Abstract
Highly pathogenic avian influenza (HPAI) remains a persistent threat to global poultry production and public health. Current vaccine platforms show limited cross-clade efficacy and often fail to induce mucosal immunity. Recent advances in microbiome research reveal critical roles for gut commensals in modulating vaccine-induced immunity, including enhancement of mucosal IgA production, CD8+ T-cell activation, and modulation of systemic immune responses. Engineered commensal bacteria such as Lactococcus lactis, Bacteroides ovatus, Bacillus subtilis, and Staphylococcus epidermidis have emerged as promising live vectors for antigen delivery. Postbiotic and synbiotic strategies further enhance protective efficacy through targeted modulation of the gut microbiota. Additionally, artificial intelligence (AI)-driven tools enable predictive modeling of host–microbiome interactions, antigen design optimization, and early detection of viral antigenic drift. These integrative technologies offer a new framework for mucosal, broadly protective, and field-deployable vaccines for HPAI control. However, species-specific microbiome variation, ecological safety concerns, and scalable manufacturing remain critical challenges. This review synthesizes emerging evidence on microbiome–immune crosstalk, commensal vector platforms, and AI-enhanced vaccine development, emphasizing the urgent need for One Health integration to mitigate zoonotic adaptation and pandemic emergence.
1. Introduction: The Global HPAI Crisis and Microbiome–Immune Interplay
Highly pathogenic avian influenza (HPAI), particularly H5N1 clade 2.3.4.4b, has triggered unprecedented global epizootics since 2020, resulting in the loss of over 150 million birds in the United States alone and causing widespread economic devastation across the poultry sector [1,2,3,4]. The virus’s rapid antigenic evolution and remarkable geographic spread, highlighted by 297 outbreaks across 26 countries within just April and May 2025, underscore its expanding ecological and epidemiological footprint [4,5,6]. Although traditional vaccines, such as inactivated whole-virus preparations and vectored platforms like a recombinant HPAI-H5 expressed by herpesvirus of Turkeys (HVT-H5) as a vector, have been deployed, they remain suboptimal [3]. Their protection tends to be clade-specific, with cross-clade efficacy dropping below 60% when hemagglutinin sequence similarity declines under 88% [3,7,8]. Moreover, these platforms often fail to induce robust mucosal immunity necessary to block respiratory or fecal–oral transmission and exhibit variable immunogenicity across avian species, for instance, ducks typically display a 3–5-fold lower seroconversion rate than chickens [3,7,8].
Recent discoveries have revealed that the gut microbiome is a key modulator of vaccine-induced immunity in poultry [9,10,11,12,13,14]. Recent studies indicate that specific commensals, such as Lactobacillus crispatus, can elicit systemic and mucosal immune responses to suppress virus infections through both TLR-mediated signaling and B-cell activation, while butyrate of a series of active short-chain fatty acid (SCFA) secreted by microbiota species (such as Faecalibacterium prausnitzii) significantly reduces viral shedding by various mechanisms including macrophage activation and enhancing CD8+ T-cell responses [9,10,11,12,13]. Conversely, microbiota disruption due to antibiotics or heat stress has been associated with impaired mucosal IgA responses and increased mortality following H5N1 exposure [12,13,14]. Additionally, interspecies differences in microbial composition may partially explain immunogenic variability, for example, ducks’ Proteobacteria-rich, aquatic-adapted microbiomes exhibit weaker adjuvant effects compared to the Firmicutes-dominated communities in chickens, necessitating higher vaccine dosages to achieve comparable immunity [9,10,11,12,13,14].
Beyond poultry, the zoonotic implications of HPAI H5N1 are increasingly concerning. Since 2020, infections have been documented in over 48 mammalian species across 26 countries, with significant mortality events in sea lions and widespread outbreaks among dairy cattle in the United States [15,16,17]. High viral loads detected in bovine milk and conjunctival swabs suggest novel transmission pathways, including occupational exposure among dairy workers [15]. Genetic adaptations such as PB2-E627K (enhancing polymerase activity at human body temperatures), HA-T721A (increasing affinity for human-type receptors), and NA-H274Y (conferring antiviral resistance) are now common in mammalian isolates, elevating pandemic risk [3,15,16,17]. Human infections reported by the WHO show a case fatality rate of 48%, with atypical symptoms including conjunctivitis and severe pneumonia [16,17]. In this context, cross-species microbiome insights—such as the role of segmented filamentous bacteria in viral resistance in mice, or bile acid-modulated immune responses in humans—further emphasize the value of commensal-targeted vaccine strategies [9,10,11,12].
The convergence of the HPAI expanding host range and microbiome-driven immune modulation necessitates a paradigm shift in vaccine design. This review synthesizes five years of advances demonstrating how commensal bacteria act as natural adjuvants (e.g., Bacteroides-vectored HA1 delivery), antigen-presenting platforms (e.g., Lactococcus lactis displaying H5 trimers), and predictors of vaccine responsiveness (e.g., Ruminococcus-associated HI titers) [11,13,14]. Integrating these insights with artificial intelligence (AI)-driven antigen refinement—including machine learning-guided epitope focusing and microbiome-mimicking MAMP (i.e., microbe-associated molecular patterns) fusion—offers a promising path toward broadly protective, mucosa-optimized vaccines [3,13]. However, the escalating spillover to mammals demands urgent “One Health” integration, where avian vaccine strategies must account for interfacial microbiomes—spanning poultry, humans, and wild birds—to mitigate zoonotic adaptation and future pandemic emergence [3,11,13]. With a focus on poultry, this review synthesizes recent years of research on microbiomic regulation of vaccine efficacy and explores emerging strategies leveraging commensal bacteria as vectors, adjuvants, and predictive biomarkers for HPAI control, integrated with AI-driven antigen refinement [15,16,17,18].
2. Microbiome–Immune Crosstalk: Implications for HPAI Vaccinology
2.1. Cross-Species Microbiome–Immune Lessons
Insights derived from various animal models have highlighted evolutionarily conserved mechanisms of microbiome-mediated immune regulation. These findings provide critical frameworks for improving poultry vaccines against HPAI infections. In murine models, segmented filamentous bacteria (SFB, e.g., ubiquitous commensal Bacteroides) have been shown to enhance influenza vaccine responses beyond their known role in rotavirus suppression [18,19,20,21]. They achieve this by engaging RANTES/exotoxin-dependent chemokine cascades, which boost systemic immunoglobulin A (IgA) and facilitate the recruitment of CD8+ T-cells to the respiratory mucosa [18,19]. Additionally, the expression of Toll-like receptor 5 (TLR5) in neonatal mice plays a vital role in shaping lifelong microbiota composition. Knockout models have demonstrated that the absence of TLR5 results in impaired antibody responses to influenza vaccination due to a reduction in Clostridia populations [18,19,22]. Furthermore, polysaccharide A (PSA) derived from Bacteroides fragilis has been implicated in correcting Th1/Th2 imbalances via TLR2 signaling, which reduces post-vaccination inflammation while preserving CD8+ cytotoxicity against heterosubtypic influenza strains [18,23,24]. Human studies have provided further insights, revealing that fecal microbiota transplants (FMTs) from high responders to the hepatitis B vaccine can significantly elevate antibody titers in low responders. This effect has been linked to bacteriophage-induced activation of the IL-12/IL-23 axis [25,26]. Additionally, secondary bile acids generated by Clostridium scindens have been shown to enhance rabies vaccine responses by promoting T follicular helper (Tfh) cell differentiation through modulation of CXCR5+ dendritic cells [12]. Conversely, exposure to antibiotics, such as azithromycin, has been shown to reduce H1N1 vaccine seroconversion by 48%, which is attributed to the depletion of Akkermansia muciniphila, a bacterium that typically enhances TLR-dependent signaling capacity for immune regulation [27,28]. In addition, early-life gut microbiome has been associated with positive vaccine efficacy against infections such as rotavirus, plague and influenza in human infants through various mechanisms (Table 1) [18,19,29,30,31].

Table 1.
Examples of cross-species microbiome-mediated antiviral mechanisms [10,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38].
Research on wild birds has uncovered mechanisms relevant to HPAI prevention likely analogical to domestic poultry [32,33,34,35,36]. For instance, gut bacteria expressing α-1,3-galactosyltransferase, such as Escherichia coli O86:B7, induce natural anti-α-Gal antibodies in Anas crecca, providing cross-protection against avian malaria sporozoites via complement-mediated lysis [32,33]. Moreover, shotgun metagenomics of Grus grus has revealed Proteobacteria-dominated microbiomes carrying a high prevalence of the mcr-1 gene, indicating environmental antibiotic pressure that may compromise mucosal immunity during migration [34,35]. Stress induced by migration-mimicking conditions has been shown to deplete Firmicutes and suppress IL-10 levels in Branta canadensis, resulting in increased susceptibility to H5N1 replication in intestinal epithelia [35,36]. Table 1 elaborates on the expanded compendium of microbiome-mediated antiviral mechanisms, summarizing key microbes and metabolites along with their associated immunological outcomes across species [10,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36]. For example, Lactobacillus species activate TLR-signaling pathways, resulting in a 4.1-fold increase in hemagglutination inhibition (HI) titers post-H5 vaccination in chickens [36]. Other notable mechanisms include the production of Reg3 lectins by segmented filamentous bacteria, which lead to a 90% reduction in rotavirus shedding in mice [10,20,30], and the synthesis of secondary bile acids by Clostridium scindens, which promote Tfh and CD8+ T-cell differentiation in humans [37,38].
