
Combined Live Oral Priming and Intramuscular Boosting Regimen with Rotarix® and a Nanoparticle-Based Trivalent Rotavirus Vaccine Evaluated in Gnotobiotic Pig Models of G4P[6] and G1P[8] Human Rotavirus Infection

 , , , and
, , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Rotaviruses for Challenge and Immunoassays
2.2. Vaccine
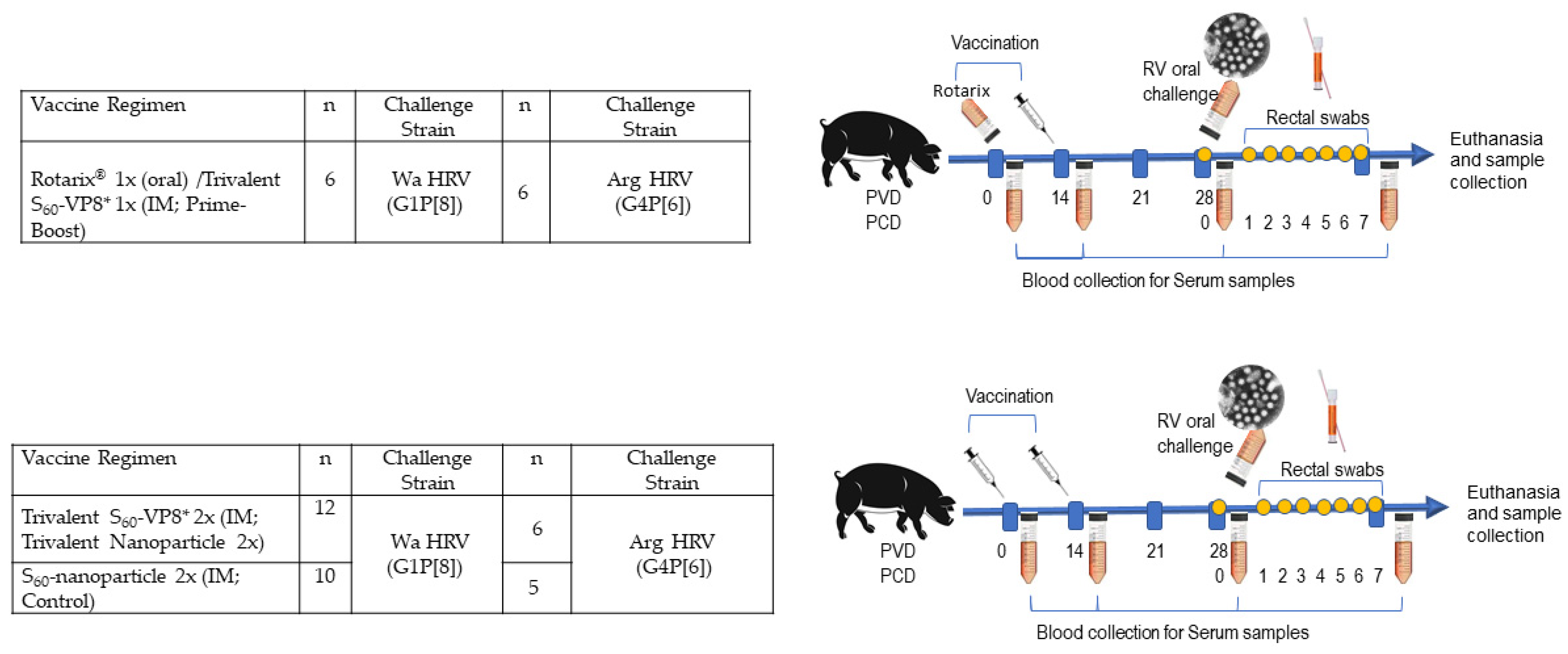
2.3. Vaccine Inoculation, Virus Challenge, and Sample Collection of Gn Pigs
2.4. Assessment of Diarrhea and Detection of Fecal Virus Shedding by Antigen ELISA and Virus CCIF
2.5. Detection of HRV VP8*-Specific Serum and Intestinal IgA and IgG Antibody by ELISA
2.6. Flow Cytometry for Detection of IFN-γ-Producing CD3+CD4+ and CD3+CD8+ T Cells
2.7. Detection of VP8*-Specific Antibody-Secreting Cells by ELISpot Assay
2.8. Virus Neutralization Assay
2.9. Statistical Analysis
3. Results
3.1. Prime–Boost Regimen Significantly Reduced Virus Shedding in Both Challenge Groups
3.2. Both Vaccine Regimens Were Highly Immunogenic and Induced Strong Serum IgG and IgA Responses in Gn Pigs before and after Challenge with Wa or Arg HRV
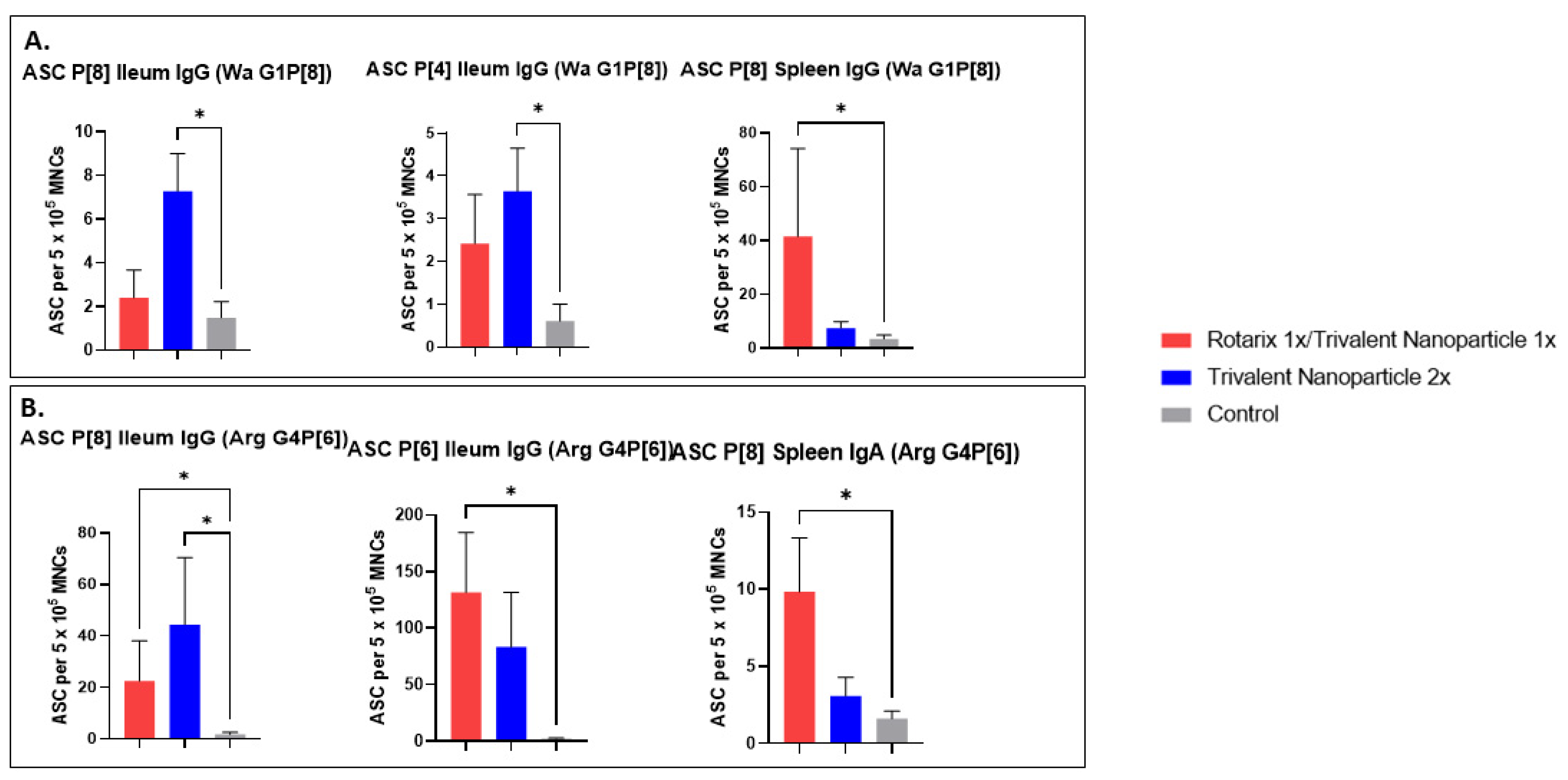
3.3. Prime–Boost Vaccinated Gn Pigs Challenged with Arg HRV Had Significantly Higher P[8]-Specific IgA in the Small Intestine at Euthanasia
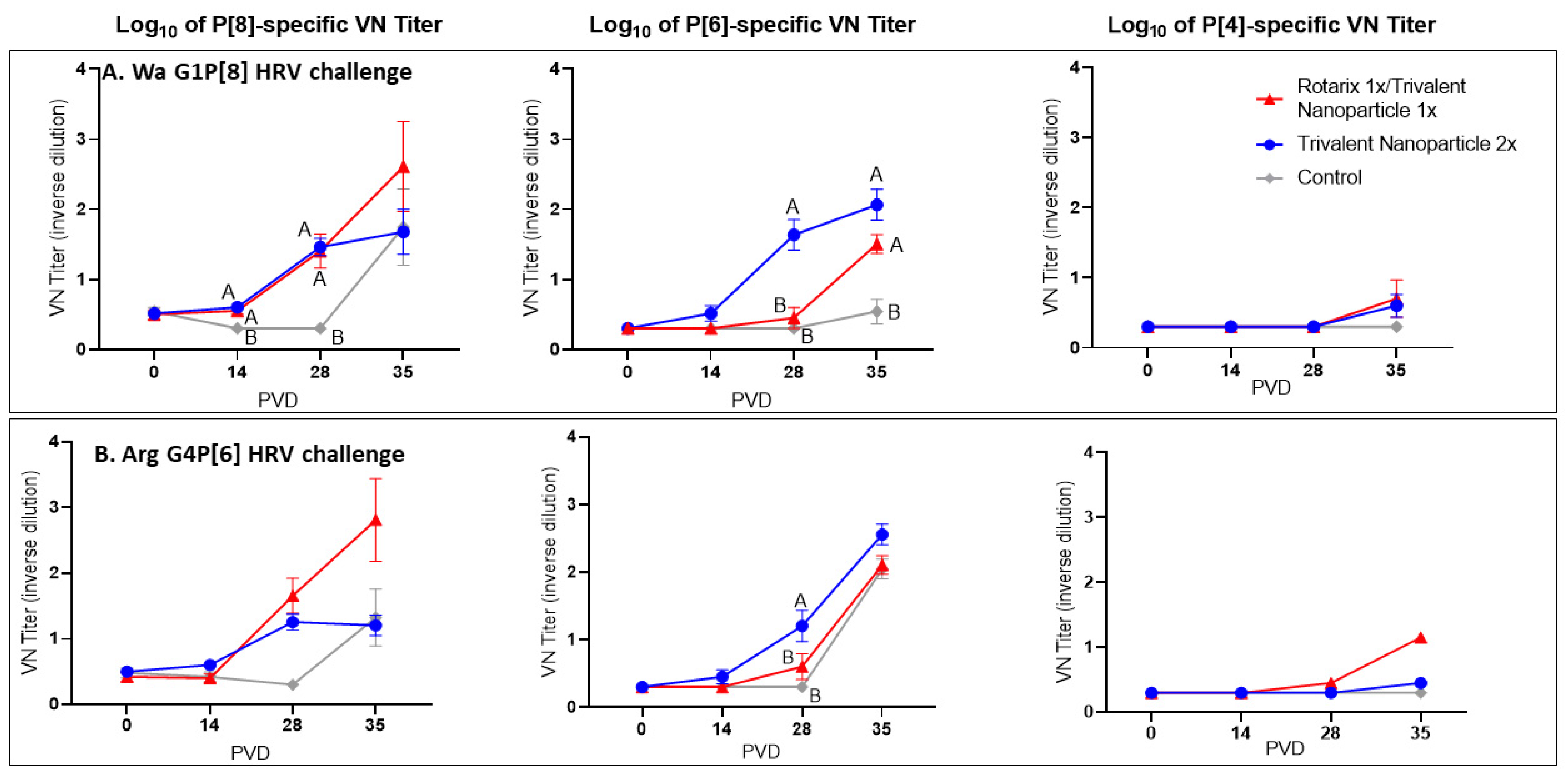
3.4. Both Vaccine Regimens Induced Strong P[8]- and P[6]-Specific Virus Neutralizing Antibody Responses in the Serum of Wa and Arg HRV-Challenged Gn Pigs
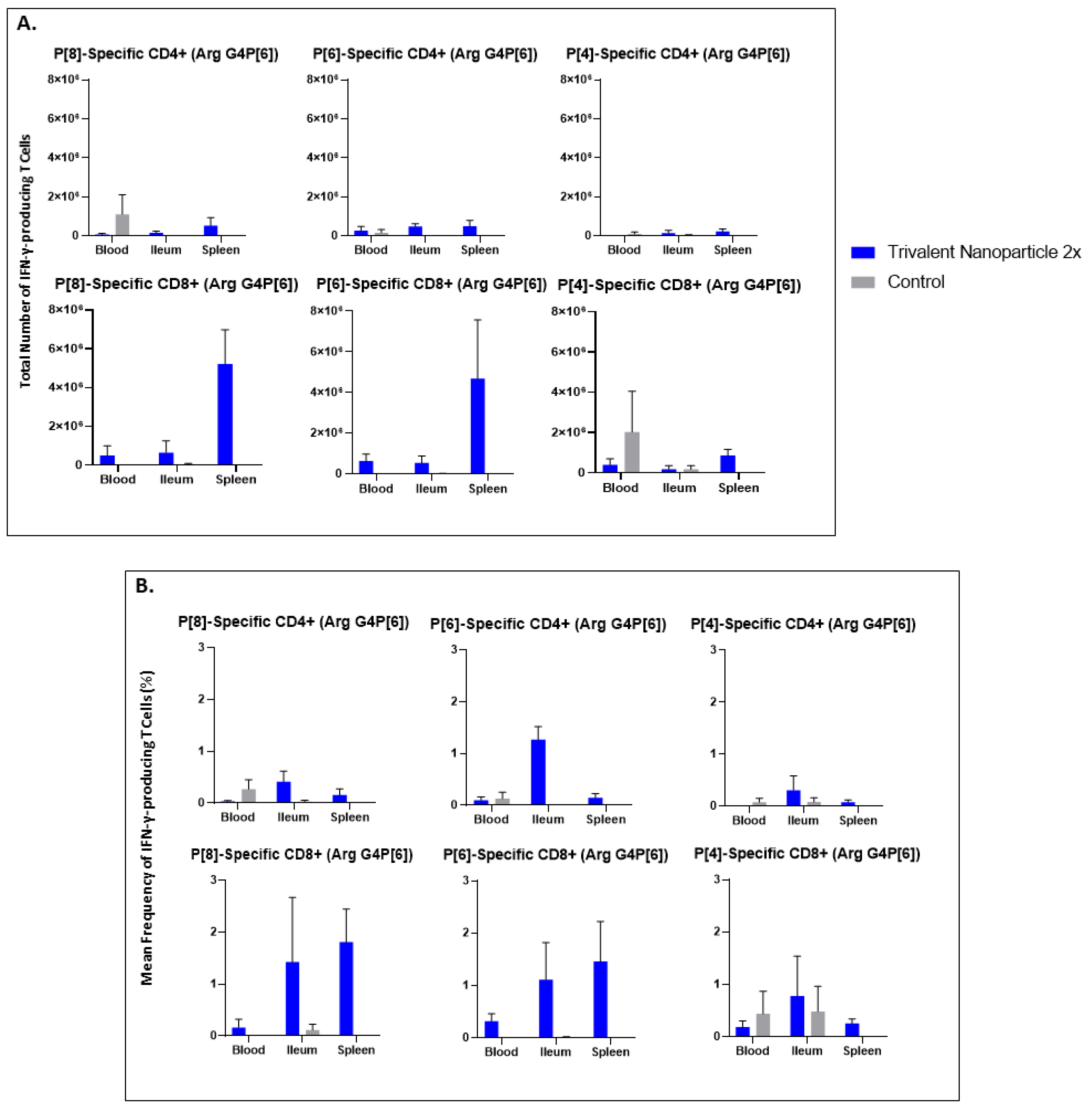
3.5. Trivalent Nanoparticle 2×-Vaccinated, Wa HRV-Challenged Gn Pigs Had Significantly Higher Total Numbers and Frequencies of P[8]-Specific CD3+CD4+IFN-γ+ and CD3+CD8+IFN-γ+ T Cells in the Ileum Post-Challenge
3.6. Both Vaccine Regimens in Both Challenge Groups Induced Significant P-Type-Specific ASC Responses in the Ileum and Spleen
