
Synthetic Peptides Elicit Humoral Response against Porcine Reproductive and Respiratory Syndrome Virus in Swine

 , , , and
, , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Peptides
2.2. Experimental Design, Animals, and Housing
2.3. Blood and Serum Sampling and Isolation of Peripheral Blood Mononuclear Cells
2.4. Quantification of Cytokine Concentrations in Serum and Antibody Detection
2.5. Immunoreactivity of Peptides against Serum Antibodies
2.6. Flow Cytometry
2.7. Statistical Analysis
3. Results
3.1. Body Weight Monitoring after Immunization
3.2. GP5-B3 Peptide Increased the Serum Concentration of Proinflammatory Cytokines
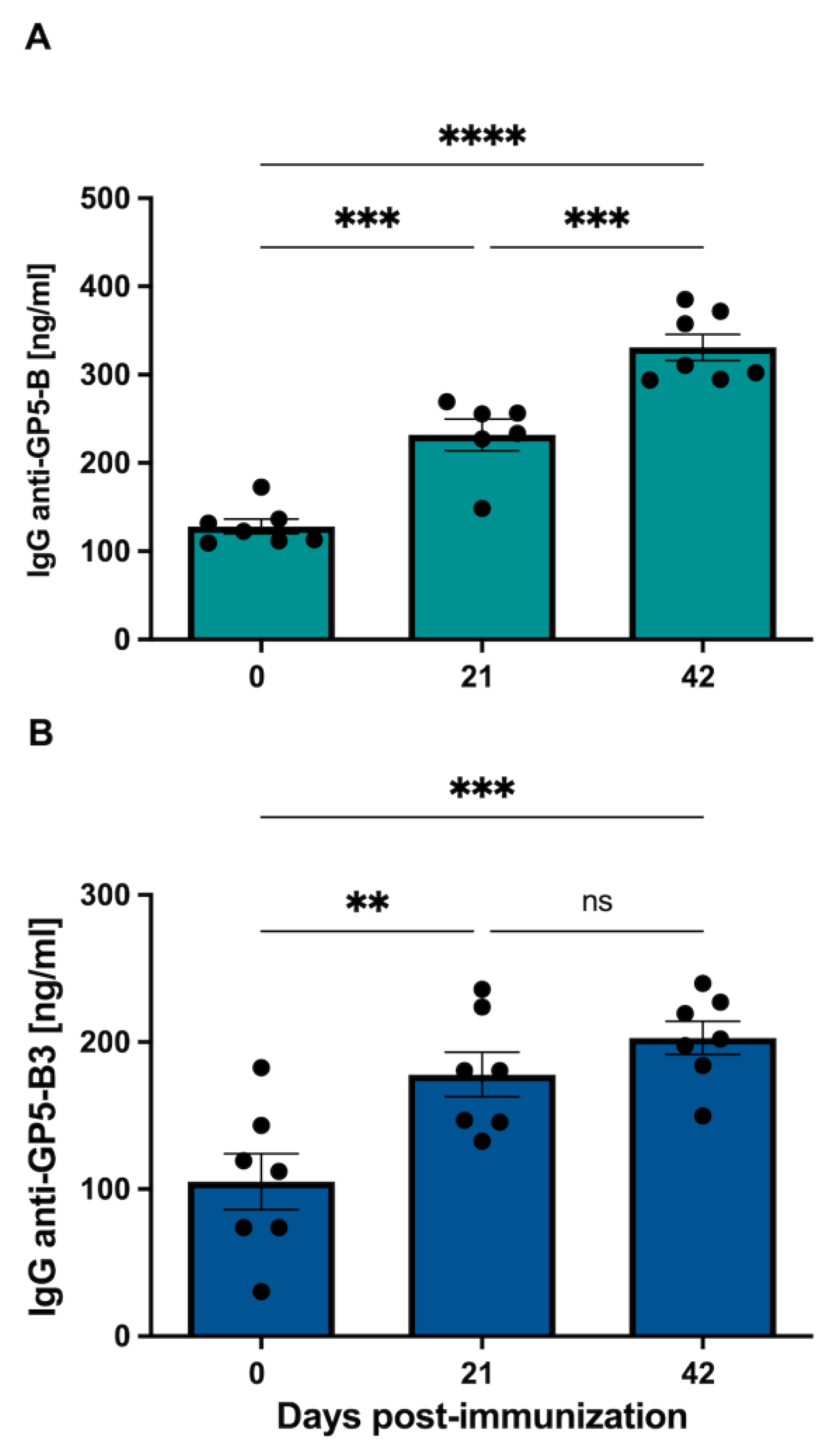
3.3. GP5-B and GP5-B3 Induce Specific IgG-Mediated Response upon Immunization
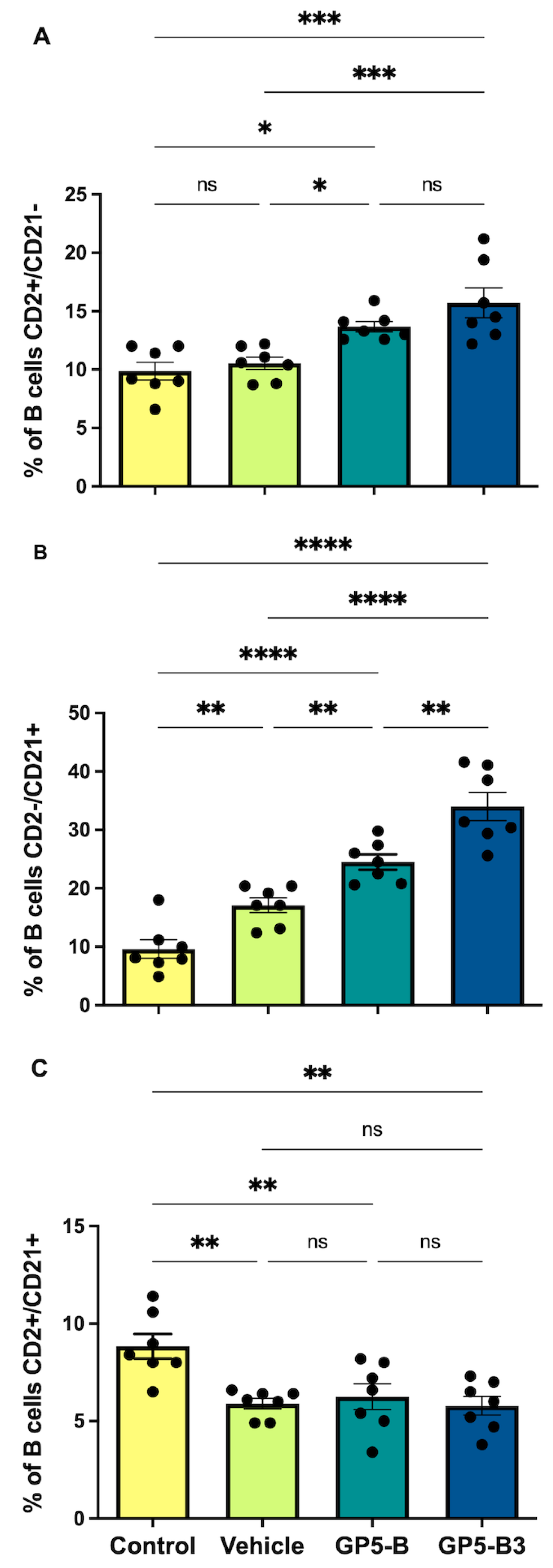
3.4. Peptides Prime B Cells and Induce the Generation of Antibody-Secreting Cells
3.5. GP5 Epitopes Elicit In Vitro Immunoreactivity in Sera from Naturally Infected and Vaccinated Pigs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nieuwenhuis, N.; Duinhof, T.F.; van Nes, A. Economic analysis of outbreaks of porcine reproductive and respiratory syndrome virus in nine sow herds. Vet. Rec. 2012, 170, 225. [Google Scholar] [CrossRef] [PubMed]
- Chase-Topping, M.; Xie, J.; Pooley, C.; Trus, I.; Bonckaert, C.; Rediger, K.; Bailey, R.I.; Brown, H.; Bitsouni, V.; Barrio, M.B.; et al. New insights about vaccine effectiveness: Impact of attenuated PRRS-strain vaccination on heterologous strain transmission. Vaccine 2020, 38, 3050–3061. [Google Scholar] [CrossRef] [PubMed]
- Ruedas-Torres, I.; Rodriguez-Gomez, I.M.; Sanchez-Carvajal, J.M.; Larenas-Munoz, F.; Pallares, F.J.; Carrasco, L.; Gomez-Laguna, J. The jigsaw of PRRSV virulence. Vet. Microbiol. 2021, 260, 109168. [Google Scholar] [CrossRef] [PubMed]
- Angulo, J.; Yang, M.; Rovira, A.; Davies, P.R.; Torremorell, M. Infection dynamics and incidence of wild-type porcine reproductive and respiratory syndrome virus in growing pig herds in the U.S. Midwest. Prev. Vet. Med. 2023, 217, 105976. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Qiao, S.; Zhang, G. Reappraising host cellular factors involved in attachment and entry to develop antiviral strategies against porcine reproductive and respiratory syndrome virus. Front. Microbiol. 2022, 13, 975610. [Google Scholar] [CrossRef] [PubMed]
- López-Heydeck, S.M.; Alonso-Morales, R.A.; Mendieta-Zerón, H.; Vázquez-Chagoyán, J.C. Síndrome reproductivo y respiratorio del cerdo (PRRS). Revisión. Rev. Mex. Cienc. Pecu. 2015, 6, 69–89. [Google Scholar]