2.2. Gut Microbiota as Immunomodulators in Poultry
The avian gastrointestinal tract hosts complex microbial ecosystems, with approximately 1012 colony-forming units (CFUs) per gram in chickens and 1010–1011 CFU per gram in the chicken ceca [11,36]. These microbial communities dynamically shape host immunity through various mechanisms. One key aspect of immune priming involves antibody potentiation, where Lactobacillus crispatus upregulates TLR/MyD88-mediated innate immunity and enhances mucosal IgA responses against viral antigens when used as a vaccine delivery platform [11,39]. Concurrently, Bifidobacterium animalis relieves inflammatory response and secretes histone deacetylase inhibitors that enhance chromatin accessibility in B-cell loci, thereby boosting antibody affinity maturation. T-cell polarization is another important mechanism, as butyrate produced by Clostridium Cluster XIVa induces FoxP3+ regulatory T-cells in the ileal Peyer’s patches [36,40,41]. This action suppresses IL-6-driven inflammation following vaccination and promotes CD8+ memory differentiation through the inhibition of HDAC3, sustaining responses to conserved nucleoprotein epitopes. Mucosal barrier fortification is facilitated by IL-22 derived from Bacteroides uniformis, which upregulates occludin and claudin-3 expression, thereby limiting HPAI invasion in chicken [11,36]. Commensal metabolites, such as indole-3-acetic acid, are known to downregulate α-2,3-sialic acid receptors in duck enterocytes, consequently limiting viral attachment. Furthermore, metabolites from Faecalibacterium prausnitzii regulate cellular inflammation, enhancing IFN-γ production by splenic macrophages in vaccinated broilers and priming rapid IL-18 responses upon heterologous influenza challenge [11,36]. Species-specific variations in microbiota responses have also been observed. Ducks, which have aquatic-adapted microbiomes dominated by Proteobacteria, exhibit weaker TLR4 responses compared to chickens, necessitating two-fold higher H5 antigen doses to achieve comparable seroconversion [34,35,36]. In turkeys, the use of butyrate nanogels combined with H7N9 vaccines has been shown to expand antigen-specific T-cells by 5.2-fold through modulation of GPR43-dependent mucosal homing [36,39]. Quail benefit from the combination of Bacillus subtilis spores and galactooligosaccharides, which extend heterosubtypic protection for up to 20 weeks by promoting sustained recruitment of IgA+ plasma cells in the jejunal lamina propria [35,42].
Dysbiosis is a significant factor contributing to vaccine failure in poultry. The administration of amoxicillin in Turkish broilers resulted in a 62% reduction in vaccine-induced IgA and a three-log increase in H5N1 shedding due to the depletion of Faecalibacterium and Bifidobacterium, which are widely recognized as beneficial components of the gut microbiome, playing significant roles in maintaining gut health and influencing the host’s immune system through a variety of mechanisms [11,34,35,36]. Environmental stressors, such as heat stress at temperatures exceeding 35 °C, have been shown to deplete Firmicutes by 40%, thereby blunting IFN-γ and granzyme B responses to H5-inactivated vaccines [11,34,35,36]. Concurrent overgrowth of Escherichia elevates prostaglandin E2 levels, further suppressing dendritic cell maturation. Additionally, H9N2 infection in chickens has been associated with an 80% reduction in mucin-producing Lactobacillus and an expansion of pathogenic Proteobacteria, correlating directly with decreased trefoil factor peptides and impaired mucosal healing [11,36]. Table 2 presents an enhanced compendium of microbiota-mediated immune modulation in poultry, detailing the commensal strains, host species, vaccine platforms, immune outcomes, and key mechanisms involved. For instance, Lactobacillus crispatus administered in chickens receiving H5N2 inactivated vaccines resulted in a 4.1-fold increase in HI titers through TLR2-dependent B-cell activation, while Bacteroides uniformis enhanced mucosal IgA production in turkeys through IL-22 induction in the gut-associated lymphoid tissue (GALT) [11,34,35,36,37,38,39,40,41,42].
Table 2.
Microbiota-mediated immune modulation of poultry vaccines [11,36,37,38,39,40,41,42].
2.3. Synthesis and Forward Vision
The conserved microbiome–immune mechanisms elucidated across species reveal three translational opportunities for next-generation HPAI vaccines. First, precision probiotics tailored to specific species, such as α-Gal-expressing E. coli for ducks or butyrate-producing Clostridia for broilers, could enhance cross-clade immunity by mimicking natural immunomodulatory pathways observed in both murine and avian models [12,13,14,15,16,17,36,37,38,39,40,41,42]. In addition, species-specific microbiome variations may affect vaccine design and efficacy, such as chickens harboring Firmicutes-dominated communities that produce beneficial short-chain fatty acids, and ducks exhibit more diverse microbiomes with higher Proteobacteria abundance [43,44]. These differences affect immune development and vaccine responses, often requiring species-specific vaccination strategies and adjusted antigen doses in waterfowl compared to chickens [45]. Probiotic supplementation with butyrate-producing bacteria can enhance vaccine responses in chickens, while ducks may require modified vaccination protocols [46]. Age-related microbiome maturation further complicates vaccination timing, particularly in young chicks with developing immune systems [47]. Second, AI-guided biomarkers that integrate cecal butyrate levels, Ruminococcus abundance, and anti-α-Gal IgM titers could facilitate the prediction of vaccine responsiveness 4–6 weeks prior to vaccination, enabling flock-specific regimen optimization [11,36,45,46,47]. Lastly, engineered commensal vectors, such as CRISPR-edited Bacteroides ovatus expressing H5 stem antigens, have demonstrated the potential for increasing survival in H5N6-challenged specific pathogen-free chickens and could be delivered orally through feed, thus overcoming cold-chain limitations in smallholder systems [11,48]. Despite these advancements, critical challenges remain in reconciling species-specific microbial ecology, for example, the Proteobacteria-dominant microbiomes of ducks compared to the Firmicutes-rich microbiomes of chickens, with universal vaccine platforms [45,46,47]. Future research must prioritize longitudinal studies that map microbiome trajectories during HPAI outbreaks, in conjunction with gnotobiotic models to isolate strain-level immunomodulators. The integration of One Health surveillance, which monitors wild bird resistomes and mammalian spillover sites, will be essential for preempting microbiome-mediated viral evolution [11,36,48].
3. Commensal Bacteria as Novel Vaccine Vectors: Emerging Platforms for HPAI Control
Commensal bacteria (including probiotic candidates) offer a transformative approach to vaccine delivery by capitalizing on natural host–microbe interactions for targeted mucosal immunization [49,50]. Their innate mucosal tropism, self-adjuvanticity via microbe-associated molecular patterns (MAMPs), and compatibility with mass delivery methods such as oral or water-based administration make them ideal platforms for poultry vaccination targeting on HPAI prevention [49]. Over the recent years, significant advances have been made in engineering these vectors, optimizing antigen presentation systems, and validating protective efficacy in preclinical and early clinical settings [49,50].
3.1. Engineered Probiotic or Commensal Vectors
Among the most developed systems is lactic acid bacteria (LAB), generally including Streptococcus gordonii, Lactococcus lactis, or multiple Lactobacillus species, which have been successfully used as vaccine vectors to express various virus antigens. The attractive advantages for LAB-based vaccine are known as follows: simple, non-invasive administration (usually oral or intranasal), experimental tractability, its fermentative scalability, and safety profile [51,52]. Particularly, several Lactobacillus species have been used to effectively deliver surface-expressed HPAI antigens (e.g., HA or NS1 of H5N1, and HA or M1 and NP of H9N2) to gut-associated lymphoid tissue (GALT), inducing both local (secretory IgA) and systemic (serum IgG) as well as T-cell immune responses in different animal systems including humans, mice, and poultry [51,52,53,54,55,56,57,58]. A recent study in 2023 utilized Lactococcus lactis, a safe and food-grade bacterium, to deliver a chicken IgY-Fc-fused HA1 antigen directly to the mucosal tissues, eliciting a strong mucosal and systemic immune response against H9N2 challenges in chickens [59]. Poultry trials confirm the LAB safety and immunogenicity by expression of HPAI antigens, with ongoing work exploring adjuvant effect, cytokine or antigen co-expression, and lyophilization for feed-based delivery [51]. Despite these promising results, the platform remains in preclinical development due to concerns over biocontainment and the technical challenges of anaerobic cultivation. Kill-switch systems and scalable feed-compatible formulations are under investigation [49,50].
