Comparative Analysis of Long-Term Measles Immune Response After Natural Infection and Routine Vaccination in China

 ,
,  ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
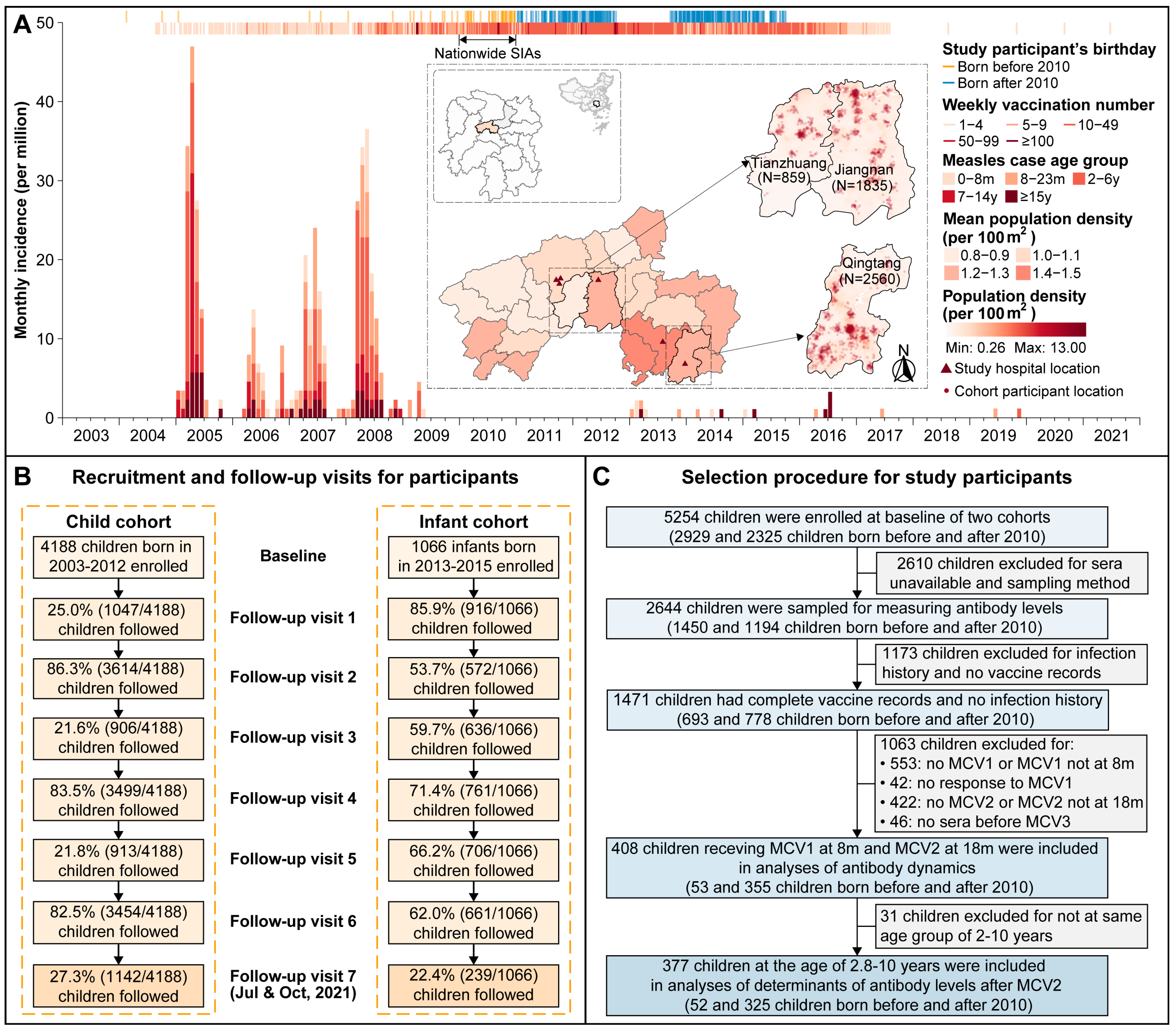
2.1. Study Population and Data Collection
2.2. Laboratory Procedures
2.3. Statistical Analyses
3. Results
3.1. Basic Characteristics of Study Participants
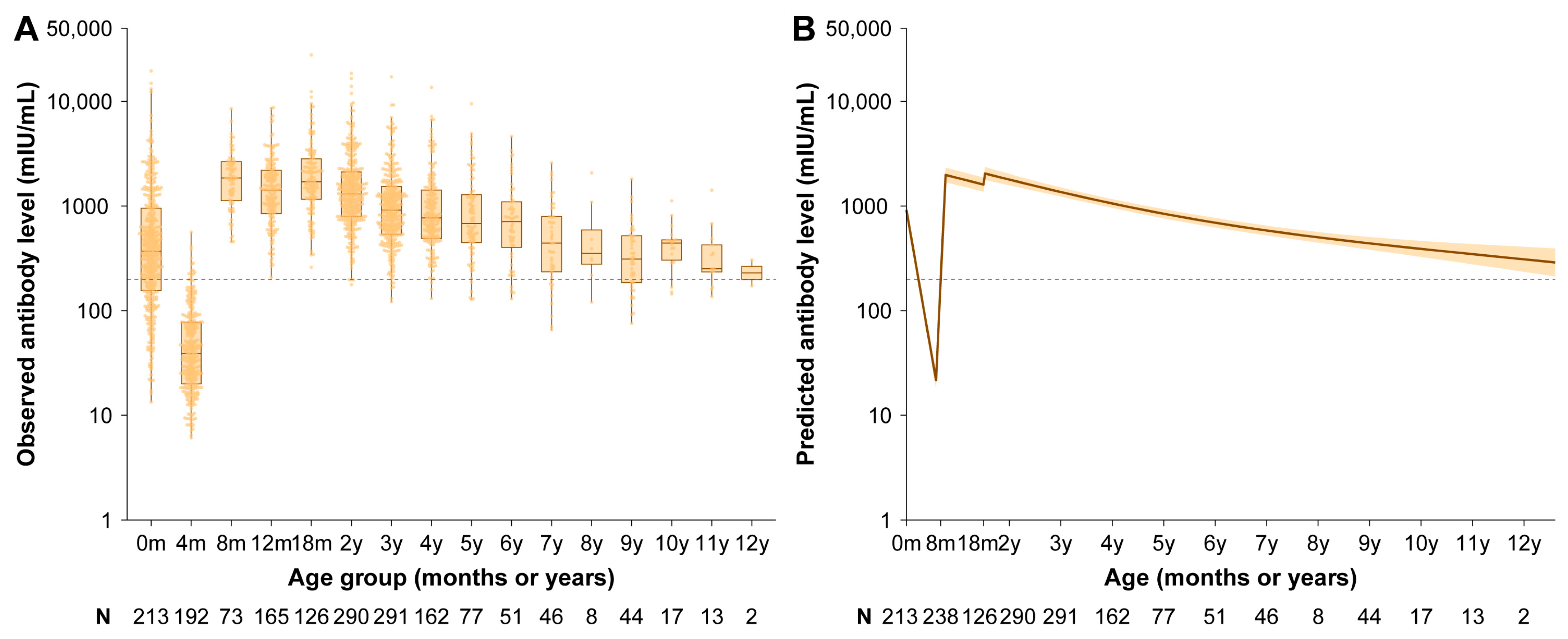
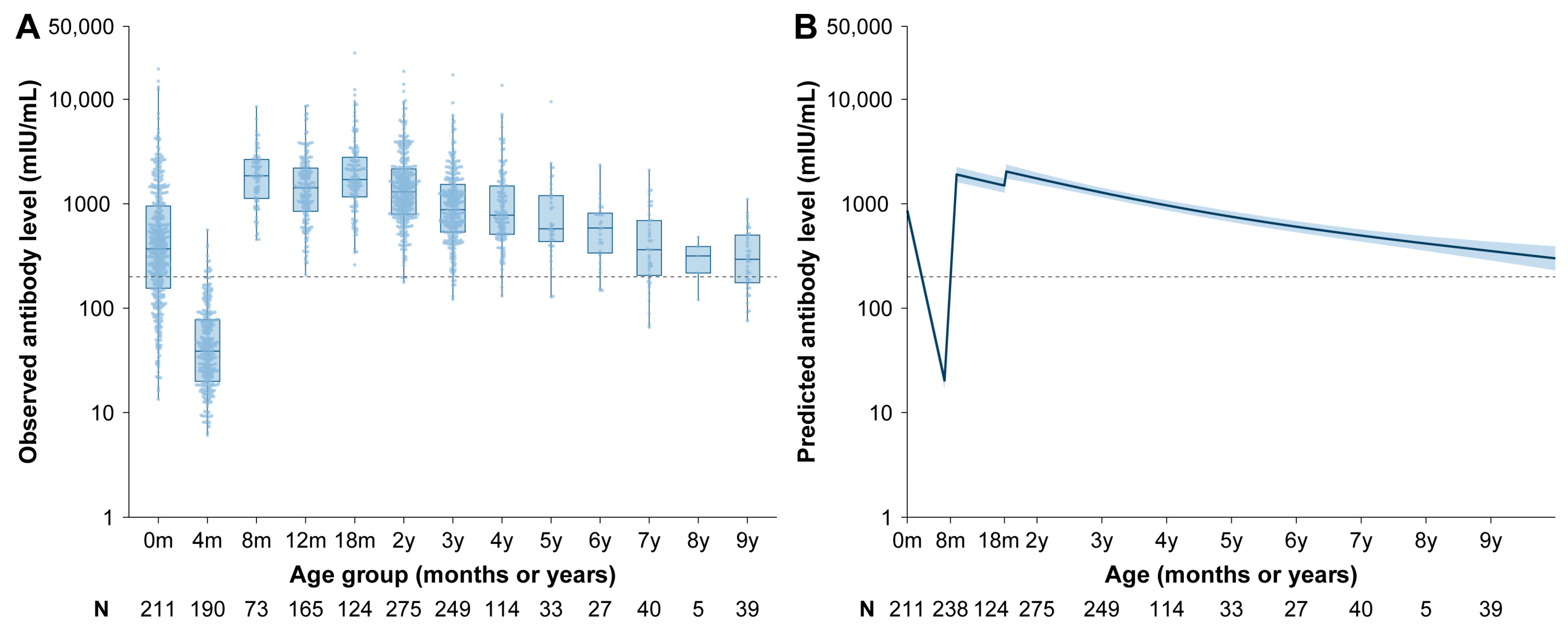
3.2. Vaccine-Induced Antibody
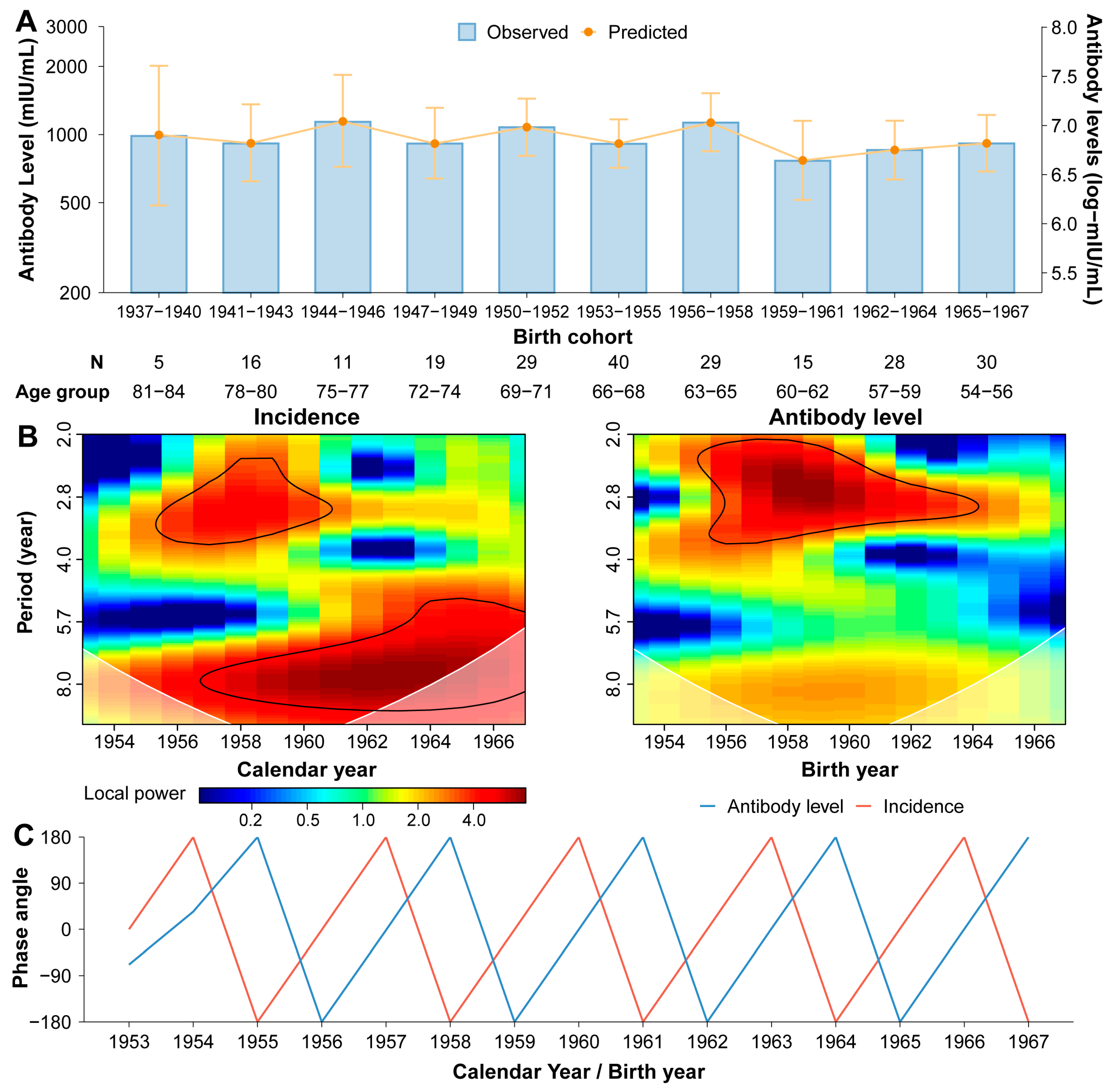
3.3. Naturally Acquired Antibody