- Pileri, E.; Mateu, E. Review on the transmission porcine reproductive and respiratory syndrome virus between pigs and farms and impact on vaccination. Vet. Res. 2016, 47, 108. [Google Scholar] [CrossRef]
- Lunney, J.K.; Fang, Y.; Ladinig, A.; Chen, N.; Li, Y.; Rowland, B.; Renukaradhya, G.J. Porcine Reproductive and Respiratory Syndrome Virus (PRRSV): Pathogenesis and Interaction with the Immune System. Annu. Rev. Anim. Biosci. 2016, 4, 129–154. [Google Scholar] [CrossRef]
- Sun, Q.; Xu, H.; An, T.; Cai, X.; Tian, Z.; Zhang, H. Recent Progress in Studies of Porcine Reproductive and Respiratory Syndrome Virus 1 in China. Viruses 2023, 15, 1528. [Google Scholar] [CrossRef]
- Mardassi, H.; Mounir, S.; Dea, S. Identification of major differences in the nucleocapsid protein genes of a Québec strain and European strains of porcine reproductive and respiratory syndrome virus. J. Gen. Virol. 1994, 75 Pt. 3, 681–685. [Google Scholar] [CrossRef]
- Duerlinger, S.; Knecht, C.; Sawyer, S.; Balka, G.; Zaruba, M.; Ruemenapf, T.; Kraft, C.; Rathkjen, P.H.; Ladinig, A. Efficacy of a Modified Live Porcine Reproductive and Respiratory Syndrome Virus 1 (PRRSV-1) Vaccine against Experimental Infection with PRRSV AUT15-33 in Weaned Piglets. Vaccines 2022, 10, 934. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.J.; Paul, P.S.; Halbur, P.G.; Lum, M.A. Characterization of a high-virulence US isolate of porcine reproductive and respiratory syndrome virus in a continuous cell line, ATCC CRL11171. J. Vet. Diagn. Investig. 1996, 8, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Snijder, E.J. The PRRSV replicase: Exploring the multifunctionality of an intriguing set of nonstructural proteins. Virus Res. 2010, 154, 61–76. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.H.; Nguyen, P.-L.; Ahn, J.-Y.; Yoon, H.; Min, J.; Lee, L.; Cho, S.-J.; Sekhon, S.S.; Kim, Y.-H. The global research trend of porcine reproductive and respiratory syndrome virus (PRRSV): A mini review. Toxicol. Environ. Health Sci. 2015, 7, 241–250. [Google Scholar] [CrossRef]
- Stoian, A.M.M.; Rowland, R.R.R. Challenges for Porcine Reproductive and Respiratory Syndrome (PRRS) Vaccine Design: Reviewing Virus Glycoprotein Interactions with CD163 and Targets of Virus Neutralization. Vet. Sci. 2019, 6, 9. [Google Scholar] [CrossRef] [PubMed]
- Montaner-Tarbes, S.; Del Portillo, H.A.; Montoya, M.; Fraile, L. Key Gaps in the Knowledge of the Porcine Respiratory Reproductive Syndrome Virus (PRRSV). Front. Vet. Sci. 2019, 6, 38. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Liu, X. Editorial: Porcine reproductive and respiratory syndrome virus—Animal virology, immunology, and pathogenesis. Front. Immunol. 2023, 14, 1194386. [Google Scholar] [CrossRef] [PubMed]