Spore-forming Bacillus subtilis offers practical advantages for vaccine formulation and stability. Spores survive feed pelleting and deliver secreted antigens to the ileum and ceca, evading degradation by gastric acid [60]. A study involving the in-ovo administration of a Bacillus subtilis strain engineered to express hemagglutinin 1 (HA1) and neuraminidase (NA) fusion proteins in broilers demonstrated promising results in protecting against a highly pathogenic avian influenza (H5N8) challenge [61,62]. Specifically, the study observed a 70% survival rate among the broilers that received this treatment when later challenged with the H5N8 virus [62]. The enhanced survival rate in this study was attributed to the robust immune responses generated by the B. subtilis strain expressing the HA1-NA fusion proteins [61,62]. The researchers observed increased levels of ileal IgA and IFN-γ, indicating improved mucosal and cellular immunity, respectively [61,63]. This suggests that the administration of the recombinant B. subtilis stimulated local immune responses in the intestinal tract, a crucial barrier against pathogens like avian influenza viruses [61,62]. Previous research has demonstrated the potential of recombinant B. subtilis as a delivery system for vaccine antigens, inducing both mucosal and systemic immunity in chickens. While promising, it is important to note that the studies concluded that the recombinant B. subtilis did not achieve the same level of protection as commercial vaccines. However, the findings suggest that this approach has significant potential as a biological agent for protecting chickens from avian influenza, and further research may lead to enhanced efficacy. This vector is commercially available as an adjuvant or probiotics (e.g., Alterion®), and next-generation strains are being engineered to co-express antigens and beneficial metabolites like butyrate [64].
Another frontier involves skin commensals such as Staphylococcus epidermidis, which engage cutaneous immunity by activating Langerhans cells via TLR2 signaling. When engineered to display extraneous antigens fusing to the bacterial Aap domain, this bacterium induces neutralizing IgG against expressed antigens and exerts a vaccine effect in mice, with protection linked to dendritic cell activation in draining lymph nodes [65,66]. Therefore, Staphylococcus-based vectors may work through the skin or mucosal pathways to intrinsically enhance immune responses by activating TLR2 pathways and stimulating mucosal IgA, Th1/Th17 polarization, and interferon-stimulated genes (ISGs), which are crucial for antiviral defense for vaccine-design against HPAI in poultry. We recently performed meta-analyses to profile poultry microbiomic datasets and weighted poultry-associated Staphylococcus species for engineering as commensal vaccine vectors using an AI-facilitated comparative matrix (Figure 1). This matrix prioritizes host safety (weighted at 50%), while immune stimulation (Immune) and genetic engineering tractability (Engineering) are each weighted at 25% to reflect their importance for vaccine efficacy and feasibility. The analysis pinpoints that several poultry commensal Staphylococcus species are promising commensal vectors for leveraging as microbiome-based interventions against evolving avian viruses (Figure 1). While still in early preclinical stages, efforts are underway to adapt the commensal vector system for poultry via skin colonization and/or nasal spray delivery formats [65,66].
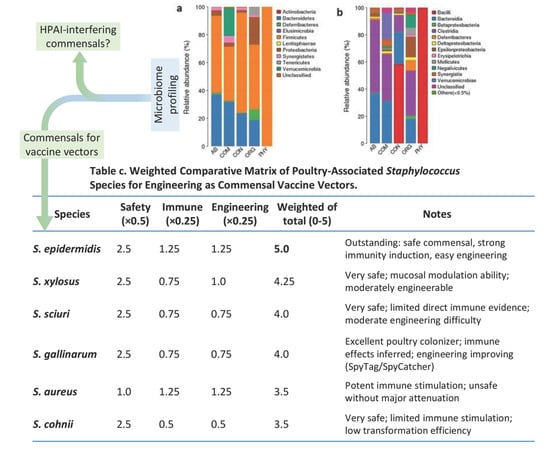
Figure 1.
Targeted microbiome profiling for selecting commensal bacteria with HPAI-interference activity and vaccine vector potential. Upper panel: (a,b) Certain gut commensal bacteria have been shown to enhance antiviral resilience through mechanisms such as maintaining epithelial homeostasis, expelling pathogens, and stimulating innate immune responses, particularly interferon (IFN) production [18,19,26]. However, effective poultry commensal species/strains capable of interfering with highly pathogenic avian influenza (HPAI) remain largely understudied. We propose to integrate meta-analysis and metagenomic profiling approaches to identify 2–4 poultry commensal strains with demonstrated HPAI-interference potential. Bottom panel: In particular, select skin-derived Staphylococcus species/strains will be engineered as vaccine vectors to surface-display HPAI antigens efficiently. Preliminary validation (Panel c) shows our AI-optimized workflow for prioritizing Staphylococcus candidates based on weighted scores emphasizing commensal safety (50%), immune stimulation capacity (25%), and genetic engineering tractability (25%), yielding a weighted total score out of 5. See references for details on the meta-analysis, species selection, and predictive scoring pipeline [49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67].
3.2. Microbiome-Targeted Adjuvants: Synbiotics and Postbiotics
Beyond live bacterial vectors, microbiome-derived products, such as postbiotics and synbiotics, offer complementary immunomodulatory effects that enhance vaccine efficacy. A recent study indicates that lactic acid bacteria produce lactate, which can play an adjuvant role in modulating trained immunity through metabolic and epigenetic rewiring in immune monocytic cells [68]. Postbiotic formulations like butyrate nanogels derived from Clostridium Cluster XIVa improve CD8+ T-cell cytotoxicity via HDAC inhibition and may enhance mucosal immunity when co-administered with HPAI vaccines [69,70]. Bacteria of microbiota (e.g., Vibrio cholera and Bdellovibrio bacteriovorus) possess enzymes similar to the mammalian cyclic GMP-AMP synthase (cGAS), which can synthesize cyclic dinucleotides (CDNs), including cyclic GMP-AMP (cGAMP) to protect bacteria against viral infection [71]. A recent study has shown that cGAMP, as an agonist of cGAS-STING pathway, when used as a mucosal adjuvant with an inactivated H7N9 vaccine, significantly enhanced humoral, cellular, and mucosal immune responses, providing complete protection against homologous virus challenge and inducing cross-protective immunity against H1N1, H3N2, and H9N2, highlighting its potential as a broad-spectrum adjuvant for pre-pandemic influenza vaccines [72]. Similarly, polysaccharide from Atractylodes macrocephala significantly enhanced the HI titer, IgG and specific sIgA levels to reduce viral loads in H9N2-vaccinated chickens [73].
Synbiotic strategies that combine probiotics with targeted prebiotics further enhance protective immunity. Combinations of Bacillus subtilis spores and galactooligosaccharides (GOSs) promote cecal butyrate production and support heterosubtypic protection for up to 20 weeks in broiler chickens [74]. The enhancement correlates strongly with elevated cecal butyrate levels. Another promising approach involves Lactobacillus crispatus paired with mannan-oligosaccharides (MOSs), which activates TLR-MyD88 signaling and alters epigenetic modification in chicken cecal tonsils to potentiate poultry immunity when combined with HPAI-targeted vaccination [68,75,76].
3.3. Challenges and Translational Considerations
Despite promising preclinical results, critical barriers impede the translation of microbiota-based HPAI vaccines from laboratory to commercial application. These barriers include the following: (1) Biocontainment and safety: Genetically engineered bacterial vectors pose significant environmental risks. Containment failures with genetically modified organisms in research settings have highlighted the need for robust biosafety measures [77]. Current mitigation strategies include using non-standard amino acid, though these require extensive validation to prevent evolutionary escape [77,78]. The European Food Safety Authority (EFSA) mandates comprehensive risk assessments for genetically modified bacterial vectors, including physical barriers, genetic safeguards, and environmental monitoring protocols [79,80]. (2) Species-specific efficacy challenges: Real-world deployment reveals critical species variations in vaccine responses. Fundamental differences exist between avian species’ gut microbiomes—chickens typically harbor Firmicutes-dominated communities while waterfowl exhibits more diverse microbiomes with higher Bacteroidetes abundance [44,81,82]. These microbiome differences significantly influence immune responses and vaccine efficacy across species [43]. Consequently, researchers are developing species-specific or broadly protective approaches, though this multiplies production costs and regulatory complexity [83,84,85]. (3) Manufacturing and scalability bottlenecks: Industrial scaling presents substantial technical hurdles. Bacterial spore vaccines show promise but face challenges in large-scale production due to complex fermentation requirements and quality control issues [86]. Live bacterial vaccine systems require cold-chain maintenance throughout production and distribution, significantly increasing costs compared to conventional inactivated vaccines [87]. Production of specialized bacterial vectors remains expensive, with cost-effectiveness being a major barrier to commercial adoption [88]. (4) Regulatory and implementation barriers: Novel bacterial vectors face extensive regulatory scrutiny due to concerns about horizontal gene transfer and environmental release [89]. Field implementation in resource-limited settings reveals additional challenges including cold-chain requirements, farmer acceptance, and infrastructure limitations that affect vaccine efficacy [90]. Regulatory fragmentation across countries creates significant market approval delays, particularly problematic for emergency outbreak responses [79,80,91]. Success requires coordinated solutions including thermostable formulations, harmonized regulatory frameworks, and cost-reduction through platform technologies enabling rapid adaptation for emerging strains [90,91,92].
4. AI-Driven Microbiome Engineering for Vaccine Refinement and Viral Surveillance
Artificial intelligence is revolutionizing microbiome-based vaccine development through validated computational tools that enhance antigen design, predict microbial interactions, and enable real-time monitoring of viral evolution. This rapidly evolving field shows promise for various biomedical applications, and several validated models relevant to microbiome studies and vaccine development are briefed here.