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cárcamo-Calvo, R.; Muñoz, C.; Buesa, J.; Rodríguez-Díaz, J.; Gozalbo-Rovira, R. The Rotavirus Vaccine Landscape, an Update. Pathogens 2021, 10, 520. [Google Scholar] [CrossRef]
- Donato, C.M.; Bines, J.E. Rotaviruses and Rotavirus Vaccines. Pathogens 2021, 10, 959. [Google Scholar] [CrossRef] [PubMed]
- Parashar, U.D.; Hummelman, E.G.; Bresee, J.S.; Miller, M.A.; Glass, R.I. Global Illness and Deaths Caused by Rotavirus Disease in Children. Emerg. Infect. Dis. 2003, 9, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, G.; Ma, J.Z.; Haque, R.; Petri, W.A. Rotavirus vaccine protection in low-income and middle-income countries. Lancet Infect. Dis. 2019, 19, 673–674. [Google Scholar] [CrossRef]
- Burnett, E.; Parashar, U.D.; Tate, J.E. Real-world effectiveness of rotavirus vaccines, 2006–2019: A literature review and meta-analysis. Lancet Glob. Health 2020, 8, e1195–e1202. [Google Scholar] [CrossRef]
- Glass, R.I.; Bresee, J.; Jiang, B.; Parashar, U.; Yee, E.; Gentsch, J. Rotavirus and rotavirus vaccines. Adv. Exp. Med. Biol. 2006, 582, 45–54. [Google Scholar]
- Lopman, B.A.; Pitzer, V.E.; Sarkar, R.; Gladstone, B.; Patel, M.; Glasser, J.; Gambhir, M.; Atchison, C.; Grenfell, B.T.; Edmunds, W.J.; et al. Understanding Reduced Rotavirus Vaccine Efficacy in Low Socio-Economic Settings. PLoS ONE 2012, 7, e41720. [Google Scholar] [CrossRef] [PubMed]
- Parker, E.P.; Ramani, S.; Lopman, B.A.; Church, J.A.; Iturriza, M.; Prendergast, A.J.; Grassly, N. Causes of impaired oral vaccine efficacy in developing countries. Future Microbiol. 2018, 13, 97–118. [Google Scholar] [CrossRef]
- Chen, M.-Y.; Kirkwood, C.D.; Bines, J.; Cowley, D.; Pavlic, D.; Lee, K.J.; Orsini, F.; Watts, E.; Barnes, G.; Danchin, M. Rotavirus specific maternal antibodies and immune response to RV3-BB neonatal rotavirus vaccine in New Zealand. Hum. Vaccines Immunother. 2017, 13, 1126–1135. [Google Scholar] [CrossRef] [PubMed]