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| MCV | measles-containing vaccine |
| MV | monovalent measles vaccine |
| MR | measles-rubella combined vaccine |
| MMR | measles-mumps-rubella combined vaccine |
| SIA | supplementary immunization activity |
| ELISA | enzyme-linked immunosorbent assay |
| GAMM | generalized additive mixed model |
| CI | confidence interval |
References
- Moss, W.J. Measles. Lancet 2017, 390, 2490–2502. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; An, Z.; Yin, Z. Achievements in prevention and control of seven infectious diseases targeted by the National Immunization Program in China across 70 years. Chin. J. Vaccines Immun. 2019, 25, 359–367. [Google Scholar]
- Trentini, F.; Poletti, P.; Merler, S.; Melegaro, A. Measles Immunity Gaps and the Progress towards Elimination: A Multi-Country Modelling Analysis. Lancet Infect. Dis. 2017, 17, 1089–1097. [Google Scholar] [CrossRef]
- Ma, C. Progress Toward Measles Elimination—China, January 2013–June 2019. MMWR Morb. Mortal. Wkly. Rep. 2019, 68, 1112–1116. [Google Scholar] [CrossRef] [PubMed]
- Leung, J.; Munir, N.A.; Mathis, A.D.; Filardo, T.D.; Rota, P.A.; Sugerman, D.E.; Sowers, S.B.; Mercader, S.; Crooke, S.N.; Gastañaduy, P.A. The Effects of Vaccination Status and Age on Clinical Characteristics and Severity of Measles Cases in the United States in the Post-Elimination Era, 2001–2022. Clin. Infect. Dis. 2024, 80, 663–672. [Google Scholar] [CrossRef]
- World Health Organization. Measles Cases Surge Worldwide, Infecting 10.3 Million People in 2023. Available online: https://www.who.int/news/item/14-11-2024-measles-cases-surge-worldwide--infecting-10.3-million-people-in-2023 (accessed on 15 November 2024).