Validated AI Models for Antigen Design: EVEscape, a deep learning model developed by Bloom et al., has demonstrated accuracy in predicting viral escape mutations, with validated performance on influenza and other RNA viruses [93]. This model shows promise for forecasting antigenically significant mutations in viral surface proteins, though its application to microbiome-based vaccine platforms remains in early development stages. AlphaFold and related protein structure prediction tools have shown utility in epitope identification and vaccine design [94,95,96]. Machine learning approaches for epitope prediction have demonstrated improved accuracy over traditional methods, particularly for conserved regions suitable for cross-protective vaccines [97]. These AI-guided approaches can significantly reduce experimental screening time while improving antigen stability in bacterial expression systems [98,99].
Machine Learning for Microbiome Optimization: Convolutional neural networks and other machine learning approaches are increasingly applied to microbiome analysis, showing promise for predicting bacterial colonization patterns and host–microbe interactions [100]. These models can identify optimal probiotic strains for specific applications, though species-specific validation remains essential [101]. Random forest and other ensemble algorithms have shown utility in analyzing metabolomic and microbiome data to predict immune responses, enabling more personalized vaccination strategies [102]. Such approaches could potentially enable real-time dose adjustments based on individual host characteristics [103].
Real-Time Viral Surveillance Integration: GISAID and other genomic surveillance platforms increasingly incorporate automated analysis tools for rapid variant detection and characterization [104]. Machine learning approaches can accelerate phylogenetic analysis and variant identification compared to traditional methods [105]. Time-series modeling and environmental surveillance data show promise for outbreak prediction, though validation across diverse settings remains limited [106]. Integration with vaccination strategies could enable more targeted and preemptive responses to emerging threats [107,108,109].
Current AI applications face significant validation challenges, particularly when applied to novel situations lacking training data [93]. Model performance often decreases when applied to new viral clades or host species, requiring continuous retraining and validation [108]. Cross-validation studies indicate that ensemble approaches combining multiple models often improve robustness compared to individual algorithms [109]. However, continued validation across diverse field conditions remains essential for widespread adoption of AI-driven vaccine development tools [108]. The integration of these AI tools represents progress toward precision vaccinology, though significant challenges remain in translating computational predictions to effective field applications.
5. Future Perspectives and One Health Integration
The convergence of the HPAI expanding zoonotic potential and microbiome-driven vaccine technologies necessitates integrated One Health strategies that coordinate microbiome-based interventions across wildlife, livestock, and human health sectors to prevent pandemic emergence.
First, it might coordinate microbiome-based surveillance and intervention. One Health integration of microbiome-based HPAI vaccines requires coordinated surveillance systems that monitor viral evolution alongside host–microbiome dynamics across species boundaries. Wild bird populations serve as natural reservoirs for influenza viruses, with their gut microbiomes influencing viral persistence and transmission potential [110,111]. Understanding these microbiome–virus interactions across migratory flyways can inform strain selection for poultry vaccines and predict zoonotic adaptation risks [112,113]. The East Asian–Australasian Flyway Partnership provides a framework for such integrated surveillance, though incorporation of microbiome data and coordinated vaccine deployment remains limited [113]. Future systems could integrate real-time monitoring of both viral genetics and host–microbiome composition to predict outbreak risks and guide preemptive vaccination strategies using microbiome-based platforms.
Second, it facilitates cross-species microbiome platform development. Engineered commensal vectors like Lactococcus lactis and Bacillus subtilis offer potential for cross-species vaccine platforms that could protect both livestock and wildlife populations at critical interfaces [15,16,17]. However, species-specific microbiome variations require careful adaptation of these platforms, for example, optimizing Bacteroides-based vectors for waterfowl while maintaining Lactococcus systems for terrestrial poultry [43,44]. In this regard, AI-driven tools can accelerate this cross-species platform development by predicting optimal bacterial characteristics for probiotics and vaccine vector suitably for different host microbiomes [93,108,109]. Such predictive modeling could enable rapid deployment of species-appropriate vaccines during cross-species transmission events, potentially interrupting zoonotic adaptation before human pandemic emergence.
Third, microbiome interventions may mitigate epidemic and zoonotic risks. Critical human–animal interfaces, particularly in poultry processing facilities and live bird markets, represent high-risk zones for zoonotic transmission where coordinated microbiome interventions could provide protection [114]. Probiotic strategies that enhance mucosal immunity in both animals and exposed human populations could create “immune barriers” against emerging epidemic and zoonotic adaptation [115]. The concept of shared microbiome-based immunity across species boundaries remains largely theoretical but represents an important direction for pandemic prevention research. Such approaches would require careful safety evaluation and regulatory coordination across veterinary and human health authorities [116,117,118].
Fourth, it applies to the regulatory and implementation framework of One-Health. Successful One Health integration of microbiome-based HPAI vaccines requires harmonized regulatory frameworks that can rapidly approve and deploy these interventions during emerging outbreak situations. The WHO–FAO–WOAH tripartite collaboration provides a foundation, though specific protocols for microbiome-based interventions need development [119]. Field deployment challenges identified in previous sections—including cold-chain requirements, species-specific efficacy variations, and manufacturing scalability—become more complex in One Health contexts requiring coordination across multiple species and jurisdictions [120,121].
Finally, AI application enhances pandemic preparedness. AI-driven surveillance systems that integrate viral genomics, host–microbiome data, and ecological factors could provide early warning of pandemic-prone viral variants [122]. These systems could automatically trigger deployment of appropriate microbiome-based vaccines across relevant species populations, potentially interrupting pandemic emergence at its source. Machine learning models trained on cross-species transmission data could predict optimal intervention strategies, including which microbiome-based platforms to deploy at specific wildlife–livestock–human interfaces [93,97]. Therefore, future research must address the following: (1) safety and efficacy of microbiome-based vaccines across species boundaries, (2) development of thermostable formulations suitable for diverse deployment conditions, (3) regulatory pathways for rapid emergency deployment, and (4) integration of microbiome interventions with existing surveillance and control systems. This will benefit the ultimate goal to create resilient, ecosystem-level immunity that prevents HPAI variants from achieving pandemic potential through coordinated microbiome-based interventions guided by AI-enhanced surveillance and deployed through integrated One Health systems. While many of these approaches remain largely theoretical or preclinical, they represent important directions for future research and development in pandemic preparedness and response strategies.