- Rennels, M.B.; Ward, R.L.; Mack, M.E.; Zito, E.T.; US Rotavirus Vaccine Efficacy Group. Concurrent Oral Poliovirus and Rhesus-Human Reassortant Rotavirus Vaccination: Effects on Immune Responses to Both Vaccines and on Efficacy of Rotavirus Vaccines. J. Infect. Dis. 1996, 173, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Harris, V.; Ali, S.A.; Fuentes, S.; Korpela, K.; Kazi, M.; Tate, J.; Parashar, U.; Wiersinga, W.J.; Giaquinto, C.; De Weerth, C.; et al. Rotavirus vaccine response correlates with the infant gut microbiota composition in Pakistan. Gut Microbes 2017, 9, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Ruck, C.E.; Odumade, O.A.; Smolen, K.K. Vaccine Interactions With the Infant Microbiome: Do They Define Health and Disease? Front. Pediatr. 2020, 8, 565368. [Google Scholar] [CrossRef] [PubMed]
- Michael, H.; Langel, S.; Miyazaki, A.; Paim, F.C.; Chepngeno, J.; Alhamo, M.A.; Fischer, D.D.; Srivastava, V.; Kathayat, D.; Deblais, L.; et al. Malnutrition Decreases Antibody Secreting Cell Numbers Induced by an Oral Attenuated Human Rotavirus Vaccine in a Human Infant Fecal Microbiota Transplanted Gnotobiotic Pig Model. Front. Immunol. 2020, 11, 196. [Google Scholar] [CrossRef] [PubMed]
- Qadri, F.; Bhuiyan, T.R.; Sack, D.A.; Svennerholm, A.-M. Immune responses and protection in children in developing countries induced by oral vaccines. Vaccine 2012, 31, 452–460. [Google Scholar] [CrossRef] [PubMed]
- Ramani, S.; Mamani, N.; Villena, R.; Bandyopadhyay, A.S.; Gast, C.; Sato, A.; Laucirica, D.; Clemens, R.; Estes, M.K.; O’ryan, M.L. Rotavirus Serum IgA Immune Response in Children Receiving Rotarix Coadministered With bOPV or IPV. Pediatr. Infect. Dis. J. 2016, 35, 1137–1139. [Google Scholar] [CrossRef]