- Garcia Duran, M.; Costa, S.; Sarraseca, J.; de la Roja, N.; Garcia, J.; Garcia, I.; Rodriguez, M.J. Generation of porcine reproductive and respiratory syndrome (PRRS) virus-like-particles (VLPs) with different protein composition. J. Virol. Methods 2016, 236, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Wissink, E.H.; Kroese, M.V.; van Wijk, H.A.; Rijsewijk, F.A.; Meulenberg, J.J.; Rottier, P.J. Envelope protein requirements for the assembly of infectious virions of porcine reproductive and respiratory syndrome virus. J. Virol. 2005, 79, 12495–12506. [Google Scholar] [CrossRef]
- Renukaradhya, G.J.; Meng, X.J.; Calvert, J.G.; Roof, M.; Lager, K.M. Live porcine reproductive and respiratory syndrome virus vaccines: Current status and future direction. Vaccine 2015, 33, 4069–4080. [Google Scholar] [CrossRef]
- Nan, Y.; Wu, C.; Gu, G.; Sun, W.; Zhang, Y.J.; Zhou, E.M. Improved Vaccine against PRRSV: Current Progress and Future Perspective. Front. Microbiol. 2017, 8, 1635. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.M.; Ni, Y.Y.; Cao, D.; Tian, D.; Yugo, D.M.; Heffron, C.L.; Overend, C.; Subramaniam, S.; Rogers, A.J.; Catanzaro, N.; et al. Recombinant Porcine Reproductive and Respiratory Syndrome Virus Expressing Membrane-Bound Interleukin-15 as an Immunomodulatory Adjuvant Enhances NK and gammadelta T Cell Responses and Confers Heterologous Protection. J. Virol. 2018, 92, e00007-18. [Google Scholar] [CrossRef] [PubMed]
- Madapong, A.; Saeng-Chuto, K.; Boonsoongnern, A.; Tantituvanont, A.; Nilubol, D. Cell-mediated immune response and protective efficacy of porcine reproductive and respiratory syndrome virus modified-live vaccines against co-challenge with PRRSV-1 and PRRSV-2. Sci. Rep. 2020, 10, 1649. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; O’Connell, C.M.; Costa, A.; Pan, Y.; Smyth, J.A.; Verardi, P.H.; Burgess, D.J.; Van Kruiningen, H.J.; Garmendia, A.E. A PRRSV GP5-Mosaic vaccine: Protection of pigs from challenge and ex vivo detection of IFNγ responses against several genotype 2 strains. PLoS ONE 2019, 14, e0208801. [Google Scholar] [CrossRef] [PubMed]
- Kick, A.R.; Amaral, A.F.; Frias-De-Diego, A.; Cortes, L.M.; Fogle, J.E.; Crisci, E.; Almond, G.W.; Kaser, T. The Local and Systemic Humoral Immune Response Against Homologous and Heterologous Strains of the Type 2 Porcine Reproductive and Respiratory Syndrome Virus. Front. Immunol. 2021, 12, 637613. [Google Scholar] [CrossRef]