- Winter, A.K.; Martinez, M.E.; Cutts, F.T.; Moss, W.J.; Ferrari, M.J.; McKee, A.; Lessler, J.; Hayford, K.; Wallinga, J.; Metcalf, C.J.E. Benefits and Challenges in Using Seroprevalence Data to Inform Models for Measles and Rubella Elimination. J. Infect. Dis. 2018, 218, 355–364. [Google Scholar] [CrossRef]
- Amanna, I.J.; Carlson, N.E.; Slifka, M.K. Duration of Humoral Immunity to Common Viral and Vaccine Antigens. N. Engl. J. Med. 2007, 357, 1903–1915. [Google Scholar] [CrossRef]
- World Health Organization. The Immunological Basis for Immunization Series: Module 4: Pertussis; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- World Health Organization. The Immunological Basis for Immunization Series: Module 23: Influenza Vaccines; World Health Organization: Geneva, Switzerland, 2017; ISBN 978-92-4-151305-0. [Google Scholar]
- Wei, X.; Yang, J.; Gao, L.; Wang, L.; Liao, Q.; Qiu, Q.; Luo, K.; Yu, S.; Zhou, Y.; Liu, F.; et al. The Transfer and Decay of Maternal Antibodies against Enterovirus A71, and Dynamics of Antibodies Due to Later Natural Infections in Chinese Infants: A Longitudinal, Paired Mother–Neonate Cohort Study. Lancet Infect. Dis. 2021, 21, 418–426. [Google Scholar] [CrossRef]
- Wang, W.; O’Driscoll, M.; Wang, Q.; Zhao, S.; Salje, H.; Yu, H. Dynamics of Measles Immunity from Birth and Following Vaccination. Nat. Microbiol. 2024, 9, 1676–1685. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, W.; Winter, A.K.; Zhan, Z.; Ajelli, M.; Trentini, F.; Wang, L.; Li, F.; Yang, J.; Xiang, X.; et al. Long-Term Measles Antibody Profiles Following Different Vaccine Schedules in China, a Longitudinal Study. Nat. Commun. 2023, 14, 1746. [Google Scholar] [CrossRef]
- Chen, R.T.; Markowitz, L.E.; Albrecht, P.; Stewart, J.A.; Mofenson, L.M.; Preblud, S.R.; Orenstein, W.A. Measles Antibody: Reevaluation of Protective Titers. J. Infect. Dis. 1990, 162, 1036–1042. [Google Scholar] [CrossRef]
- Lauri, E.M.; Jaime, S.; Jose, L.D.-O.; Jose, L.V.; Paul, A.; Elizabeth, R.Z.; John, S.; Maria, L.Z.; Roger, H.B. Immunization of Six-Month-Old Infants with Different Doses of Edmonston–Zagreb and Schwarz Measles Vaccines. N. Engl. J. Med. 1990, 322, 580–587. [Google Scholar] [CrossRef]
- Lam, H.M.; Phuong, H.T.; Thao Vy, N.H.; Le Thanh, N.T.; Dung, P.N.; Ngoc Muon, T.T.; Van Vinh Chau, N.; Rodríguez-Barraquer, I.; Cummings, D.A.T.; Wills, B.A.; et al. Serological Inference of Past Primary and Secondary Dengue Infection: Implications for Vaccination. J. R. Soc. Interface 2019, 16, 20190207. [Google Scholar] [CrossRef]