- Nyblade, C.; Hensley, C.; Parreño, V.; Zhou, P.; Frazier, M.; Frazier, A.; Ramesh, A.; Lei, S.; Degiuseppe, J.I.; Tan, M.; et al. A New Gnotobiotic Pig Model of P[6] Human Rotavirus Infection and Disease for Preclinical Evaluation of Rotavirus Vaccines. Viruses 2022, 14, 2803. [Google Scholar] [CrossRef]
- Steele, A.D.; Neuzil, K.M.; Cunliffe, N.A.; Madhi, S.A.; Bos, P.; Ngwira, B.; Witte, D.; Todd, S.; Louw, C.; Kirsten, M.; et al. Human rotavirus vaccine Rotarix™ provides protection against diverse circulating rotavirus strains in African infants: A randomized controlled trial. BMC Infect. Dis. 2012, 12, 213. [Google Scholar] [CrossRef]
- Vesikari, T.; Matson, D.O.; Dennehy, P.; Van Damme, P.; Santosham, M.; Rodriguez, Z.; Dallas, M.J.; Heyse, J.F.; Goveia, M.G.; Black, S.B.; et al. Safety and Efficacy of a Pentavalent Human–Bovine (WC3) Reassortant Rotavirus Vaccine. N. Engl. J. Med. 2006, 354, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Koch, J.; Harder, T.; von Kries, R.; Wichmann, O. Risk of Intussusception After Rotavirus Vaccination: A Systematic Literature Review and Meta-Analysis. Dtsch. Ärzteblatt Int. 2017, 114, 255–262. [Google Scholar] [CrossRef]
- Leino, T.; Ollgren, J.; Strömberg, N.; Elonsalo, U. Evaluation of the Intussusception Risk after Pentavalent Rotavirus Vaccination in Finnish Infants. PLoS ONE 2016, 11, e0144812. [Google Scholar] [CrossRef] [PubMed]
- Pitzer, V.E.; Bennett, A.; Bar-Zeev, N.; Jere, K.C.; Lopman, B.A.; Lewnard, J.A.; Parashar, U.D.; Cunliffe, N.A. Evaluating strategies to improve rotavirus vaccine impact during the second year of life in Malawi. Sci. Transl. Med. 2019, 11, eaav6419. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L. Vaccine Efficacy Evaluation: The Gnotobiotic Pig Model, 1st ed.; CRC Press: Boca Raton, FL, USA; Taylor and Francis Group: Oxford, UK, 2022; pp. 1–205. [Google Scholar]
