
Vaccines and Animal Models of Nipah Virus: Current Situation and Future Prospects


Abstract
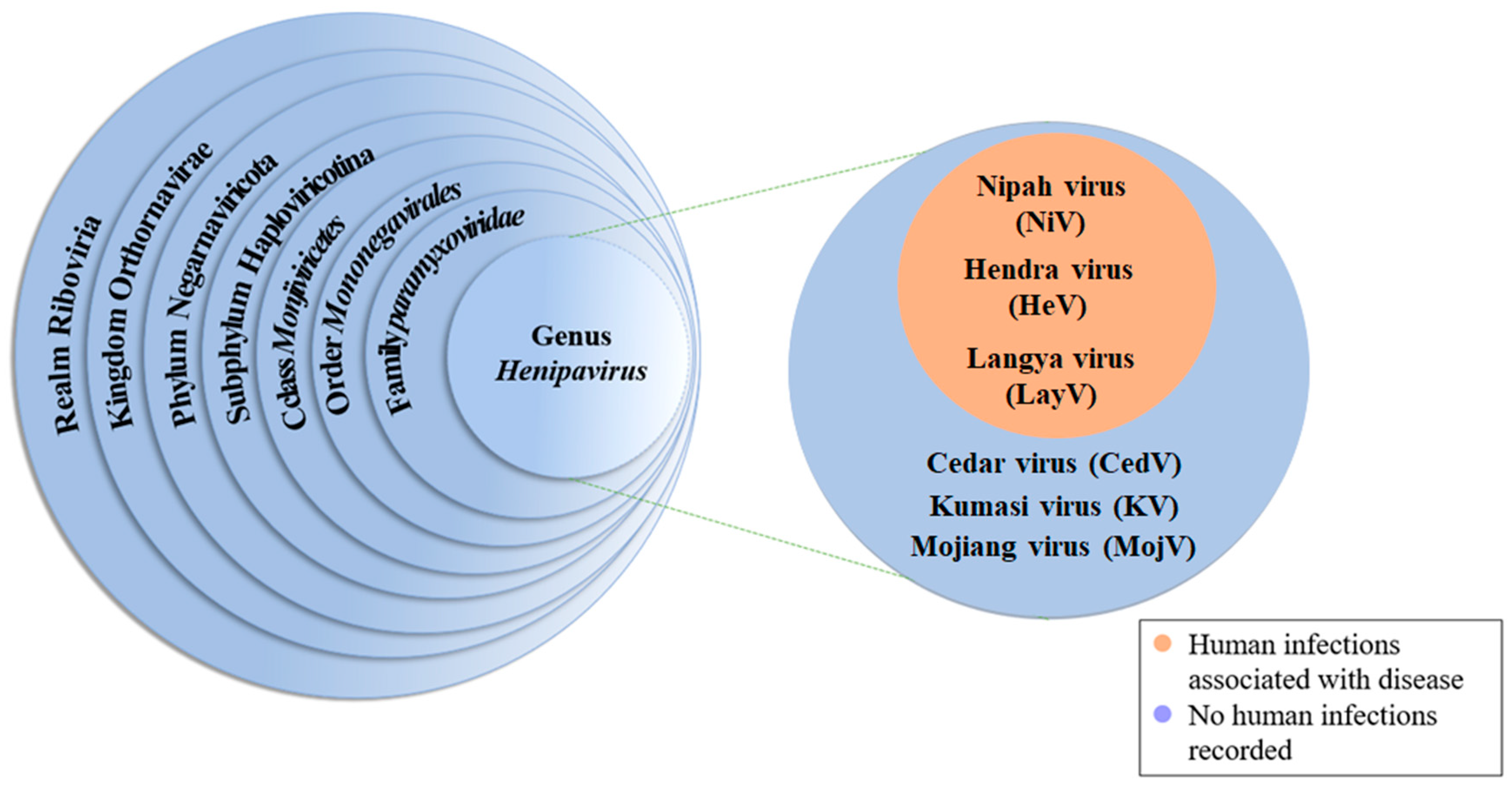
1. Introduction
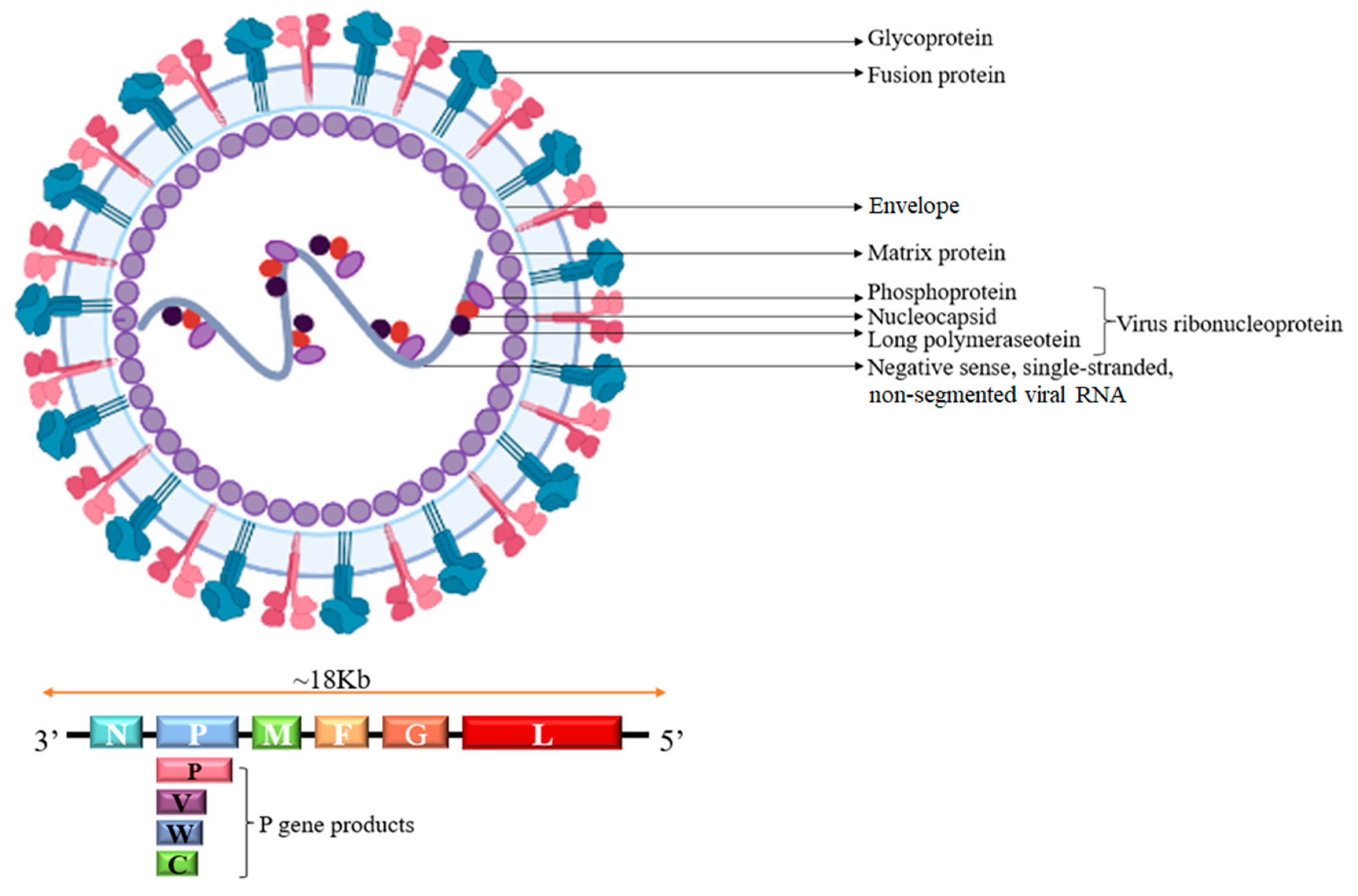
2. Biological Characteristics
3. The Structure of NiV
4. Animal Models
4.1. Golden Syrian Hamster Model
4.2. Ferret Model
4.3. African Green Monkey Model
4.4. Cavia Porcellus Model
4.5. Cat Model
4.6. Pig Model
4.7. Mouse Model

{kind=link}
{kind=link}
| Species | Age | Virus Strain | Dose | Route | Clinical Symptoms | Applications | References |
|---|---|---|---|---|---|---|---|
| Golden Syrian hamster | 2-month-old male | NiV-M | 1~104 pfu | IP | Tremor, limb paralysis | Antibody efficacy evaluation; pathogenesis study | [31,33] |
| Ferret | 1~2 years old | NiV-M |
5 × 102~5 × 104 TCID50 | IN&ON | Multisystem vasculitis, respiratory distress, neurological signs (rhinitis, encephalitis) | Monoclonal antibody therapy assessment | [34,36] |
| African green monkey | Adult, 5–7 kg | NiV-M | 2.5 × 103~1.3 × 106 pfu | IT&Oral | Acute respiratory distress, fever, neurological symptoms, multi-organ vasculitis | Gold standard for pathogenesis and vaccine durability studies | [40,41] |
| Cavia pocellus | 4-month-old male | NiV-M |
6 × 104~1 × 107 pfu | IP | Mild ataxia (IP); severe vasculitis and genital tract inflammation (high-dose IP) |
Virus excretion research; preliminary vaccine evaluation | [44,46] |
| Cat | 1~2 years old | NiV-M |
5 × 102.5~5 × 103 TCID50 | ON | Fever, increased respiratory rates, vomiting, decreased grooming, depression, dyspnea | Preliminary vaccine evaluation | [18] |
| Pig | 6-week-old female | NiV-M | 5 × 104 TCID50 | Oral&IH | Fever, respiratory distress, ataxia, exudative dermatitis, septic meningitis | Vaccine protection evaluation (e.g., F/G protein vaccine) | [18,47,49] |
| Mouse | 4-month-old male | NiV-M | 6 × 104 pfu | IP | Vasculitis, lymphocytic meningitis and inflammation ofthe genitourinary tract | Susceptibility assessment; pathologic studies; pathogenesis studies; antibody efficacy evaluation | [52] |
5. Vaccine Research Progress
5.1. Subunit Vaccines
5.2. Virus-like Particle Vaccines
5.3. Recombinant Virus Vaccines
5.4. Inactivated Vaccines
5.5. DNA Vaccines
5.6. mRNA Vaccine
5.7. Nanoparticle Vaccine
6. Treatment
6.1. Monoclonal Antibody
6.2. Favipiravir (T-705)
6.3. Ribavirin (Virazole)
7. Prevention and Control
8. Discussion
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lo, M.K.; Rota, P.A. The emergence of Nipah virus, a highly pathogenic paramyxovirus. J. Clin. Virol. 2008, 43, 396–400. [Google Scholar] [CrossRef] [PubMed]
- Epstein, J.H.; Anthony, S.J.; Islam, A.; Kilpatrick, A.M.; Khan, S.A.; Balkey, M.D.; Ross, N.; Smith, I.; Zambrana-Torrelio, C.; Tao, Y.; et al. Nipah virus dynamics in bats and implications for spillover to humans. Proc. Natl. Acad. Sci. USA 2020, 117, 29190–29201. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Deb, N.; Roy, P.; Jaiswal, V.; Sah, S.; Pandey, Y.; Edara, R.S.R.; Mohanty, A.; Henao-Martínez, A.F.; Sah, R. Recent Nipah virus outbreak in India: Lessons and imperatives. Ther. Adv. Infect. Dis. 2023, 10, 20499361231208535. [Google Scholar] [CrossRef]