- Grenfell, B.T.; Bjørnstad, O.N.; Kappey, J. Travelling Waves and Spatial Hierarchies in Measles Epidemics. Nature 2001, 414, 716–723. [Google Scholar] [CrossRef]
- Torrence, C.; Compo, G.P. A Practical Guide to Wavelet Analysis. Bull. Am. Meteorol. Soc. 1998, 79, 61–78. [Google Scholar] [CrossRef]
- Yang, L.; Grenfell, B.T.; Mina, M.J. Waning Immunity and Re-Emergence of Measles and Mumps in the Vaccine Era. Curr. Opin. Virol. 2020, 40, 48–54. [Google Scholar] [CrossRef]
- Fischinger, S.; Boudreau, C.M.; Butler, A.L.; Streeck, H.; Alter, G. Sex Differences in Vaccine-Induced Humoral Immunity. Semin. Immunopathol. 2019, 41, 239–249. [Google Scholar] [CrossRef]
- Bolotin, S.; Osman, S.; Hughes, S.L.; Ariyarajah, A.; Tricco, A.C.; Khan, S.; Li, L.; Johnson, C.; Friedman, L.; Gul, N.; et al. In Elimination Settings, Measles Antibodies Wane After Vaccination but Not After Infection: A Systematic Review and Meta-Analysis. J. Infect. Dis. 2022, 226, 1127–1139. [Google Scholar] [CrossRef] [PubMed]
- Mossong, J.; Nokes, D.J.; Edmunds, W.J.; Cox, M.J.; Ratnam, S.; Muller, C.P. Modeling the Impact of Subclinical Measles Transmission in Vaccinated Populations with Waning Immunity. Am. J. Epidemiol. 1999, 150, 1238–1249. [Google Scholar] [CrossRef]
- Ozanne, G.; d’Halewyn, M.A. Secondary Immune Response in a Vaccinated Population during a Large Measles Epidemic. J. Clin. Microbiol. 1992, 30, 1778–1782. [Google Scholar] [CrossRef] [PubMed]
- Dai, B.; Chen, Z.H.; Liu, Q.C.; Wu, T.; Guo, C.Y.; Wang, X.Z.; Fang, H.H.; Xiang, Y.Z. Duration of Immunity Following Immunization with Live Measles Vaccine: 15 Years of Observation in Zhejiang Province, China. Bull. World Health Organ. 1991, 69, 415–423. [Google Scholar] [PubMed]
- Borremans, B.; Mummah, R.O.; Guglielmino, A.H.; Galloway, R.L.; Hens, N.; Prager, K.C.; Lloyd-Smith, J.O. Inferring Time of Infection from Field Data Using Dynamic Models of Antibody Decay. Methods Ecol. Evol. 2023, 14, 2654–2667. [Google Scholar] [CrossRef]
- Salje, H.; Cummings, D.A.T.; Rodriguez-Barraquer, I.; Katzelnick, L.C.; Lessler, J.; Klungthong, C.; Thaisomboonsuk, B.; Nisalak, A.; Weg, A.; Ellison, D.; et al. Reconstruction of Antibody Dynamics and Infection Histories to Evaluate Dengue Risk. Nature 2018, 557, 719–723. [Google Scholar] [CrossRef]
- Russell, T.W.; Townsley, H.; Hellewell, J.; Gahir, J.; Shawe-Taylor, M.; Greenwood, D.; Hodgson, D.; Hobbs, A.; Dowgier, G.; Penn, R.; et al. Real-Time Estimation of Immunological Responses against Emerging SARS-CoV-2 Variants in the UK: A Mathematical Modelling Study. Lancet Infect. Dis. 2024, 25, 80–93. [Google Scholar] [CrossRef]