- Yuan, L.; Saif, L.J. Induction of mucosal immune responses and protection against enteric viruses: Rotavirus infection of gnotobiotic pigs as a model. Vet. Immunol. Immunopathol. 2002, 87, 147–160. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Grow, S.; Iturriza-Gómara, M.; Hausdorff, W.P.; Fix, A.; Kirkwood, C.D. The Challenges and Opportunities of Next-Generation Rotavirus Vaccines: Summary of an Expert Meeting with Vaccine Developers. Viruses 2022, 14, 2565. [Google Scholar] [CrossRef] [PubMed]
- Knipe, D.M.; Howley, P. Fields Virology, 6th ed.; Wolters Kluwer/Lippincott Williams & Wilkins Health: Philadelphia, PA, USA, 2013; Volume 2. [Google Scholar]
- Liu, Y.; Huang, P.; Tan, M.; Liu, Y.; Biesiada, J.; Meller, J.; Castello, A.A.; Jiang, B.; Jiang, X. Rotavirus VP8*: Phylogeny, host range, and interaction with histo-blood group antigens. J. Virol. 2012, 86, 9899–9910. [Google Scholar] [CrossRef] [PubMed]
- Feng, N.; Hu, L.; Ding, S.; Sanyal, M.; Zhao, B.; Sankaran, B.; Ramani, S.; McNeal, M.; Yasukawa, L.L.; Song, Y.; et al. Human VP8* mAbs neutralize rotavirus selectively in human intestinal epithelial cells. J. Clin. Investig. 2019, 130, 3839–3851. [Google Scholar] [CrossRef]
- Ruggeri, F.M.; Greenberg, H.B. Antibodies to the Trypsin Cleavage Peptide Vp8-Star Neutralize Rotavirus by Inhibiting Binding of Virions to Target-Cells in Culture. J. Virol. 1991, 65, 2211–2219. [Google Scholar] [CrossRef]
- Kovacs-Nolan, J.; Mine, Y. Tandem copies of a human rotavirus VP8 epitope can induce specific neutralizing antibodies in BALB/c mice. Biochim. et Biophys. Acta (BBA)—Gen. Subj. 2006, 1760, 1884–1893. [Google Scholar] [CrossRef]
- Lee, J.; Babiuk, L.A.; Harland, R.; Gibbons, E.; Elazhary, Y.; Yoo, D. Immunological response to recombinant VP8* subunit protein of bovine roravirus in pregnant cattle. J. Gen. Virol. 1995, 76 Pt 10, 2477–2483. [Google Scholar] [CrossRef]