- Singh, R.K.; Dhama, K.; Chakraborty, S.; Tiwari, R.; Natesan, S.; Khandia, R.; Munjal, A.; Vora, K.S.; Latheef, S.K.; Karthik, K.; et al. Nipah virus: Epidemiology, pathology, immunobiology and advances in diagnosis, vaccine designing and control strategies—A comprehensive review. Vet. Q. 2019, 39, 26–55. [Google Scholar] [CrossRef] [PubMed]
- Alam, A.M. Nipah virus, an emerging zoonotic disease causing fatal encephalitis. Clin. Med. 2022, 22, 348–352. [Google Scholar] [CrossRef]
- Chan, X.H.S.; Haeusler, I.L.; Choy, B.J.K.; Hassan, Z.; Takata, J.; Hurst, T.P.; Jones, L.M.; Loganathan, S.; Harriss, E.; Dunning, J.; et al. Therapeutics for Nipah virus disease: A systematic review to support prioritisation of drug candidates for clinical trials. Lancet Microbe 2025, 6, 101002. [Google Scholar] [CrossRef]
- Munster, V.J.; Prescott, J.B.; Bushmaker, T.; Long, D.; Rosenke, R.; Thomas, T.; Scott, D.; Fischer, E.R.; Feldmann, H.; de Wit, E. Rapid Nipah virus entry into the central nervous system of hamsters via the olfactory route. Sci. Rep. 2012, 2, 736. [Google Scholar] [CrossRef]
- Wong, K.T.; Shieh, W.J.; Kumar, S.; Norain, K.; Abdullah, W.; Guarner, J.; Goldsmith, C.S.; Chua, K.B.; Lam, S.K.; Tan, C.T.; et al. Nipah virus infection: Pathology and pathogenesis of an emerging paramyxoviral zoonosis. Am. J. Pathol. 2002, 161, 2153–2167. [Google Scholar] [CrossRef]
- Harit, A.K.; Ichhpujani, R.L.; Gupta, S.; Gill, K.S. Nipah/Hendra virus outbreak in Siliguri, West Bengal, India in 2001. Indian J. Med. Res. 2006, 123, 553–560. [Google Scholar]
- Thulaseedaran, N.K.; Kumar, K.G.S.; Kumar, J.; Geetha, P.; Jayachandran, N.V.; Kamalasanan, C.G.; Mathew, S.; Pv, S. A Case Series on the Recent Nipah Epidemic in Kerala. J. Assoc. Physicians India 2018, 66, 63–67. [Google Scholar]
- Hu, S.; Kim, H.; Yang, P.; Yu, Z.; Ludeke, B.; Mobilia, S.; Pan, J.; Stratton, M.; Bian, Y.; Fearns, R.; et al. Structural and functional analysis of the Nipah virus polymerase complex. Cell 2025, 188, 688–703.e18. [Google Scholar] [CrossRef] [PubMed]
- Satterfield, B.A.; Dawes, B.E.; Milligan, G.N. Status of vaccine research and development of vaccines for Nipah virus. Vaccine 2016, 34, 2971–2975. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Yao, Y.; Liu, H.; Zhang, X.; Li, X.; Liu, Y.; Peng, Y.; Chen, T.; Sun, Y.; Gao, G.; et al. Vaccines based on the fusion protein consensus sequence protect Syrian hamsters from Nipah virus infection. JCI Insight 2023, 8, e175461. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Cheng, R.; Yao, Y.; Zhang, G.; Li, X.; Wang, B.; Wang, Y.; Yu, F.; Yang, S.; Liu, H.; et al. An attachment glycoprotein nanoparticle elicits broadly neutralizing antibodies and protects against lethal Nipah virus infection. npj Vaccines 2024, 9, 158. [Google Scholar] [CrossRef]