Author Contributions
Conceptualization, R.J.W., S.N.N. and Y.S.; methodology, S.N.N. and Y.S.; investigation, S.N.N. and Y.S.; resources, R.J.W. and S.N.N.; data curation, Y.S.; writing—original draft preparation, Y.S.; writing—review and editing, Y.S., R.J.W. and S.N.N.; supervision, Y.S. and S.N.N.; project administration, Y.S.; funding acquisition, R.J.W., S.N.N. and Y.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was primarily supported by USDA NIFA Evans-Allen-1013186, NIFA 2018-67016-28313, and NIFA AFRI 2020-67016-31347 to Y.S., and in part through reagent sharing of NSF-IOS-1831988 to Y.S.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Niu, Q.; Jiang, Z.; Wang, L.; Ji, X.; Baele, G.; Qin, Y.; Lin, L.; Lai, A.; Chen, Y.; Veit, M.; et al. Prevention and control of avian influenza virus: Recent advances in diagnostic technologies and surveillance strategies. Nat. Commun. 2025, 16, 3558. [Google Scholar] [CrossRef]
- Xie, R.; Edwards, K.M.; Wille, M.; Wei, X.; Wong, S.S.; Zanin, M.; El-Shesheny, R.; Ducatez, M.; Poon, L.L.M.; Kayali, G.; et al. The episodic resurgence of highly pathogenic avian influenza H5 virus. Nature 2023, 622, 810–817. [Google Scholar] [CrossRef] [PubMed]
- Kovács, L.; Farkas, M.; Dobra, P.F.; Lennon, G.; Könyves, L.P.; Rusvai, M. Avian Influenza Clade 2.3.4.4b: Global Impact and Summary Analysis of Vaccine Trials. Vaccines 2025, 13, 453. [Google Scholar] [CrossRef]
- Webby, R.J.; Uyeki, T.M. An Update on Highly Pathogenic Avian Influenza A(H5N1) Virus, Clade 2.3.4.4b. J. Infect Dis. 2024, 230, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Yang, J.; Jiao, W.; Li, X.; Iqbal, M.; Liao, M.; Dai, M. 2025. Clade 2.3.4.4b highly pathogenic avian influenza H5N1 viruses: Knowns, unknowns, and challenges. J. Virol. 2025, 99, e0042425. [Google Scholar] [CrossRef] [PubMed]
- Sacristán, C.; Ewbank, A.C.; Ibáñez Porras, P.; Pérez-Ramírez, E.; de la Torre, A.; Briones, V.; Iglesias, I. Novel Epidemiologic Features of High Pathogenicity Avian Influenza Virus A H5N1 2.3.3.4b Panzootic: A Review. Transbound. Emerg. Dis. 2024, 5322378. [Google Scholar] [CrossRef]
- Piesche, R.; Cazaban, C.; Frizzo da Silva, L.; Ramírez-Martínez, L.; Hufen, H.; Beer, M.; Harder, T.; Grund, C. Immunogenicity and Protective Efficacy of Five Vaccines Against Highly Pathogenic Avian Influenza Virus H5N1, Clade 2.3.4.4b, in Fattening Geese. Vaccines 2025, 13, 399. [Google Scholar] [CrossRef] [PubMed]
- Sajjadi, N.C.; Abolnik, C.; Baldinelli, F.; Brown, I.; Cameron, A.; de Wit, S.; Dhingra, M.; Espeisse, O.; Guerin, J.-L.; Harder, T.; et al. Vaccination and surveillance for high pathogenicity avian influenza in poultry-current situation and perspectives. Biologicals 2025, 91, 101840. [Google Scholar] [CrossRef]
- Sheta, B.; Waheed, O.; Ayad, E.; Habbak, L.; Hyder, A. Constitutive immunity is influenced by avian influenza virus-induced modification of gut microbiota in Eurasian teal (Anas crecca). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2024, 278, 109867. [Google Scholar] [CrossRef]
- Alwin, A.; Karst, S.M. The influence of microbiota-derived metabolites on viral infections. Curr. Opin. Virol. 2021, 49, 151–156. [Google Scholar] [CrossRef]
- Abd El-Hack, M.E.; El-Saadony, M.T.; Alqhtani, A.H.; Swelum, A.A.; Salem, H.M.; Elbestawy, A.R.; Noreldin, A.E.; Babalghith, A.O.; Khafaga, A.F.; Hassan, M.I.; et al. The relationship among avian influenza, gut microbiota and chicken immunity: An updated overview. Poult. Sci. 2022, 101, 102021. [Google Scholar] [CrossRef]
- Feng, Y.; de Jong, S.E.; Oliveira, A.P.B.N.; Samaha, H.; Yang, F.; Hu, M.; Wang, Y.; Beydoun, N.; Xie, X.; Zhang, H.; et al. Antibiotic-induced gut microbiome perturbation alters the immune responses to the rabies vaccine. Cell Host Microbe. 2025, 33, 705–718.e5. [Google Scholar] [CrossRef]
- Alqazlan, N.; Astill, J.; Raj, S.; Sharif, S. Strategies for enhancing immunity against avian influenza virus in chickens: A review. Avian Pathol. 2022, 51, 211–235. [Google Scholar] [CrossRef]
- Raj, S.; Alizadeh, M.; Shoojadoost, B.; Hodgins, D.; Nagy, É.; Mubareka, S.; Karimi, K.; Behboudi, S.; Sharif, S. Determining the Protective Efficacy of Toll-Like Receptor Ligands to Minimize H9N2 Avian Influenza Virus Transmission in Chickens. Viruses 2023, 15, 238. [Google Scholar] [CrossRef]
- Plaza, P.I.; Gamarra-Toledo, V.; Euguí, J.R.; Lambertucci, S.A. Recent Changes in Patterns of Mammal Infection with Highly Pathogenic Avian Influenza A(H5N1) Virus Worldwide. Emerg. Infect Dis. 2024, 30, 444–452. [Google Scholar] [CrossRef] [PubMed]
- Gilbertson, B.; Subbarao, K. Mammalian infections with highly pathogenic avian influenza viruses renew concerns of pandemic potential. J. Exp. Med. 2023, 220, e20230447. [Google Scholar] [CrossRef]
- Peacock, T.P.; Moncla, L.; Dudas, G.; VanInsberghe, D.; Sukhova, K.; Lloyd-Smith, J.O.; Worobey, M.; Lowen, A.C.; Nelson, M.I. The global H5N1 influenza panzootic in mammals. Nature 2025, 637, 304–313. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Liwinski, T.; Elinav, E. Interaction between microbiota and immunity in health and disease. Cell Res. 2020, 30, 492–506. [Google Scholar] [CrossRef]
- Li, K.; Ly, K.; Mehta, S.; Braithwaite, A. Importance of crosstalk between the microbiota and the neuroimmune system for tissue homeostasis. Clin. Transl. Immunol. 2022, 11, e1394. [Google Scholar] [CrossRef]
- Ngo, V.L.; Wang, Y.; Wang, Y.; Shi, Z.; Britton, R.; Zou, J.; Ramani, S.; Jiang, B.; Gewirtz, A.T. Select Gut Microbiota Impede Rotavirus Vaccine Efficacy. Cell Mol. Gastroenterol. Hepatol. 2024, 18, 101393. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Huang, Y.; Liu, G.; Li, X.; Xiao, Y.; Liu, C.; Zhang, Y.; Li, J.; Xu, J.; Lu, S.; et al. A Novel Immunobiotics Bacteroides dorei Ameliorates Influenza Virus Infection in Mice. Front. Immunol. 2022, 12, 828887. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.Z.; Ravindran, R.; Chassaing, B.; Carvalho, F.A.; Maddur, M.S.; Bower, M.; Hakimpour, P.; Gill, K.P.; Nakaya, H.I.; Yarovinsky, F.; et al. TLR5-mediated sensing of gut microbiota is necessary for antibody responses to seasonal influenza vaccination. Immunity 2014, 41, 478–492. [Google Scholar] [CrossRef]
- Dasgupta, S.; Erturk-Hasdemir, D.; Ochoa-Reparaz, J.; Reinecker, H.C.; Kasper, D.L. Plasmacytoid dendritic cells mediate anti-inflammatory responses to a gut commensal molecule via both innate and adaptive mechanisms. Cell Host Microbe. 2014, 15, 413–423. [Google Scholar] [CrossRef]
- Ramakrishna, C.; Kujawski, M.; Chu, H.; Li, L.; Mazmanian, S.K.; Cantin, E.M. Bacteroides fragilis polysaccharide A induces IL-10 secreting B and T cells that prevent viral encephalitis. Nat. Commun. 2019, 10, 2153. [Google Scholar] [CrossRef]
- Chauhan, A.; Kumar, R.; Sharma, S.; Mahanta, M.; Vayuuru, S.K.; Nayak, B.; Kumar, S.; Shalimar. Fecal Microbiota Transplantation in Hepatitis B e Antigen-Positive Chronic Hepatitis B Patients: A Pilot Study. Dig. Dis. Sci. 2021, 66, 873–880. [Google Scholar] [CrossRef]
- Liu, Y.; Zhou, J.; Yang, Y.; Chen, X.; Chen, L.; Wu, Y. Intestinal Microbiota and Its Effect on Vaccine-Induced Immune Amplification and Tolerance. Vaccines 2024, 12, 868. [Google Scholar] [CrossRef]
- Hagan, T.; Cortese, M.; Rouphael, N.; Boudreau, C.; Linde, C.; Maddur, M.S.; Das, J.; Wang, H.; Guthmiller, J.; Zheng, N.Y.; et al. Antibiotics-Driven Gut Microbiome Perturbation Alters Immunity to Vaccines in Humans. Cell 2019, 178, 1313–1328.e13. [Google Scholar] [CrossRef]