- Xia, M.; Huang, P.; Sun, C.; Han, L.; Vago, F.S.; Li, K.; Zhong, W.; Jiang, W.; Klassen, J.S.; Jiang, X.; et al. Bioengineered Norovirus S60 Nanoparticles as a Multifunctional Vaccine Platform. ACS Nano 2018, 12, 10665–10682. [Google Scholar] [CrossRef]
- Xia, M.; Hoq, R.; Huang, P.; Jiang, W.; Jiang, X.; Tan, M. Bioengineered pseudovirus nanoparticles displaying the HA1 antigens of influenza viruses for enhanced immunogenicity. Nano Res. 2022, 15, 4181–4190. [Google Scholar] [CrossRef]
- Xia, M.; Huang, P.; Jiang, X.; Tan, M. Immune response and protective efficacy of the S particle presented rotavirus VP8* vaccine in mice. Vaccine 2019, 37, 4103–4110. [Google Scholar] [CrossRef]
- Xia, M.; Huang, P.; Tan, M. A Pseudovirus Nanoparticle-Based Trivalent Rotavirus Vaccine Candidate Elicits High and Cross P Type Immune Response. Pharmaceutics 2022, 14, 1597. [Google Scholar] [CrossRef]
- Ward, L.A.; Rosen, B.I.; Yuan, L.; Saif, L.J. Pathogenesis of an attenuated and a virulent strain of group A human rotavirus in neonatal gnotobiotic pigs. J. Gen. Virol. 1996, 77 Pt 7, 1431–1441. [Google Scholar] [CrossRef]
- Yuan, L.; Jobst, P.M.; Weiss, M. Gnotobiotic Pigs: From Establishing Facility to Modeling Human Infectious Diseases. In Gnotobiotics; Trenton, K.A.E., Schoeb, R., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 349–364. [Google Scholar]
- Yuan, L.; Kang, S.Y.; Ward, L.A.; To, T.L.; Saif, L.J. Antibody-secreting cell responses and protective immunity assessed in gnotobiotic pigs inoculated orally or intramuscularly with inactivated human rotavirus. J. Virol. 1998, 72, 330–338. [Google Scholar] [CrossRef]
- Yuan, L.; Wen, K.; Azevedo, M.S.; Gonzalez, A.M.; Zhang, W.; Saif, L.J. Virus-specific intestinal IFN-gamma producing T cell responses induced by human rotavirus infection and vaccines are correlated with protection against rotavirus diarrhea in gnotobiotic pigs. Vaccine 2008, 26, 3322–3331. [Google Scholar] [CrossRef]