- Nie, J.; Liu, L.; Wang, Q.; Chen, R.; Ning, T.; Liu, Q.; Huang, W.; Wang, Y. Nipah pseudovirus system enables evaluation of vaccines in vitro and in vivo using non-BSL-4 facilities. Emerg. Microbes Infect. 2019, 8, 272–281. [Google Scholar] [CrossRef]
- DeBuysscher, B.L.; Scott, D.; Marzi, A.; Prescott, J.; Feldmann, H. Single-dose live-attenuated Nipah virus vaccines confer complete protection by eliciting antibodies directed against surface glycoproteins. Vaccine 2014, 32, 2637–2644. [Google Scholar] [CrossRef]
- Lo, M.K.; Bird, B.H.; Chattopadhyay, A.; Drew, C.P.; Martin, B.E.; Coleman, J.D.; Rose, J.K.; Nichol, S.T.; Spiropoulou, C.F. Single-dose replication-defective VSV-based Nipah virus vaccines provide protection from lethal challenge in Syrian hamsters. Antivir. Res. 2014, 101, 26–29. [Google Scholar] [CrossRef]
- McEachern, J.A.; Bingham, J.; Crameri, G.; Green, D.J.; Hancock, T.J.; Middleton, D.; Feng, Y.-R.; Broder, C.C.; Wang, L.-F.; Bossart, K.N. A recombinant subunit vaccine formulation protects against lethal Nipah virus challenge in cats. Vaccine 2008, 26, 3842–3852. [Google Scholar] [CrossRef]
- Mire, C.E.; Versteeg, K.M.; Cross, R.W.; Agans, K.N.; Fenton, K.A.; Whitt, M.A.; Geisbert, T.W. Single injection recombinant vesicular stomatitis virus vaccines protect ferrets against lethal Nipah virus disease. Virol. J. 2013, 10, 353. [Google Scholar] [CrossRef]
- Bossart, K.N.; Rockx, B.; Feldmann, F.; Brining, D.; Scott, D.; LaCasse, R.; Geisbert, J.B.; Feng, Y.-R.; Chan, Y.-P.; Hickey, A.C.; et al. A Hendra virus G glycoprotein subunit vaccine protects African green monkeys from Nipah virus challenge. Sci. Transl. Med. 2012, 4, 146ra107. [Google Scholar] [CrossRef]
- Prescott, J.; DeBuysscher, B.L.; Feldmann, F.; Gardner, D.J.; Haddock, E.; Martellaro, C.; Scott, D.; Feldmann, H. Single-dose live-attenuated vesicular stomatitis virus-based vaccine protects African green monkeys from Nipah virus disease. Vaccine 2015, 33, 2823–2829. [Google Scholar] [CrossRef] [PubMed]
- Weingartl, H.M.; Berhane, Y.; Caswell, J.L.; Loosmore, S.; Audonnet, J.-C.; Roth, J.A.; Czub, M. Recombinant nipah virus vaccines protect pigs against challenge. J. Virol. 2006, 80, 7929–7938. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Yao, Y.; Zhang, X.; Liu, H.; Gao, G.; Peng, Y.; Chen, M.; Zhao, J.; Zhang, X.; Yin, C.; et al. Both chimpanzee adenovirus-vectored and DNA vaccines induced long-term immunity against Nipah virus infection. npj Vaccines 2023, 8, 170. [Google Scholar] [CrossRef] [PubMed]
- Harcourt, B.H.; Tamin, A.; Ksiazek, T.G.; Rollin, P.E.; Anderson, L.J.; Bellini, W.J.; Rota, P.A. Molecular characterization of Nipah virus, a newly emergent paramyxovirus. Virology 2000, 271, 334–349. [Google Scholar] [CrossRef]
- Zhang, X.A.; Li, H.; Jiang, F.C.; Zhu, F.; Zhang, Y.-F.; Chen, J.-J.; Tan, C.-W.; Anderson, D.E.; Fan, H.; Dong, L.-Y.; et al. A Zoonotic Henipavirus in Febrile Patients in China. N. Engl. J. Med. 2022, 387, 470–472. [Google Scholar] [CrossRef]