- Garcia-Vello, P.; Tytgat, H.L.P.; Elzinga, J.; Van Hul, M.; Plovier, H.; Tiemblo-Martin, M.; Cani, P.D.; Nicolardi, S.; Fragai, M.; De Castro, C.; et al. The lipooligosaccharide of the gut symbiont Akkermansia muciniphila exhibits a remarkable structure and TLR signaling capacity. Nat. Commun. 2024, 15, 8411. [Google Scholar] [CrossRef]
- Ryan, F.J.; Clarke, M.; Lynn, M.A.; Benson, S.C.; McAlister, S.; Giles, L.C.; Choo, J.M.; Rossouw, C.; Ng, Y.Y.; Semchenko, E.A.; et al. Bifidobacteria support optimal infant vaccine responses. Nature 2025, 641, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Wagner, J.; Handley, A.; Donato, C.M.; Lyons, E.A.; Pavlic, D.; Ong, D.S.; Bonnici, R.; Bogdanovic-Sakran, N.; Parker, E.P.K.; Bronowski, C.; et al. Early-life gut microbiome associates with positive vaccine take and shedding in neonatal schedule of the human neonatal rotavirus vaccine RV3-BB. Nat. Commun. 2025, 16, 3432. [Google Scholar] [CrossRef] [PubMed]
- Jordan, A.; Carding, S.R.; Hall, L.J. The early-life gut microbiome and vaccine efficacy. Lancet Microbe. 2022, 3, e787–e794. [Google Scholar] [CrossRef] [PubMed]
- Palinauskas, V.; Mateos-Hernandez, L.; Wu-Chuang, A.; de la Fuente, J.; Aželytė, J.; Obregon, D.; Cabezas-Cruz, A. Exploring the Ecological Implications of Microbiota Diversity in Birds: Natural Barriers Against Avian Malaria. Front. Immunol. 2022, 13, 807682. [Google Scholar] [CrossRef]
- Yilmaz, B.; Portugal, S.; Tran, T.M.; Gozzelino, R.; Ramos, S.; Gomes, J.; Regalado, A.; Cowan, P.J.; d’Apice, A.J.; Chong, A.S.; et al. Gut microbiota elicits a protective immune response against malaria transmission. Cell 2014, 159, 1277–1289. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Hu, Y.; Liu, F.; Wang, Y.; Bi, Y.; Lv, N.; Li, J.; Zhu, B.; Gao, G.F. Metagenomic analysis reveals the microbiome and resistome in migratory birds. Microbiome 2020, 8, 26. [Google Scholar] [CrossRef] [PubMed]
- Gul, S.; Shi, Y.; Hu, J.; Song, S. The Influence of Microbiota on Wild Birds’ Parental Coprophagy Behavior: Current Advances and Future Research Directions. Microorganisms 2024, 12, 2468. [Google Scholar] [CrossRef]
- Abaidullah, M.; Peng, S.; Kamran, M.; Song, X.; Yin, Z. Current Findings on Gut Microbiota Mediated Immune Modulation against Viral Diseases in Chicken. Viruses 2019, 11, 681. [Google Scholar] [CrossRef]
- Alavi, S.; Mitchell, J.D.; Cho, J.Y.; Liu, R.; Macbeth, J.C.; Hsiao, A. Interpersonal Gut Microbiome Variation Drives Susceptibility and Resistance to Cholera Infection. Cell 2020, 181, 1533–1546.e13. [Google Scholar] [CrossRef]
- Jin, W.B.; Xiao, L.; Jeong, M.; Han, S.J.; Zhang, W.; Yano, H.; Shi, H.; Arifuzzaman, M.; Lyu, M.; Wang, D.; et al. Microbiota-derived bile acids antagonize the host androgen receptor and drive anti-tumor immunity. Cell 2025, 188, 2336–2353.e38. [Google Scholar] [CrossRef]
- Ullah, H.; Arbab, S.; Tian, Y.; Chen, Y.; Liu, C.Q.; Li, Q.; Li, K. Crosstalk between gut microbiota and host immune system and its response to traumatic injury. Front. Immunol. 2024, 15, 1413485. [Google Scholar] [CrossRef]
- Paradowska, M.; Dunislawska, A.; Siwek, M.; Slawinska, A. Avian Cell Culture Models to Study Immunomodulatory Properties of Bioactive Products. Animals 2022, 12, 670. [Google Scholar] [CrossRef]
- Idowu, P.A.; Mpofu, T.J.; Magoro, A.M.; Modiba, M.C.; Nephawe, K.A.; Mtileni, B. Impact of probiotics on chicken gut microbiota, immunity, behavior, and productive performance—A systematic review. Front. Anim. Sci. 2025, 6, 1562527. [Google Scholar] [CrossRef]
- Sultan, A.; Murtaza, S.; Naz, S.; Islam, Z.; Alrefaei, A.F.; Khan, R.U.; HAbdelrahman, S.; Chandrasekaran, A. Live performance, nutrient digestibility, immune response and fecal microbial load modulation in Japanese quails fed a Bacillus-based probiotic alone or combination with xylanase. Vet. Q. 2024, 44, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Stanley, D.; Hughes, R.J.; Moore, R.J. Microbiota of the chicken gastrointestinal tract. Appl. Microbiol. Biotechnol. 2014, 98, 4301–4310. [Google Scholar] [CrossRef] [PubMed]
- Waite, D.W.; Taylor, M.W. Characterizing the avian gut microbiota: Membership, driving influences, and potential function. Front. Microbiol. 2014, 5, 223. [Google Scholar] [CrossRef]
- Pantin-Jackwood, M.J.; Swayne, D.E. Pathogenesis and pathobiology of avian influenza virus infection in birds. Rev. Sci. Tech. 2009, 28, 113–136. [Google Scholar] [CrossRef] [PubMed]
- Brisbin, J.T.; Gong, J.; Sharif, S. Interactions between commensal bacteria and the gut-associated immune system of the chicken. Anim. Health Res. Rev. 2008, 9, 101–110. [Google Scholar] [CrossRef]
- Crhanova, M.; Hradecka, H.; Faldynova, M.; Matulova, M.; Havlickova, H.; Sisak, F.; Rychlik, I. Immune response of chicken gut to natural colonization by gut microflora. Infect. Immun. 2011, 79, 2755–2763. [Google Scholar] [CrossRef]
- Wang, W.C.; Sayedahmed, E.E.; Sambhara, S.; Mittal, S.K. Progress towards the Development of a Universal Influenza Vaccine. Viruses 2022, 14, 1684. [Google Scholar] [CrossRef]
- Lloren, K.K.S.; Senevirathne, A.; Lee, J.H. Advancing vaccine technology through the manipulation of pathogenic and commensal bacteria. Mater. Today Bio 2024, 29, 101349. [Google Scholar] [CrossRef]
- Xu, C.; Aqib, A.I.; Fatima, M.; Muneer, S.; Zaheer, T.; Peng, S.; Ibrahim, E.H.; Li, K. Deciphering the Potential of Probiotics in Vaccines. Vaccines 2024, 12, 711. [Google Scholar] [CrossRef]
- LeCureux, J.S.; Dean, G.A. Lactobacillus Mucosal Vaccine Vectors: Immune Responses against Bacterial and Viral Antigens. mSphere 2018, 3, e00061-18. [Google Scholar] [CrossRef]
- Wang, Z.S.; Yu, Q.H.; Gao, J.K.; Yang, Q. Mucosal and systemic immune responses induced by recombinant Lactobacillus spp. expressing the hemagglutinin of the avian influenza virus H5N1. Clin. Vaccine Immunol. 2012, 19, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.H.; Yang, W.T.; Yang, G.L.; Cong, Y.L.; Huang, H.B.; Wang, Q.; Cai, R.P.; Ye, L.P.; Hu, J.T.; Zhou, J.Y.; et al. Immunoprotection against influenza virus H9N2 by the oral administration of recombinant Lactobacillus plantarumNC8 expressing hemagglutinin in BALB/c mice. Virology 2014, 464–465, 166–176. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.H.; Yang, W.T.; Yang, G.L.; Zhang, X.K.; Liu, Y.Y.; Zhang, L.J.; Ye, L.P.; Hu, J.T.; Xing, X.; Qi, C.; et al. Lactobacillus plantarum vaccine vector expressing hemagglutinin provides protection against H9N2 challenge infection. Virus Res. 2016, 211, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Levit, R.; Cortes-Perez, N.G.; de Moreno de Leblanc, A.; Loiseau, J.; Aucouturier, A.; Langella, P.; LeBlanc, J.G.; Bermúdez-Humarán, L.G. Use of genetically modified lactic acid bacteria and bifidobacteria as live delivery vectors for human and animal health. Gut Microbes. 2022, 14, 2110821. [Google Scholar] [CrossRef]
- Wang, Z.; Yu, Q.; Fu, J.; Liang, J.; Yang, Q. Immune responses of chickens inoculated with recombinant Lactobacillus expressing the haemagglutinin of the avian influenza virus. J. Appl. Microbiol. 2013, 115, 1269–1277. [Google Scholar] [CrossRef]
- Yang, W.T.; Shi, S.H.; Yang, G.L.; Jiang, Y.L.; Zhao, L.; Li, Y.; Wang, C.F. Cross-protective efficacy of dendritic cells targeting conserved influenza virus antigen expressed by Lactobacillus plantarum. Sci. Rep. 2016, 6, 39665. [Google Scholar] [CrossRef]
- Tan, T.S.; Syed Hassan, S.; Yap, W.B. Expression of surface-bound nonstructural 1 (NS1) protein of influenza virus A H5N1 on Lactobacillus casei strain C1. Lett. Appl. Microbiol. 2017, 64, 446–451. [Google Scholar] [CrossRef]
- Zhang, R.; Xu, T.; Li, Z.; Li, L.; Li, C.; Li, X.; Wang, Z.; Wang, S.; Wang, X.; Zhang, H. Vaccination with recombinant Lactococcus lactis expressing HA1-IgY Fc fusion protein provides protective mucosal immunity against H9N2 avian influenza virus in chickens. Virol. J. 2023, 20, 76. [Google Scholar] [CrossRef]