- Choi, K.; Park, C.; Jeong, J.; Kang, I.; Park, S.J.; Chae, C. Comparison of commercial type 1 and type 2 PRRSV vaccines against heterologous dual challenge. Vet. Rec. 2016, 178, 291. [Google Scholar] [CrossRef]
- Zhang, H.; Luo, Q.; He, Y.; Zheng, Y.; Sha, H.; Li, G.; Kong, W.; Liao, J.; Zhao, M. Research Progress on the Development of Porcine Reproductive and Respiratory Syndrome Vaccines. Vet. Sci. 2023, 10, 491. [Google Scholar] [CrossRef] [PubMed]
- Scortti, M.; Prieto, C.; Alvarez, E.; Simarro, I.; Castro, J.M. Failure of an inactivated vaccine against porcine reproductive and respiratory syndrome to protect gilts against a heterologous challenge with PRRSV. Vet. Rec. 2007, 161, 809–813. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kim, H.K.; Jung, J.H.; Choi, Y.J.; Kim, J.; Um, C.G.; Hyun, S.B.; Shin, S.; Lee, B.; Jang, G.; et al. The assessment of efficacy of porcine reproductive respiratory syndrome virus inactivated vaccine based on the viral quantity and inactivation methods. Virol. J. 2011, 8, 323. [Google Scholar] [CrossRef]
- Charerntantanakul, W. Porcine reproductive and respiratory syndrome virus vaccines: Immunogenicity, efficacy and safety aspects. World J. Virol. 2012, 1, 23–30. [Google Scholar] [CrossRef]
- Jeong, J.; Park, C.; Oh, T.; Park, K.H.; Yang, S.; Kang, I.; Park, S.J.; Chae, C. Cross-protection of a modified-live porcine reproductive and respiratory syndrome virus (PRRSV)-2 vaccine against a heterologous PRRSV-1 challenge in late-term pregnancy gilts. Vet. Microbiol. 2018, 223, 119–125. [Google Scholar] [CrossRef]
- Vreman, S.; Stockhofe-Zurwieden, N.; Popma-de Graaf, D.J.; Savelkoul, H.F.J.; Barnier-Quer, C.; Collin, N.; Collins, D.; McDaid, D.; Moore, A.C.; Rebel, J.M.J. Immune responses induced by inactivated porcine reproductive and respiratory syndrome virus (PRRSV) vaccine in neonatal pigs using different adjuvants. Vet. Immunol. Immunopathol. 2021, 232, 110170. [Google Scholar] [CrossRef]
- Du, T.; Nan, Y.; Xiao, S.; Zhao, Q.; Zhou, E.M. Antiviral Strategies against PRRSV Infection. Trends Microbiol. 2017, 25, 968–979. [Google Scholar] [CrossRef]
- Chen, C.; Li, J.; Bi, Y.; Yang, L.; Meng, S.; Zhou, Y.; Jia, X.; Meng, S.; Sun, L.; Liu, W. Synthetic B- and T-cell epitope peptides of porcine reproductive and respiratory syndrome virus with Gp96 as adjuvant induced humoral and cell-mediated immunity. Vaccine 2013, 31, 1838–1847. [Google Scholar] [CrossRef]