- Soman Pillai, V.; Krishna, G.; Valiya Veettil, M. Nipah Virus: Past Outbreaks and Future Containment. Viruses 2020, 12, 465. [Google Scholar] [CrossRef]
- Sharma, V.; Kaushik, S.; Kumar, R.; Yadav, J.P.; Kaushik, S. Emerging trends of Nipah virus: A review. Rev. Med. Virol. 2019, 29, e2010. [Google Scholar] [CrossRef]
- Hino, K.; Sato, H.; Sugai, A.; Kato, M.; Yoneda, M.; Kai, C. Downregulation of Nipah virus N mRNA occurs through interaction between its 3′ untranslated region and hnRNP D. J. Virol. 2013, 87, 6582–6588. [Google Scholar] [CrossRef]
- Chan, Y.P.; Chua, K.B.; Koh, C.L.; Lim, M.E.; Lam, S.K. Complete nucleotide sequences of Nipah virus isolates from Malaysia. J. Gen. Virol. 2001, 82, 2151–2155. [Google Scholar] [CrossRef]
- Omi-Furutani, M.; Yoneda, M.; Fujita, K.; Ikeda, F.; Kai, C. Novel phosphoprotein-interacting region in Nipah virus nucleocapsid protein and its involvement in viral replication. J. Virol. 2010, 84, 9793–9799. [Google Scholar] [CrossRef]
- Shaw, M.L.; García-Sastre, A.; Palese, P.; Basler, C.F. Nipah virus V and W proteins have a common STAT1-binding domain yet inhibit STAT1 activation from the cytoplasmic and nuclear compartments, respectively. J. Virol. 2004, 78, 5633–5641. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Chen, L.; Aguilar, H.C.; Chou, K.C. A stochastic assembly model for Nipah virus revealed by super-resolution microscopy. Nat. Commun. 2018, 9, 3050. [Google Scholar] [CrossRef] [PubMed]
- Wickenhagen, A.; van Tol, S.; Munster, V. Molecular determinants of cross-species transmission in emerging viral infections. Microbiol. Mol. Biol. Rev. 2024, 88, e0000123. [Google Scholar] [CrossRef] [PubMed]
- Geisbert, T.W.; Bobb, K.; Borisevich, V.; Geisbert, J.B.; Agans, K.N.; Cross, R.W.; Prasad, A.N.; Fenton, K.A.; Yu, H.; Fouts, T.R.; et al. A single dose investigational subunit vaccine for human use against Nipah virus and Hendra virus. npj Vaccines 2021, 6, 23. [Google Scholar] [CrossRef]
- Wong, K.T.; Grosjean, I.; Brisson, C.; Blanquier, B.; Fevre-Montange, M.; Bernard, A.; Loth, P.; Georges-Courbot, M.-C.; Chevallier, M.; Akaoka, H.; et al. A golden hamster model for human acute Nipah virus infection. Am. J. Pathol. 2003, 163, 2127–2137. [Google Scholar] [CrossRef]
- de Wit, E.; Bushmaker, T.; Scott, D.; Feldmann, H.; Munster, V.J. Nipah virus transmission in a hamster model. PLoS Negl. Trop. Dis. 2011, 5, e1432. [Google Scholar] [CrossRef]
- Guillaume, V.; Wong, K.T.; Looi, R.Y.; Georges-Courbot, M.-C.; Barrot, L.; Buckland, R.; Wild, T.F.; Horvat, B. Acute Hendra virus infection: Analysis of the pathogenesis and passive antibody protection in the hamster model. Virology 2009, 387, 459–465. [Google Scholar] [CrossRef]
- Bossart, K.N.; Zhu, Z.; Middleton, D.; Klippel, J.; Crameri, G.; Bingham, J.; McEachern, J.A.; Green, D.; Hancock, T.J.; Chan, Y.-P.; et al. A neutralizing human monoclonal antibody protects against lethal disease in a new ferret model of acute nipah virus infection. PLoS Pathog. 2009, 5, e1000642. [Google Scholar] [CrossRef]