- Ogbuewu, I.P.; Mabelebele, M.; Sebola, N.A.; Mbajiorgu, C. Bacillus Probiotics as Alternatives to In-feed Antibiotics and Its Influence on Growth, Serum Chemistry, Antioxidant Status, Intestinal Histomorphology, and Lesion Scores in Disease-Challenged Broiler Chickens. Front. Vet. Sci. 2022, 9, 876725. [Google Scholar] [CrossRef]
- Mou, C.; Zhu, L.; Yang, J.; Xu, W.; Cheng, X.; Yang, Q. Immune Responses Induced by Recombinant Bacillus Subtilis Expressing the Hemagglutinin Protein of H5N1 in chickens. Sci. Rep. 2016, 6, 38403. [Google Scholar] [CrossRef]
- Rudometova, N.B.; Fando, A.A.; Kisakova, L.A.; Kisakov, D.N.; Borgoyakova, M.B.; Litvinova, V.R.; Yakovlev, V.A.; Tigeeva, E.V.; Vahitov, D.I.; Sharabrin, S.V.; et al. Immunogenic and Protective Properties of Recombinant Hemagglutinin of Influenza A (H5N8) Virus. Vaccines 2024, 12, 143. [Google Scholar] [CrossRef]
- Lv, P.; Zhang, X.; Song, M.; Hao, G.; Wang, F.; Sun, S. Oral administration of recombinant Bacillus subtilis expressing a multi-epitope protein induces strong immune responses against Salmonella Enteritidis. Vet. Microbiol. 2023, 276, 109632. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP); Rychen, G.; Aquilina, G.; Azimonti, G.; Bampidis, V.; Bastos, M.L.; Bories, G.; Chesson, A.; Cocconcelli, P.S.; Flachowsky, G.; et al. Safety and efficacy of Alterion NE® (Bacillus subtilis DSM 29784) as a feed additive for chickens for fattening and chickens reared for laying. EFSA J. 2017, 15, e04933. [Google Scholar] [CrossRef]
- Gribonika, I.; Band, V.I.; Chi, L.; Perez-Chaparro, P.J.; Link, V.M.; Ansaldo, E.; Oguz, C.; Bousbaine, D.; Fischbach, M.A.; Belkaid, Y. Skin autonomous antibody production regulates host-microbiota interactions. Nature 2025, 638, 1043–1053. [Google Scholar] [CrossRef] [PubMed]
- Bousbaine, D.; Bauman, K.D.; Chen, Y.E.; Lalgudi, P.V.; Nguyen, T.T.D.; Swenson, J.M.; Yu, V.K.; Tsang, E.; Conlan, S.; Li, D.B.; et al. Discovery and engineering of the antibody response to a prominent skin commensal. Nature 2025, 638, 1054–1064. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Animal Health and Animal Welfare (AHAW); European Union Reference Laboratory for Avian Influenza; Nielsen, S.S.; Alvarez, J.; Bicout, D.J.; Calistri, P.; Canali, E.; Drewe, J.A.; Garin-Bastuji, B.; Gonzales Rojas, J.L.; et al. Vaccination of poultry against highly pathogenic avian influenza—Part 1. Available vaccines and vaccination strategies. EFSA J. 2023, 21, e08271. [Google Scholar] [CrossRef]
- Ziogas, A.; Novakovic, B.; Ventriglia, L.; Galang, N.; Tran, K.A.; Li, W.; Matzaraki, V.; van Unen, N.; Schlüter, T.; Ferreira, A.V.; et al. Long-term histone lactylation connects metabolic and epigenetic rewiring in innate immune memory. Cell 2025, 188, 2992–3012. [Google Scholar] [CrossRef]
- He, Y.; Fu, L.; Li, Y.; Wang, W.; Gong, M.; Zhang, J.; Dong, X.; Huang, J.; Wang, Q.; Mackay, C.R.; et al. Gut microbial metabolites facilitate anticancer therapy efficacy by modulating cytotoxic CD8+ T cell immunity. Cell Metab. 2021, 33, 988–1000.e7. [Google Scholar] [CrossRef]
- Bodman-Harris, O.; Rollier, C.S.; Iqbal, M. Approaches to Enhance the Potency of Vaccines in Chickens. Vaccines 2024, 12, 1337. [Google Scholar] [CrossRef]
- Cohen, D.; Melamed, S.; Millman, A.; Shulman, G.; Oppenheimer-Shaanan, Y.; Kacen, A.; Doron, S.; Amitai, G.; Sorek, R. Cyclic GMP-AMP signalling protects bacteria against viral infection. Nature 2019, 574, 691–695. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Liu, X.; Xiong, F.; Gao, F.; Yi, Y.; Zhang, M.; Chen, Z.; Tan, W. Enhancing Immune Response and Heterosubtypic Protection Ability of Inactivated H7N9 Vaccine by Using STING Agonist as a Mucosal Adjuvant. Front. Immunol. 2019, 10, 2274. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Hong, H.; Wang, J.; Huang, J.; Li, J.; Tao, Y.; Liu, M.; Pang, H.; Li, J.; Bo, R. Mucosal immune responses and protective efficacy elicited by oral administration AMP-ZnONPs-adjuvanted inactivated H9N2 virus in chickens. Poult. Sci. 2024, 103, 103496. [Google Scholar] [CrossRef] [PubMed]
- Khosravi, A.; Boldaji, F.; Dastar, B.; Torshizi, M.A.K.; Alemi, M.; Hoseinifar, S.H. A synbiotic improves performance and gut health in broiler chickens. Sci. Rep. 2025, 15, 19164. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.Z.; Li, F.; Huang, X.; Nouman, M.; Bibi, R.; Fan, X.; Zhou, H.; Shan, Z.; Wang, L.; Jiang, Y.; et al. Oral Immunization of Chickens with Probiotic Lactobacillus crispatus Constitutively Expressing the α-β2-ε-β1 Toxoids to Induce Protective Immunity. Vaccines 2022, 10, 698. [Google Scholar] [CrossRef]
- Dunislawska, A.; Slawinska, A.; Siwek, M. Hepatic DNA Methylation in Response to Early Stimulation of Microbiota with Lactobacillus Synbiotics in Broiler Chickens. Genes 2020, 11, 579. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Torres, L.; Krüger, A.; Csibra, E.; Gianni, E.; Pinheiro, V.B. Synthetic biology approaches to biological containment: Pre-emptively tackling potential risks. Essays Biochem. 2016, 60, 393–410. [Google Scholar] [CrossRef]
- Forti, A.M.; Jones, M.A.; Elbeyli, D.N.; Butler, N.D.; Kunjapur, A.M. Engineered orthogonal and obligate bacterial commensalism mediated by a non-standard amino acid. Nat. Microbiol. 2025, 10, 1404–1416. [Google Scholar] [CrossRef] [PubMed]
- EFSA (European Food Safety Authority). Guidance on the assessment of genetically modified microorganisms and their products intended for food and feed use. EFSA J. 2011, 9, 2193. [Google Scholar] [CrossRef]
- EFSA Panel on Biological Hazards (BIOHAZ); Koutsoumanis, K.; Allende, A.; Alvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; De Cesare, A.; Hilbert, F.; Lindqvist, R.; et al. Update of the list of qualified presumption of safety (QPS) recommended microbiological agents intentionally added to food or feed as notified to EFSA 20: Suitability of taxonomic units notified to EFSA until March 2024. EFSA J. 2024, 22, e8882. [Google Scholar] [CrossRef]
- Maraci, Ö.; Antonatou-Papaioannou, A.; Jünemann, S.; Castillo-Gutiérrez, O.; Busche, T.; Kalinowski, J.; Caspers, B.A. The Gut Microbial Composition Is Species-Specific and Individual-Specific in Two Species of Estrildid Finches, the Bengalese Finch and the Zebra Finch. Front. Microbiol. 2021, 12, 619141. [Google Scholar] [CrossRef] [PubMed]
- Bodawatta, K.H.; Koane, B.; Maiah, G.; Sam, K.; Poulsen, M.; Jønsson, K.A. Species-specific but not phylosymbiotic gut microbiomes of New Guinean passerine birds are shaped by diet and flight-associated gut modifications. Proc. Biol. Sci. 2021, 288, 20210446. [Google Scholar] [CrossRef]
- Qin, C.; Gong, L.; Zhang, X.; Wang, Y.; Wang, Y.; Wang, B.; Li, Y.; Li, W. Effect of Saccharomyces boulardii and Bacillus subtilis B10 on gut microbiota modulation in broilers. Anim. Nutr. 2018, 4, 358–366. [Google Scholar] [CrossRef]
- Liu, Y.; Zhao, D.; Zhang, J.; Huang, X.; Han, K.; Liu, Q.; Yang, J.; Zhang, L.; Li, Y. Development of an Inactivated Avian Influenza Virus Vaccine against Circulating H9N2 in Chickens and Ducks. Vaccines 2023, 11, 596. [Google Scholar] [CrossRef]
- Chen, T.H.; Yang, Y.L.; Jan, J.T.; Chen, C.C.; Wu, S.C. Site-Specific Glycan-Masking/Unmasking Hemagglutinin Antigen Design to Elicit Broadly Neutralizing and Stem-Binding Antibodies Against Highly Pathogenic Avian Influenza H5N1 Virus Infections. Front. Immunol. 2021, 12, 692700. [Google Scholar] [CrossRef]
- Todorov, S.D.; Ivanova, I.V.; Popov, I.; Weeks, R.; Chikindas, M.L. Bacillus spore-forming probiotics: Benefits with concerns? Crit. Rev. Microbiol. 2022, 48, 513–530. [Google Scholar] [CrossRef]
- Li, L.; Yang, L.; Zhang, L.; He, F.; Xia, Z.; Xiang, B. Multi-omic analysis reveals that Bacillus licheniformis enhances pekin ducks growth performance via lipid metabolism regulation. Front. Pharmacol. 2024, 15, 1412231. [Google Scholar] [CrossRef]