- Popescu, L.N.; Trible, B.R.; Chen, N.; Rowland, R.R.R. GP5 of porcine reproductive and respiratory syndrome virus (PRRSV) as a target for homologous and broadly neutralizing antibodies. Vet. Microbiol. 2017, 209, 90–96. [Google Scholar] [CrossRef]
- Murthy, A.M.V.; Ni, Y.; Meng, X.; Zhang, C. Production and Evaluation of Virus-Like Particles Displaying Immunogenic Epitopes of Porcine Reproductive and Respiratory Syndrome Virus (PRRSV). Int. J. Mol. Sci. 2015, 16, 8382–8396. [Google Scholar] [CrossRef]
- Ostrowski, M.; Galeota, J.A.; Jar, A.M.; Platt, K.B.; Osorio, F.A.; Lopez, O.J. Identification of neutralizing and nonneutralizing epitopes in the porcine reproductive and respiratory syndrome virus GP5 ectodomain. J. Virol. 2002, 76, 4241–4250. [Google Scholar] [CrossRef]
- Zhao, G.; Zhang, J.; Sun, W.; Xie, C.; Zhang, H.; Gao, Y.; Wen, S.; Ha, Z.; Nan, F.; Zhu, X.; et al. Immunological evaluation of recombination PRRSV GP3 and GP5 DNA vaccines in vivo. Front. Cell Infect. Microbiol. 2022, 12, 1016897. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Zheng, Y.; Zhang, H.; Yang, Z.; Sha, H.; Kong, W.; Zhao, M.; Wang, N. Research Progress on Glycoprotein 5 of Porcine Reproductive and Respiratory Syndrome Virus. Animals 2023, 13, 813. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Chen, Y. Epitope screening and vaccine molecule design of PRRSV GP3 and GP5 protein based on immunoinformatics. J. Cell Mol. Med. 2024, 28, e18103. [Google Scholar] [CrossRef] [PubMed]
- Purcell, A.W.; McCluskey, J.; Rossjohn, J. More than one reason to rethink the use of peptides in vaccine design. Nat. Rev. Drug Discov. 2007, 6, 404–414. [Google Scholar] [CrossRef]
- Charerntantanakul, W.; Platt, R.; Johnson, W.; Roof, M.; Vaughn, E.; Roth, J.A. Immune responses and protection by vaccine and various vaccine adjuvant candidates to virulent porcine reproductive and respiratory syndrome virus. Vet. Immunol. Immunopathol. 2006, 109, 99–115. [Google Scholar] [CrossRef]
- Charerntantanakul, W. Adjuvants for swine vaccines: Mechanisms of actions and adjuvant effects. Vaccine 2020, 38, 6659–6681. [Google Scholar] [CrossRef]
- Vashisht, K.; Goldberg, T.L.; Husmann, R.J.; Schnitzlein, W.; Zuckermann, F.A. Identification of immunodominant T-cell epitopes present in glycoprotein 5 of the North American genotype of porcine reproductive and respiratory syndrome virus. Vaccine 2008, 26, 4747–4753. [Google Scholar] [CrossRef]