- Geisbert, T.W.; Daddario-DiCaprio, K.M.; Hickey, A.C.; Smith, M.A.; Chan, Y.-P.; Wang, L.-F.; Mattapallil, J.J.; Geisbert, J.B.; Bossart, K.N.; Broder, C.C. Development of an acute and highly pathogenic nonhuman primate model of Nipah virus infection. PLoS ONE 2010, 5, e10690. [Google Scholar] [CrossRef]
- Geisbert, T.W.; Feldmann, H.; Broder, C.C. Animal challenge models of henipavirus infection and pathogenesis. Curr. Top. Microbiol. Immunol. 2012, 359, 153–177. [Google Scholar]
- Lawrence, P.; Escudero-Pérez, B. Henipavirus Immune Evasion and Pathogenesis Mechanisms: Lessons Learnt from Natural Infection and Animal Models. Viruses 2022, 14, 936. [Google Scholar] [CrossRef] [PubMed]
- Woolsey, C.; Borisevich, V.; Fears, A.C.; Agans, K.N.; Deer, D.J.; Prasad, A.N.; O’toole, R.; Foster, S.L.; Dobias, N.S.; Geisbert, J.B.; et al. Recombinant vesicular stomatitis virus-vectored vaccine induces long-lasting immunity against Nipah virus disease. J. Clin. Investig. 2023, 133, e164946. [Google Scholar] [CrossRef] [PubMed]
- Maisner, A.; Neufeld, J.; Weingartl, H. Organ- and endotheliotropism of Nipah virus infections in vivo and in vitro. Thromb. Haemost. 2009, 102, 1014–1023. [Google Scholar]
- Torres-Velez, F.J.; Shieh, W.J.; Rollin, P.E.; Morken, T.; Brown, C.; Ksiazek, T.G.; Zaki, S.R. Histopathologic and immunohistochemical characterization of Nipah virus infection in the guinea pig. Vet. Pathol. 2008, 45, 576–585. [Google Scholar] [CrossRef]
- Weingartl, H.M.; Berhane, Y.; Czub, M. Animal models of henipavirus infection: A review. Vet. J. 2009, 181, 211–220. [Google Scholar] [CrossRef]
- Middleton, D.J.; Morrissy, C.J.; van der Heide, B.M.; Russell, G.M.; Braun, M.A.; Westbury, H.A.; Halpin, K.; Daniels, P.W. Experimental Nipah virus infection in pteropid bats (Pteropus poliocephalus). J. Comp. Pathol. 2007, 136, 266–272. [Google Scholar] [CrossRef]
- Chua, K.B. Nipah virus outbreak in Malaysia. J. Clin. Virol. 2003, 26, 265–275. [Google Scholar] [CrossRef]
- Middleton, D.J.; Westbury, H.A.; Morrissy, C.J.; van der Heide, B.; Russell, G.; Braun, M.; Hyatt, A. Experimental Nipah virus infection in pigs and cats. J. Comp. Pathol. 2002, 126, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Mungall, B.A.; Middleton, D.; Crameri, G.; Halpin, K.; Bingham, J.; Eaton, B.T.; Broder, C.C. Vertical transmission and fetal replication of Nipah virus in an experimentally infected cat. J. Infect. Dis. 2007, 196, 812–816. [Google Scholar] [CrossRef]
- Wong, K.T.; Shieh, W.J.; Zaki, S.R.; Tan, C.T. Nipah virus infection, an emerging paramyxoviral zoonosis. Springer Semin. Immunopathol. 2002, 24, 215–228. [Google Scholar] [CrossRef]