- Vega, C.; Garaicoechea, L.; Degiuseppe, J.; Bok, M.; Rivolta, A.; Piantanida, A.; Asenzo, G.; Guerrero, M.A.; Wigdorovitz, A.; Stupka, J.; et al. ROTADIAL: The first nanobody-based immunoassay to detect Group A Rotavirus. J. Virol. Methods 2021, 298, 114279. [Google Scholar] [CrossRef]
- Garaicoechea, L.; Olichon, A.; Marcoppido, G.; Wigdorovitz, A.; Mozgovoj, M.; Saif, L.; Surrey, T.; Parreño, V. Llama-Derived Single-Chain Antibody Fragments Directed to Rotavirus VP6 Protein Possess Broad Neutralizing Activity In Vitro and Confer Protection against Diarrhea in Mice. J. Virol. 2008, 82, 9753–9764. [Google Scholar] [CrossRef]
- Hemming, M.; Huhti, L.; Räsänen, S.; Salminen, M.; Vesikari, T. Rotavirus Antigenemia in Children is Associated With More Severe Clinical Manifestations of Acute Gastroenteritis. Pediatr. Infect. Dis. J. 2014, 33, 366–371. [Google Scholar] [CrossRef]
- Clements, J.D.; Freytag, L.C. Parenteral Vaccination Can Be an Effective Means of Inducing Protective Mucosal Responses. Clin. Vaccine Immunol. 2016, 23, 438–441. [Google Scholar] [CrossRef]
- De Calisto, J.; Villablanca, E.J.; Wang, S.; Bono, M.R.; Rosemblatt, M.; Mora, J.R. T-Cell Homing to the Gut Mucosa: General Concepts and Methodological Considerations. Methods Mol. Biol. 2011, 757, 411–434. [Google Scholar] [CrossRef]
- Dudda, J.C.; Simon, J.C.; Martin, S. Dendritic Cell Immunization Route Determines CD8+ T Cell Trafficking to Inflamed Skin: Role for Tissue Microenvironment and Dendritic Cells in Establishment of T Cell-Homing Subsets. J. Immunol. 2004, 172, 857–863. [Google Scholar] [CrossRef]
- Blutt, S.E.; Matson, D.O.; Crawford, S.E.; Staat, M.A.; Azimi, P.; Bennett, B.L.; Piedra, P.A.; Conner, M.E. Rotavirus Antigenemia in Children Is Associated with Viremia. PLoS Med. 2007, 4, e121. [Google Scholar] [CrossRef]