- Calderon-Rico, F.; Bravo-Patiño, A.; Mendieta, I.; Perez-Duran, F.; Zamora-Aviles, A.G.; Franco-Correa, L.E.; Ortega-Flores, R.; Hernandez-Morales, I.; Nuñez-Anita, R.E. Glycoprotein 5-Derived Peptides Induce a Protective T-Cell Response in Swine against the Porcine Reproductive and Respiratory Syndrome Virus. Viruses 2023, 16, 14. [Google Scholar] [CrossRef]
- Lopez, O.J.; Osorio, F.A. Role of neutralizing antibodies in PRRSV protective immunity. Vet. Immunol. Immunopathol. 2004, 102, 155–163. [Google Scholar] [CrossRef]
- Pettini, E.; Medaglini, D.; Ciabattini, A. Profiling the B cell immune response elicited by vaccination against the respiratory virus SARS-CoV-2. Front. Immunol. 2022, 13, 1058748. [Google Scholar] [CrossRef]
- Opriessnig, T.; Mattei, A.A.; Karuppannan, A.K.; Halbur, P.G. Future perspectives on swine viral vaccines: Where are we headed? Porc. Health Manag. 2021, 7, 1. [Google Scholar] [CrossRef] [PubMed]
- Rahe, M.C.; Dvorak, C.M.T.; Patterson, A.; Roof, M.; Murtaugh, M.P. The PRRSV-Specific Memory B Cell Response Is Long-Lived in Blood and Is Boosted During Live Virus Re-exposure. Front. Immunol. 2020, 11, 247. [Google Scholar] [CrossRef] [PubMed]
- Braun, R.O.; Python, S.; Summerfield, A. Porcine B Cell Subset Responses to Toll-like Receptor Ligands. Front. Immunol. 2017, 8, 1044. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Wu, M.; Albo, J.; Rao, Q. Non-Specific Binding and Cross-Reaction of ELISA: A Case Study of Porcine Hemoglobin Detection. Foods 2021, 10, 1708. [Google Scholar] [CrossRef]
- Engvall, E. The ELISA, enzyme-linked immunosorbent assay. Clin. Chem. 2010, 56, 319–320. [Google Scholar] [CrossRef]
- Guzman-Bautista, E.R.; Garcia-Ruiz, C.E.; Gama-Espinosa, A.; Ramirez-Estudillo, C.; Rojas-Gomez, O.I.; Vega-Lopez, M.A. Effect of age and maternal antibodies on the systemic and mucosal immune response after neonatal immunization in a porcine model. Immunology 2014, 141, 609–616. [Google Scholar] [CrossRef]
- Paudyal, B.; Mwangi, W.; Rijal, P.; Schwartz, J.C.; Noble, A.; Shaw, A.; Sealy, J.E.; Bonnet-Di Placido, M.; Graham, S.P.; Townsend, A.; et al. Fc-Mediated Functions of Porcine IgG Subclasses. Front. Immunol. 2022, 13, 903755. [Google Scholar] [CrossRef]
- Buermann, A.; Borns, K.; Römermann, D.; Plege-Fleck, A.; Schwinzer, R. B cell activation and induced antibody responses to porcine antigen can be diminished by PD-L1-mediated triggering of PD-1. Xenotransplantation 2014, 21, 192. [Google Scholar] [CrossRef]
- Sinkora, M.; Butler, J.E.; Lager, K.M.; Potockova, H.; Sinkorova, J. The comparative profile of lymphoid cells and the T and B cell spectratype of germ-free piglets infected with viruses SIV, PRRSV or PCV2. Vet. Res. 2014, 45, 91. [Google Scholar] [CrossRef]