- Berhane, Y.; Weingartl, H.M.; Lopez, J.; Neufeld, J.; Czub, S.; Embury-Hyatt, C.; Goolia, M.; Copps, J.; Czub, M. Bacterial infections in pigs experimentally infected with Nipah virus. Transbound. Emerg. Dis. 2008, 55, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Spengler, J.R.; Lo, M.K.; Welch, S.R.; Spiropoulou, C.F. Henipaviruses: Epidemiology, ecology, disease, and the development of vaccines and therapeutics. Clin. Microbiol. Rev. 2025, 38, e0012823. [Google Scholar] [CrossRef] [PubMed]
- Rodrigue, V.; Gravagna, K.; Yao, J.; Nafade, V.; Basta, N.E. Current progress towards prevention of Nipah and Hendra disease in humans: A scoping review of vaccine and monoclonal antibody candidates being evaluated in clinical trials. Trop. Med. Int. Health 2024, 29, 354–364. [Google Scholar] [CrossRef]
- Van Herck, S.; Feng, B.; Tang, L. Delivery of STING agonists for adjuvanting subunit vaccines. Adv. Drug Deliv. Rev. 2021, 179, 114020. [Google Scholar] [CrossRef]
- Broder, C.C. Passive immunization and active vaccination against Hendra and Nipah viruses. Dev. Biol. 2013, 135, 125–138. [Google Scholar]
- Pallister, J.A.; Klein, R.; Arkinstall, R.; Haining, J.; Long, F.; White, J.R.; Payne, J.; Feng, Y.-R.; Wang, L.-F.; Broder, C.C.; et al. Vaccination of ferrets with a recombinant G glycoprotein subunit vaccine provides protection against Nipah virus disease for over 12 months. Virol. J. 2013, 10, 237. [Google Scholar] [CrossRef]
- Roldão, A.; Mellado, M.C.; Castilho, L.R.; Carrondo, M.J.; Alves, P.M. Virus-like particles in vaccine development. Expert Rev. Vaccines 2010, 9, 1149–1176. [Google Scholar] [CrossRef]
- Walpita, P.; Barr, J.; Sherman, M.; Basler, C.F.; Wang, L. Vaccine potential of Nipah virus-like particles. PLoS ONE 2011, 6, e18437. [Google Scholar] [CrossRef]
- Walpita, P.; Cong, Y.; Jahrling, P.B.; Rojas, O.; Postnikova, E.; Yu, S.; Johns, L.; Holbrook, M.R. A VLP-based vaccine provides complete protection against Nipah virus challenge following multiple-dose or single-dose vaccination schedules in a hamster model. npj Vaccines 2017, 2, 21. [Google Scholar] [CrossRef]
- Mire, C.E.; Geisbert, J.B.; Agans, K.N.; Versteeg, K.M.; Deer, D.J.; Satterfield, B.A.; Fenton, K.A.; Geisbert, T.W. Use of Single-Injection Recombinant Vesicular Stomatitis Virus Vaccine to Protect Nonhuman Primates Against Lethal Nipah Virus Disease. Emerg. Infect. Dis. 2019, 25, 1144–1152. [Google Scholar] [CrossRef]
- Gomme, E.A.; Wanjalla, C.N.; Wirblich, C.; Schnell, M.J. Rabies virus as a research tool and viral vaccine vector. Adv. Virus Res. 2011, 79, 139–164. [Google Scholar] [PubMed]
- Keshwara, R.; Shiels, T.; Postnikova, E.; Kurup, D.; Wirblich, C.; Johnson, R.F.; Schnell, M.J. Erratum: Publisher Correction: Rabies-based vaccine induces potent immune responses against Nipah virus. npj Vaccines 2019, 4, 18. [Google Scholar] [CrossRef] [PubMed]
- Ploquin, A.; Szécsi, J.; Mathieu, C.; Guillaume, V.; Barateau, V.; Ong, K.C.; Wong, K.T.; Cosset, F.-L.; Horvat, B.; Salvetti, A. Protection against henipavirus infection by use of recombinant adeno-associated virus-vector vaccines. J. Infect. Dis. 2013, 207, 469–478. [Google Scholar] [CrossRef] [PubMed]