- Cargnin Faccin, F.; Perez, D.R. Pandemic preparedness through vaccine development for avian influenza viruses. Hum. Vaccin. Immunother. 2024, 20, 2347019. [Google Scholar] [CrossRef] [PubMed]
- Mohamadzadeh, M.; Pfeiler, E.A.; Brown, J.B.; Zadeh, M.; Gramarossa, M.; Managlia, E.; Bere, P.; Sarraj, B.; Khan, M.W.; Pakanati, K.C.; et al. Regulation of induced colonic inflammation by Lactobacillus acidophilus deficient in lipoteichoic acid. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4623–4630. [Google Scholar] [CrossRef] [PubMed]
- Swayne, D.E.; Kapczynski, D. Strategies and challenges for eliciting immunity against avian influenza virus in birds. Immunol. Rev. 2008, 225, 314–331. [Google Scholar] [CrossRef] [PubMed]
- Capua, I.; Alexander, D.J. Avian influenza infection in birds: A challenge and opportunity for the poultry veterinarian. Poult. Sci. 2009, 88, 842–846. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Gao, G.F. A brief history of human infections with H5Ny avian influenza viruses. Cell Host Microbe. 2025, 33, 176–181. [Google Scholar] [CrossRef]
- Thadani, N.N.; Gurev, S.; Notin, P.; Youssef, N.; Rollins, N.J.; Ritter, D.; Sander, C.; Gal, Y.; Marks, D.S. Learning from prepandemic data to forecast viral escape. Nature 2023, 622, 818–825. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Notin, P.; Marks, D.S.; Weitzman, R.; Gal, Y. ProteinNPT: Improving Protein Property Prediction and Design with Non-Parametric Transformers. bioRxiv 2023. [Google Scholar] [CrossRef]
- Hayes, T.; Rao, R.; Akin, H.; Sofroniew, N.J.; Oktay, D.; Lin, Z.; Verkuil, R.; Tran, V.Q.; Deaton, J.; Wiggert, M.; et al. Simulating 500 million years of evolution with a language model. Science 2025, 387, 850–858. [Google Scholar] [CrossRef]
- Bukhari, S.N.H.; Jain, A.; Haq, E.; Mehbodniya, A.; Webber, J. Machine Learning Techniques for the Prediction of B-Cell and T-Cell Epitopes as Potential Vaccine Targets with a Specific Focus on SARS-CoV-2 Pathogen: A Review. Pathogens 2022, 11, 146. [Google Scholar] [CrossRef] [PubMed]
- Patronov, A.; Doytchinova, I. T-cell epitope vaccine design by immunoinformatics. Open Biol. 2013, 3, 120139. [Google Scholar] [CrossRef]
- Cia, G.; Pucci, F.; Rooman, M. Critical review of conformational B-cell epitope prediction methods. Brief. Bioinform. 2023, 24, bbac567. [Google Scholar] [CrossRef]
- Reiman, D.; Metwally, A.A.; Sun, J.; Dai, Y. PopPhy-CNN: A phylogenetic tree embedded architecture for convolutional neural networks to predict host phenotype from metagenomic data. IEEE J. Biomed. Health Inform. 2017, 24, 2993–3001. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Haiminen, N.; Carrieri, A.P.; Hu, R.; Jiang, L.; Parida, L.; Russell, B.; Allaband, C.; Zarrinpar, A.; Vázquez-Baeza, Y.; et al. Human Skin, Oral, and Gut Microbiomes Predict Chronological Age. mSystems 2020, 5, e00630-19. [Google Scholar] [CrossRef]
- Thaiss, C.A.; Zmora, N.; Levy, M.; Elinav, E. The microbiome and innate immunity. Nature 2016, 535, 65–74. [Google Scholar] [CrossRef]
- Lynch, S.V.; Pedersen, O. The human intestinal microbiome in health and disease. N. Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef]
- Khare, S.; Gurry, C.; Freitas, L.; Schultz, M.B.; Bach, G.; Diallo, A.; Akite, N.; Ho, J.; Lee, R.T.; Yeo, W.; et al. GISAID’s Role in Pandemic Response. China CDC Wkly. 2021, 3, 1049–1051. [Google Scholar] [CrossRef] [PubMed]
- Hadfield, J.; Megill, C.; Bell, S.M.; Huddleston, J.; Potter, B.; Callender, C.; Sagulenko, P.; Bedford, T.; Neher, R.A. Nextstrain: Real-time tracking of pathogen evolution. Bioinformatics 2018, 34, 4121–4123. [Google Scholar] [CrossRef] [PubMed]
- Viboud, C.; Sun, K.; Gaffey, R.; Ajelli, M.; Fumanelli, L.; Merler, S.; Zhang, Q.; Chowell, G.; Simonsen, L.; Vespignani, A.; et al. The RAPIDD Ebola forecasting challenge: Synthesis and lessons learnt. Epidemics 2018, 22, 13–21. [Google Scholar] [CrossRef]
- Shah, S.A.W.; Palomar, D.P.; Barr, I.; Poon, L.L.M.; Quadeer, A.A.; McKay, M.R. Seasonal antigenic prediction of influenza A H3N2 using machine learning. Nat. Commun. 2024, 15, 3833. [Google Scholar] [CrossRef] [PubMed]
- Ching, T.; Himmelstein, D.S.; Beaulieu-Jones, B.K.; Kalinin, A.A.; Do, B.T.; Way, G.P.; Ferrero, E.; Agapow, P.-M.; Zietz, M.; Hoffman, M.M.; et al. Opportunities and obstacles for deep learning in biology and medicine. J. R. Soc. Interface 2018, 15, 20170387. [Google Scholar] [CrossRef]
- Wang, S.; Sun, S.; Li, Z.; Zhang, R.; Xu, J. Accurate De Novo Prediction of Protein Contact Map by Ultra-Deep Learning Model. PLoS Comput. Biol. 2017, 13, e1005324. [Google Scholar] [CrossRef]
- Zamani, O.; Bittmann, T.; Ortega, D.L. The effect of avian influenza outbreaks on retail price premiums in the United States poultry market. Poult. Sci. 2024, 103, 104102. [Google Scholar] [CrossRef]
- van Dijk, J.G.; Hoye, B.J.; Verhagen, J.H.; Nolet, B.A.; Fouchier, R.A.; Klaassen, M. Juveniles and migrants as drivers for seasonal epizootics of avian influenza virus. J. Anim. Ecol. 2014, 83, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Verhagen, J.H.; van der Jeugd, H.P.; Nolet, B.A.; Slaterus, R.; Kharitonov, S.P.; de Vries, P.P.; Vuong, O.; Majoor, F.; Kuiken, T.; Fouchier, R.A. Wild bird surveillance around outbreaks of highly pathogenic avian influenza A(H5N8) virus in the Netherlands, 2014, within the context of global flyways. Euro. Surveill. 2015, 20, 21069. [Google Scholar] [CrossRef]
- Dupas, M.-C.; Vincenti-Gonzalez, M.F.; Dhingra, M.; Guinat, C.; Vergne, T.; Wint, W.; Hendrickx, G.; Marsboom, C.; Gilbert, M.; Dellicour, S. Global risk mapping of highly pathogenic avian influenza H5N1 and H5Nx in the light of epidemic episodes occurring from 2020 onward. Epidemiol. Glob. Health 2025, 14, RP104748. [Google Scholar] [CrossRef]
- Rabinowitz, P.; Kock, R.; Kachani, M.; Kunkel, R.; Thomas, J.; Gilbert, J.; Wallace, R.; Blackmore, C.; Wong, D.; Karesh, W.; et al. Toward proof of concept of a One Health approach to disease prediction and control. Emerg. Infect. Dis. 2013, 19, e130265. [Google Scholar] [CrossRef] [PubMed]
- Hemarajata, P.; Versalovic, J. Effects of probiotics on gut microbiota: Mechanisms of intestinal immunomodulation and neuromodulation. Ther. Adv. Gastroenterol. 2013, 6, 39–51. [Google Scholar] [CrossRef]
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global trends in emerging infectious diseases. Nature 2008, 451, 990–993. [Google Scholar] [CrossRef]
- Atarashi, K.; Tanoue, T.; Oshima, K.; Suda, W.; Nagano, Y.; Nishikawa, H.; Fukuda, S.; Saito, T.; Narushima, S.; Hase, K.; et al. Treg induction by a rationally selected mixture of Clostridia strains from the human microbiota. Nature 2013, 500, 232–236. [Google Scholar] [CrossRef]
- Riglar, D.T.; Giessen, T.W.; Baym, M.; Kerns, S.J.; Niederhuber, M.J.; Bronson, R.T.; Kotula, J.W.; Gerber, G.K.; Way, J.C. Engineered bacteria can function in the mammalian gut long-term as live diagnostics of inflammation. Nat. Biotechnol. 2017, 35, 653–658. [Google Scholar] [CrossRef]
- Mackenzie, J.S.; Jeggo, M. The One Health approach—why is it so important? Trop. Med. Infect. Dis. 2019, 4, 88. [Google Scholar] [CrossRef]
- Lerner, H.; Berg, C. The concept of health in One Health and some practical implications for research and education: What is One Health? Infect. Ecol. Epidemiol. 2015, 5, 25300. [Google Scholar] [CrossRef]
- Zinsstag, J.; Schelling, E.; Waltner-Toews, D.; Tanner, M. From “one medicine” to “one health” and systemic approaches to health and well-being. Prev. Vet. Med. 2011, 101, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Lewis, N.S.; Russell, C.A.; Langat, P.; Anderson, T.K.; Berger, K.; Bielejec, F.; Burke, D.F.; Dudas, G.; Fonville, J.M.; Fouchier, R.A.; et al. The global antigenic diversity of swine influenza A viruses. Elife 2016, 5, e12217. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).