- Sinkora, M.; Sinkorova, J. B cell lymphogenesis in swine is located in the bone marrow. J. Immunol. 2014, 193, 5023–5032. [Google Scholar] [CrossRef]
- Turner, J.S.; Marthi, M.; Benet, Z.L.; Grigorova, I. Transiently antigen-primed B cells return to naive-like state in absence of T-cell help. Nat. Commun. 2017, 8, 15072. [Google Scholar] [CrossRef]
- Kienzler, A.-K.; Eibel, H. Human B Cell Development and Tolerance. In Encyclopedia of Immunobiology; Ratcliffe, M.J.H., Ed.; Academic Press: Oxford, UK, 2016; pp. 105–121. [Google Scholar]
- Mulupuri, P.; Zimmerman, J.J.; Hermann, J.; Johnson, C.R.; Cano, J.P.; Yu, W.; Dee, S.A.; Murtaugh, M.P. Antigen-specific B-cell responses to porcine reproductive and respiratory syndrome virus infection. J. Virol. 2008, 82, 358–370. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Lu, H.; Wen, S.; Sun, W.; Yan, F.; Chen, X.; Jing, J.; Liu, H.; Liu, C.; Xue, F.; et al. Enhanced immune responses in pigs by DNA vaccine coexpressing GP3 and GP5 of European type porcine reproductive and respiratory syndrome virus. J. Virol. Methods 2014, 206, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Thorarinsdottir, K.; Camponeschi, A.; Cavallini, N.; Grimsholm, O.; Jacobsson, L.; Gjertsson, I.; Mårtensson, I.L. CD21(-/low) B cells in human blood are memory cells. Clin. Exp. Immunol. 2016, 185, 252–262. [Google Scholar] [CrossRef]
- Vanhee, M.; Van Breedam, W.; Costers, S.; Geldhof, M.; Noppe, Y.; Nauwynck, H. Characterization of antigenic regions in the porcine reproductive and respiratory syndrome virus by the use of peptide-specific serum antibodies. Vaccine 2011, 29, 4794–4804. [Google Scholar] [CrossRef]
- Su, C.M.; Rowland, R.R.R.; Yoo, D. Recent Advances in PRRS Virus Receptors and the Targeting of Receptor-Ligand for Control. Vaccines 2021, 9, 354. [Google Scholar] [CrossRef]
- An, T.Q.; Li, J.N.; Su, C.M.; Yoo, D. Molecular and Cellular Mechanisms for PRRSV Pathogenesis and Host Response to Infection. Virus Res. 2020, 286, 197980. [Google Scholar] [CrossRef]
- Young, J.E.; Dvorak, C.M.T.; Graham, S.P.; Murtaugh, M.P. Isolation of Porcine Reproductive and Respiratory Syndrome Virus GP5-Specific, Neutralizing Monoclonal Antibodies From Hyperimmune Sows. Front. Immunol. 2021, 12, 638493. [Google Scholar] [CrossRef]