- van Doremalen, N.; Lambe, T.; Sebastian, S.; Bushmaker, T.; Fischer, R.; Feldmann, F.; Haddock, E.; Letko, M.; Avanzato, V.A.; Rissanen, I.; et al. A single-dose ChAdOx1-vectored vaccine provides complete protection against Nipah Bangladesh and Malaysia in Syrian golden hamsters. PLoS Negl. Trop. Dis. 2019, 13, e0007462. [Google Scholar] [CrossRef]
- Guillaume, V.; Contamin, H.; Loth, P.; Georges-Courbot, M.-C.; Lefeuvre, A.; Marianneau, P.; Chua, K.B.; Lam, S.K.; Buckland, R.; Deubel, V.; et al. Nipah virus: Vaccination and passive protection studies in a hamster model. J. Virol. 2004, 78, 834–840. [Google Scholar] [CrossRef]
- Blackburn, N.K.; Besselaar, T.G. A study of the effect of chemical inactivants on the epitopes of Rift Valley fever virus glycoproteins using monoclonal antibodies. J. Virol. Methods 1991, 33, 367–374. [Google Scholar] [CrossRef]
- Buonavoglia, C.; Fioretti, A.; Tollis, M.; Menna, F.; Papparella, V. A preliminary vaccine potency trial of a Newcastle disease virus inactivated with binary ethylenimine. Vet. Res. Commun. 1988, 12, 195–197. [Google Scholar] [CrossRef]
- Wang, X.; Ge, J.; Hu, S.; Wang, Q.; Wen, Z.; Chen, H.; Bu, Z. Efficacy of DNA immunization with F and G protein genes of Nipah virus. Ann. N. Y. Acad. Sci. 2006, 1081, 243–245. [Google Scholar] [CrossRef]
- Loomis, R.J.; DiPiazza, A.T.; Falcone, S.; Ruckwardt, T.J.; Morabito, K.M.; Abiona, O.M.; Chang, L.A.; Caringal, R.T.; Presnyak, V.; Narayanan, E.; et al. Chimeric Fusion (F) and Attachment (G) Glycoprotein Antigen Delivery by mRNA as a Candidate Nipah Vaccine. Front. Immunol. 2021, 12, 772864. [Google Scholar] [CrossRef]
- Rangacharya, O.; Parab, A.; Adkine, S.; Nagargoje, R. A study on the design of an in silico self-amplifying mRNA vaccine against Nipah virus using immunoinformatics. J. Biomol. Struct. Dyn. 2023, 41, 12777–12788. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, X.; Yao, Y.; Wang, S.; Li, K.; Zhang, B.; Ye, T.; Chen, L.; Wu, Y.; Li, E.; et al. Ferritin nanoparticle-based Nipah virus glycoprotein vaccines elicit potent protective immune responses in mice and hamsters. Virol. Sin. 2024, 39, 909–916. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Tang, D.; Atluri, S. Patient-Specific Carotid Plaque Progression Simulation Using 3D Meshless Generalized Finite Difference Models with Fluid-Structure Interactions Based on Serial In Vivo MRI Data. Comput. Model. Eng. Sci. 2011, 72, 53–77. [Google Scholar] [PubMed]
- Playford, E.G.; Munro, T.; Mahler, S.M.; Elliott, S.; Gerometta, M.; Hoger, K.L.; Jones, M.L.; Griffin, P.; Lynch, K.D.; Carroll, H.; et al. Safety, tolerability, pharmacokinetics, and immunogenicity of a human monoclonal antibody targeting the G glycoprotein of henipaviruses in healthy adults: A first-in-human, randomised, controlled, phase 1 study. Lancet Infect. Dis. 2020, 20, 445–454. [Google Scholar] [CrossRef]