- Gustafson, T.L.; Lievens, A.W.; Brunell, P.A.; Moellenberg, R.G.; Buttery, C.M.; Sehulster, L.M. Measles Outbreak in a Fully Immunized Secondary-School Population. N. Engl. J. Med. 1987, 316, 771–774. [Google Scholar] [CrossRef]
- Wintermeyer, L.; Myers, M.G. Measles in a Partially Immunized Community. Am. J. Public Health 1979, 69, 923–927. [Google Scholar] [CrossRef]
- Gans, H.A.; Yasukawa, L.L.; Sung, P.; Sullivan, B.; DeHovitz, R.; Audet, S.; Beeler, J.; Arvin, A.M. Measles Humoral and Cell-Mediated Immunity in Children Aged 5–10 Years after Primary Measles Immunization Administered at 6 or 9 Months of Age. J. Infect. Dis. 2013, 207, 574–582. [Google Scholar] [CrossRef]
- Li, M.; Wang, W.; Chen, J.; Zhan, Z.; Xu, M.; Liu, N.; Ren, L.; You, L.; Zheng, W.; Shi, H.; et al. Transplacental Transfer Efficiency of Maternal Antibodies against Influenza A(H1N1)Pdm09 Virus and Dynamics of Naturally Acquired Antibodies in Chinese Children: A Longitudinal, Paired Mother–Neonate Cohort Study. Lancet Microbe 2023, 4, e893–e902. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total (N = 408) | Born Before 2010 (N = 53) | Born After 2010 (N = 355) | p-Value | |
|---|---|---|---|---|
| Age at baseline, years | ||||
| Median (Interquartile range, IQR) | 0 (0−1.9) | 3.28 (3−3.9) | 0 (0−1.5) | 4.13 × 10−34 |
| Sex a | ||||
| Male | 199 (48.8) | 23 (43.4) | 176 (49.6) | 0.488 |
| Female | 209 (51.2) | 30 (56.6) | 179 (50.4) | |
| Socioeconomic status b | ||||
| Low | 64 (15.7) | 10 (18.9) | 54 (15.2) | 0.941 |
| Middle | 198 (48.5) | 28 (52.8) | 170 (47.9) | |
| High | 98 (24.0) | 15 (28.3) | 83 (23.4) | |
| Missing | 48 (11.8) | 0 (0) | 48 (13.5) | |
| Mode of delivery | ||||
| Vaginal delivery | 255 (62.5) | 35 (66.0) | 220 (62.0) | 0.676 |
| Caesarean section | 153 (37.5) | 18 (34.0) | 135 (38.0) | |
| Term of pregnancy | ||||
| Preterm birth | 17 (4.2) | 2 (3.8) | 15 (4.2) | 0.538 |
| Full-term birth | 379 (92.9) | 51 (96.2) | 328 (92.4) | |
| Post-term birth | 12 (2.9) | 0 (0.0) | 12 (3.4) | |
| Birth weight, grams | ||||
| Median (IQR) | 3300 (3000−3600) | 3250 (2925−3500) | 3300 (3000−3600) | 0.509 |
| Breastfeeding before 6 months | ||||
| No | 35 (8.6) | 6 (11.3) | 29 (8.2) | 0.432 |
| Yes | 373 (91.4) | 47 (88.7) | 326 (91.8) | |
| MCV doses received | ||||
| 2 doses | 403 (98.8) | 51 (96.2) | 352 (99.2) | 0.128 |
| 3 doses | 5 (1.2) | 2 (3.8) | 3 (0.8) | |
| Age at MCV1, months | ||||
| Median (IQR) | 8.2 (8.0−8.5) | 8.3 (8.1−8.5) | 8.2 (8.0−8.6) | 0.683 |
| Age at MCV2, months | ||||
| Median (IQR) | 18.2 (18.0−18.5) | 18.1 (18.0−18.5) | 18.2 (18.0−18.6) | 0.315 |
| Participants’ Characteristics | Mean | = 0.25) | = 0.50) | = 0.75) | |||||