- Thaa, B.; Sinhadri, B.C.; Tielesch, C.; Krause, E.; Veit, M. Signal peptide cleavage from GP5 of PRRSV: A minor fraction of molecules retains the decoy epitope, a presumed molecular cause for viral persistence. PLoS ONE 2013, 8, e65548. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.H.; Chen, J.; Tong, G.Z.; Tian, Z.J.; Zhou, Y.J.; Li, G.X.; Li, X.; Peng, J.M.; An, T.Q.; Yang, H.C. A recombinant plasmid co-expressing swine ubiquitin and the GP5 encoding-gene of porcine reproductive and respiratory syndrome virus induces protective immunity in piglets. Vaccine 2008, 26, 1438–1449. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Origin | Farm | Group (N = 7) | Formulation | Commercial Vaccine 104.9DICC50 | ||||
|---|---|---|---|---|---|---|---|---|
| PBS (µL) | Carrier Maleimide BSA (µg/µL) | Adjuvant Aluminum Hydroxide (µL) | GP5-B Peptide (µg/µL) | GP5-B3 Peptide (µg/µL) | ||||
| El Dapo | La Carreta | Control | 800 | |||||
| Vehicle | 200 | 200 | 400 | |||||
| GP5-B | 200 | 400 | 200 | |||||
| GP5-B3 | 200 | 400 | 200 | |||||
| El Dapo | Vaccinated | 2000 | ||||||
| El Limon | El Limon | Natural infection | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perez-Duran, F.; Calderon-Rico, F.; Franco-Correa, L.E.; Zamora-Aviles, A.G.; Ortega-Flores, R.; Durand-Herrera, D.; Bravo-Patiño, A.; Cortes-Vieyra, R.; Hernandez-Morales, I.; Nuñez-Anita, R.E. Synthetic Peptides Elicit Humoral Response against Porcine Reproductive and Respiratory Syndrome Virus in Swine. Vaccines 2024, 12, 652. https://doi.org/10.3390/vaccines12060652
Perez-Duran F, Calderon-Rico F, Franco-Correa LE, Zamora-Aviles AG, Ortega-Flores R, Durand-Herrera D, Bravo-Patiño A, Cortes-Vieyra R, Hernandez-Morales I, Nuñez-Anita RE. Synthetic Peptides Elicit Humoral Response against Porcine Reproductive and Respiratory Syndrome Virus in Swine. Vaccines. 2024; 12(6):652. https://doi.org/10.3390/vaccines12060652
Chicago/Turabian StylePerez-Duran, Francisco, Fernando Calderon-Rico, Luis Enrique Franco-Correa, Alicia Gabriela Zamora-Aviles, Roberto Ortega-Flores, Daniel Durand-Herrera, Alejandro Bravo-Patiño, Ricarda Cortes-Vieyra, Ilane Hernandez-Morales, and Rosa Elvira Nuñez-Anita. 2024. "Synthetic Peptides Elicit Humoral Response against Porcine Reproductive and Respiratory Syndrome Virus in Swine" Vaccines 12, no. 6: 652. https://doi.org/10.3390/vaccines12060652
APA StylePerez-Duran, F., Calderon-Rico, F., Franco-Correa, L. E., Zamora-Aviles, A. G., Ortega-Flores, R., Durand-Herrera, D., Bravo-Patiño, A., Cortes-Vieyra, R., Hernandez-Morales, I., & Nuñez-Anita, R. E. (2024). Synthetic Peptides Elicit Humoral Response against Porcine Reproductive and Respiratory Syndrome Virus in Swine. Vaccines, 12(6), 652. https://doi.org/10.3390/vaccines12060652