- Sen, N.; Kanitkar, T.R.; Roy, A.A.; Soni, N.; Amritkar, K.; Supekar, S.; Nair, S.; Singh, G.; Madhusudhan, M.S. Predicting and designing therapeutics against the Nipah virus. PLoS Negl. Trop. Dis. 2019, 13, e0007419. [Google Scholar] [CrossRef] [PubMed]
- Clayton, B.A.; Middleton, D.; Bergfeld, J.; Haining, J.; Arkinstall, R.; Wang, L.; Marsh, G.A. Transmission routes for nipah virus from Malaysia and Bangladesh. Emerg. Infect. Dis. 2012, 18, 1983–1993. [Google Scholar] [CrossRef]
- Doyle, M.P.; Kose, N.; Borisevich, V.; Binshtein, E.; Amaya, M.; Nagel, M.; Annand, E.J.; Armstrong, E.; Bombardi, R.; Dong, J.; et al. Cooperativity mediated by rationally selected combinations of human monoclonal antibodies targeting the henipavirus receptor binding protein. Cell Rep. 2021, 36, 109628. [Google Scholar] [CrossRef]
- Dawes, B.E.; Kalveram, B.; Ikegami, T.; Juelich, T.; Smith, J.K.; Zhang, L.; Park, A.; Lee, B.; Komeno, T.; Furuta, Y.; et al. Favipiravir (T-705) protects against Nipah virus infection in the hamster model. Sci. Rep. 2018, 8, 7604. [Google Scholar] [CrossRef]
- Broder, C.C. Henipavirus outbreaks to antivirals: The current status of potential therapeutics. Curr. Opin. Virol. 2012, 2, 176–187. [Google Scholar] [CrossRef]
- Paton, N.I.; Leo, Y.S.; Zaki, S.R.; Auchus, A.P.; Lee, K.E.; Ling, A.E.; Chew, S.K.; Ang, B.; Rollin, P.E.; Umapathi, T.; et al. Outbreak of Nipah-virus infection among abattoir workers in Singapore. Lancet 1999, 354, 1253–1256. [Google Scholar] [CrossRef]
- Chong, H.T.; Kamarulzaman, A.; Tan, C.T.; Goh, K.-J.; Thayaparan, T.; Kunjapan, S.R.; Chew, N.-K.; Chua, K.-B.; Lam, S.-K. Treatment of acute Nipah encephalitis with ribavirin. Ann. Neurol. 2001, 49, 810–813. [Google Scholar] [CrossRef]
- Freiberg, A.N.; Worthy, M.N.; Lee, B.; Holbrook, M.R. Combined chloroquine and ribavirin treatment does not prevent death in a hamster model of Nipah and Hendra virus infection. J. Gen. Virol. 2010, 91, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.Z.; Sazzad, H.M.S.; Luby, S.P.; Sturm-Ramirez, K.; Bhuiyan, M.U.; Rahman, M.Z.; Islam, M.M.; Ströher, U.; Sultana, S.; Kafi, M.A.H.; et al. Nipah Virus Contamination of Hospital Surfaces during Outbreaks, Bangladesh, 2013–2014. Emerg. Infect. Dis. 2018, 24, 15–21. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, C.; He, J.; Zhang, Q.; Wang, T. Vaccines and Animal Models of Nipah Virus: Current Situation and Future Prospects. Vaccines 2025, 13, 608. https://doi.org/10.3390/vaccines13060608
Lv C, He J, Zhang Q, Wang T. Vaccines and Animal Models of Nipah Virus: Current Situation and Future Prospects. Vaccines. 2025; 13(6):608. https://doi.org/10.3390/vaccines13060608
Chicago/Turabian StyleLv, Chaoxiang, Jiayue He, Qiqi Zhang, and Tiecheng Wang. 2025. "Vaccines and Animal Models of Nipah Virus: Current Situation and Future Prospects" Vaccines 13, no. 6: 608. https://doi.org/10.3390/vaccines13060608
APA StyleLv, C., He, J., Zhang, Q., & Wang, T. (2025). Vaccines and Animal Models of Nipah Virus: Current Situation and Future Prospects. Vaccines, 13(6), 608. https://doi.org/10.3390/vaccines13060608