D.; West, J.L. Photo-thermal tumor ablation in mice using near infrared-absorbing nanoparticles. Cancer Lett. 2004, 209, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Ito, A.; Kobayashi, T. Intracellular hyperthermia using magnetic nanoparticles: A novel method for hyperthermia clinical applications. Therm. Med. 2008, 24, 113–129. [Google Scholar] [CrossRef]
- Kobayashi, T. Cancer hyperthermia using magnetic nanoparticles. Biotech. J. 2011, 6, 1342–1347. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Vaccine Type | Advantages | Disadvantages |
|---|---|---|
| Live attenuated | A single dose of this type of vaccine is more potent as infectious agent can replicate in host. | May cause disease itself. |
| Multiple doses may not be required. | Since vaccine is composed of live organism, storage is very critical. | |
| Since micro-organism itself is used, immune response against all antigens is generated. | Cannot be given to immunosuppressed individuals. | |
| Killed/Inactivated | Safe to use in immunosuppressed patients. | Less immunogenic than live attenuated vaccines. |
| Can’t cause disease state. | May require more booster doses to achieve desired immunity. | |
| Storage conditions are not critical compare to live attenuated vaccines. | ||
| Recombinant/DNA | Better stability compare to traditional vaccines. | High production cost compare to other vaccine types. |
| Storage conditions not critical. | Mutation in host DNA is possible in case of DNA vaccines. | |
| Better control on vaccine design as desired gene can be added or deleted. | ||
| Subunit | Safe to use in immunosuppressed patients. | Less immunogenic than live attenuated vaccines. |
| Cannot cause disease state. | Particular antigen or antigens should be identified causing the disease. | |
| Because of the purified antigenic component, less chances of side-effects. | ||
| Conjugated | Safe to use in immunosuppressed patients. | Conjugation chemistry is difficult to control which could cause batch-wise variation. |
| Cannot cause disease state. | Choice of carrier protein is crucial as they could be immunogenic causing suppression of antigenic immune response. | |
| Because of the purified antigenic component, less chances of side-effects. |
| Disease | Lipid composition | Antigen | Adjuvant | Ref. |
|---|---|---|---|---|
| Pneumonic Plague | DOTAP, DOPE | Ovalbumin (OVA) F1-V recombinant fusion protein of Y. pestis | MPLA | [66] |
| Hepatitis B | SPC, MPC, SA | HBsAg | MPLA | [72] |
| Tuberculosis | DDA | BCG | TDB | [73] |
| Yeast lipids | Alpha crystalline protein 1 (Acr 1) | - | [74] | |
| HIV | DMPC, DMPG, Chol, and MPLA (ALF liposomes) | CN54 gp140 protein | MPLA,QS21 (a triterpenoid glycoside saponin) | [75] |
| DOPC, DOPG, DSPE-PEG | MPER peptide | LACK-1 and HIV-30 (CD4+ epitope) | [76] | |
| Cancer | DOTAP, DOPE, PC, DSPE-PEG | OVA-peptide, TRP-2peptide | Alpha-galactoceramide (α-GC) | [77] |
| EYPC, DOPE, 3-Methyl glutarylated poly(glycidol) (MGluPG) and 3-methylglutarylated dextran | OVA | IFN-g-encoding plasmid DNA | [78] | |
| EPC, DSPE-PEG, Cholesterol, Sterylated R8 | - | Alpha-galactoceramide (α-GC) | [79] | |
| POPE, YSK05, Cholesterol, DMG-PEG | - | Cyclic diGMP | [57] | |
| PC, Cholesterol, (α and β) Galactosyl-DLPE | OVA | [71] | ||
| DPPC, Cholesterol, Rha-TEG-Cholesterol | Tn | Pam3Cys | [67] | |
| DNA vaccine | EPC, Cholesterol, DSPE-PEG | IVTT mix , plasmid DNA (β galactosidase) | IVTT | [80] |
| Alzheimer’s disease | POPG, DOPC, Cholesterol, S1P | Amyloid-beta peptide (A β) | CFA/IFA | [81] |
| Foot and Mouth Disease (FMD) | Lecithin, Cholesterol | Inactivated FMDV | polyinosinic–polycytidylic acid (poly I:C) and oligonucleotide CpGmotif (CpG) | [82] |
| Condition | Sponsor/Collaborators | Status |
|---|---|---|
| Lung cancer | Eastern Cooperative Oncology Grp.; National Cancer Institute (NCI) | Phase II |
| Chronic Lymphocytic Leukemia (CLL) | XEME biopharma Inc.; National Cancer Institute (NCI) | Phase I |
| Tuberculosis | National Institute of Allergy and Infectious Disease (NIAID) | Phase I |
| Non-small cell lung cancer | EMD Serono; Merck KGaA | Phase III |
| Breast cancer | EMD Serono | Phase III |
| Influenza | NasVax Ltd. | Phase II |
| Tuberculosis | Statens Serum Institut | Phase I |
| Disease | Antigen | Immuno-Adjuvant | Modification | Ref. |
|---|---|---|---|---|
| Cancer (Melanoma) | Melan-A:26 , gp100:209 (peptides). OVA as model antigen. | Poly (I:C), CpG | Mannose functionalized delivery system (PLGA, PEG-PLGA and Mannose-PEG-PCL) was developed to target CD206/MR on DC. | [96] |
| MART-1 (peptide) | - | Biotinylated anti-human DEC-205 monoclonal antibodies were used to target DCs. | [97] | |
| Cancer | OVA as model protein antigen | Pam3Csk4, Poly (I:C) | Agonistic α-CD40-mAb were conjugated on the surface of PLGA nanoparticles for CD-40 targeted DC delivery. | [98] |
| Cancer cell membrane obtained from mouse-melanoma cells | - | PLGA nanoparticle were coated with cancer cell membrane to introduce multiple surface antigen which is challenging with traditional synthetic methods. | [99] | |
| OVA as model protein antigen along with SOCS1 siRNA | - | Silencing of immunosuppressive SOCS1 gene augmented pro-inflammatory cytokine response. | [100] | |
| Improved Hybrid polymer-lipid particle | BSA | - | Cholesterol coated PLGA particle showed improved stability with better cellular uptake and more controlled antigen release. | [101] |
| Malaria | Pfs25 (Plasmodium falciparum Transmission-Blocking Antigen) | - | - | [102] |
| VMP001 | MPLA | Lipid (DOPC, DOPG, mal-PE) coated PLGA particles were developed with surface presentation of antigen using maleimide-thiol conjugation. | [103] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vartak, A.; Sucheck, S.J. Recent Advances in Subunit Vaccine Carriers. Vaccines 2016, 4, 12. https://doi.org/10.3390/vaccines4020012
Vartak A, Sucheck SJ. Recent Advances in Subunit Vaccine Carriers. Vaccines. 2016; 4(2):12. https://doi.org/10.3390/vaccines4020012
Chicago/Turabian StyleVartak, Abhishek, and Steven J. Sucheck. 2016. "Recent Advances in Subunit Vaccine Carriers" Vaccines 4, no. 2: 12. https://doi.org/10.3390/vaccines4020012
APA StyleVartak, A., & Sucheck, S. J. (2016). Recent Advances in Subunit Vaccine Carriers. Vaccines, 4(2), 12. https://doi.org/10.3390/vaccines4020012