|---|---|---|---|---|---|---|---|---|---|
| (95% CI) | p-Value | (95% CI) | p-Value | (95% CI) | p-Value | (95% CI) | p-Value | ||
| Intercept | 7.29 (7.05, 7.53) | 0.00 | 7.34 (7.03, 7.65) | 1.35 × 10−254 | 7.38 (7.07, 7.68) | 5.91 × 10−260 | 7.38 (7.08, 7.69) | 2.66 × 10−261 | |
| Time since MCV2, months b | |||||||||
| Median (IQR) | 31.2 (21.3, 47.7) | −0.01 (−0.02, −0.01) | 0.001 | −0.02 (−0.02, −0.01) | 4.20 × 10−9 | −0.01 (−0.02, −0.01) | 1.10 × 10−8 | −0.01 (−0.02, −0.01) | 4.48 × 10−10 |
| Sex a | |||||||||
| Male | 180 (47.7) | Reference | − | Reference | − | Reference | − | Reference | − |
| Female | 197 (52.3) | 0.24 (0.07, 0.41) | 0.006 | 0.12 (−0.07, 0.32) | 0.215 | 0.14 (−0.05, 0.34) | 0.146 | 0.16 (−0.03, 0.36) | 0.097 |
| Mode of delivery | |||||||||
| Vaginal delivery | 236 (62.6) | Reference | − | Reference | − | Reference | − | Reference | − |
| Caesarean section | 141 (37.4) | −0.06 (−0.23, 0.11) | 0.411 | −0.02 (−0.19, 0.16) | 0.864 | 0.00 (−0.18, 0.17) | 0.986 | −0.02 (−0.19, 0.15) | 0.828 |
| Birth cohort | |||||||||
| Before 2010 | 52 (13.8) | Reference | − | Reference | − | Reference | − | Reference | − |
| After 2010 | 325 (86.2) | −0.23 (−0.46, −0.01) | 0.046 | −0.29 (−0.55, −0.02) | 0.036 | −0.25 (−0.52, 0.01) | 0.061 | −0.23 (−0.50, 0.04) | 0.093 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, S.; Wang, Q.; Yang, J.; Liao, Q.; Zhang, J.; Zhou, X.; Zhou, J.; Zhao, Z.; Liang, Y.; Luo, J.; et al. Comparative Analysis of Long-Term Measles Immune Response After Natural Infection and Routine Vaccination in China. Vaccines 2025, 13, 555. https://doi.org/10.3390/vaccines13060555
Zhao S, Wang Q, Yang J, Liao Q, Zhang J, Zhou X, Zhou J, Zhao Z, Liang Y, Luo J, et al. Comparative Analysis of Long-Term Measles Immune Response After Natural Infection and Routine Vaccination in China. Vaccines. 2025; 13(6):555. https://doi.org/10.3390/vaccines13060555
Chicago/Turabian StyleZhao, Sihong, Qianli Wang, Juan Yang, Qiaohong Liao, Juanjuan Zhang, Xiaoyu Zhou, Jiaxin Zhou, Zeyao Zhao, Yuxia Liang, Junteng Luo, and et al. 2025. "Comparative Analysis of Long-Term Measles Immune Response After Natural Infection and Routine Vaccination in China" Vaccines 13, no. 6: 555. https://doi.org/10.3390/vaccines13060555
APA StyleZhao, S., Wang, Q., Yang, J., Liao, Q., Zhang, J., Zhou, X., Zhou, J., Zhao, Z., Liang, Y., Luo, J., Cai, J., Wu, Y., Wang, W., & Yu, H. (2025). Comparative Analysis of Long-Term Measles Immune Response After Natural Infection and Routine Vaccination in China. Vaccines, 13(6), 555. https://doi.org/10.3390/vaccines13060555