- Justino, M.C.A.; Campos, E.A.; Mascarenhas, J.D.P.; Soares, L.S.; Guerra, S.D.F.S.; Furlaneto, I.P.; Pavao, M.J.C., Jr.; Maciel, T.S.; Farias, F.P.; Bezerra, O.M.; et al. Rotavirus antigenemia as a common event among children hospitalised for severe, acute gastroenteritis in Belém, northern Brazil. BMC Pediatr. 2019, 19, 193. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Signs of Diarrhea b | Virus Shedding (CCIF) b | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatments (Vaccine Dose µg) a | n | Percentage with Diarrhea | Mean Days to Onset | Mean Duration Days | Mean Cumulative Fecal Score | AUC of Diarrhea | Percentage of Shedding Virus | Mean Days to Onset | Mean Duration Days | Mean Peak Titer (FFU/g of Feces) e | AUC of Virus Shedding e |
| Rotarix 1x/Trivalent Nanoparticle 1x | 6 | 100% | 2 | 3.70 | 10.3 | 8.70 | 100% | 2.33 (0.21) A,c,d | 3.00 (0.4) B | 18,594 | 51,800 |
| Trivalent Nanoparticle 2x | 12 | 100% | 2 | 4.58 | 12.4 | 11.1 | 100% | 1.33 (0.14) B | 5.67 (0.38) A | 24,771 | 80,583 |
| Control | 10 | 100% | 2 | 4.86 | 13.0 | 11.4 | 100% | 1.70 (0.33) AB | 5.00 (0.37) A | 24,552 | 57,440 |
| Clinical Signs of Diarrhea b | Virus Shedding (CCIF) b | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatments (Vaccine Dose µg) a | n | Percentage with Diarrhea | Mean Days to Onset | Mean Duration Days | Mean Cumulative Fecal Score | AUC of Diarrhea | Percentage of Shedding Virus | Mean Days to Onset | Mean Duration Days | Mean Peak Titer (FFU/g of Feces) e | AUC of Virus Shedding e |
| Rotarix 1x/Trivalent Nanoparticle 1x | 6 | 100% | 2 | 4.70 | 13.0 | 10.8 | 100% | 1.50 | 3.33 (0.67) B,c,d | 15,554 (26,842) C | 78,200 (41,088) B |
| Trivalent Nanoparticle 2x | 6 | 100% | 2 | 5.00 | 14.2 | 12.3 | 100% | 1.17 | 6.00 (0.37) A | 40,778 (15,756) B | 138,767 (47,474) B |
| Control | 5 | 100% | 1 | 5.20 | 14.6 | 12.8 | 100% | 1.00 | 6.80 (0.20) A | 107,187 (36,620) A | 368,600 (93,322) A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hensley, C.; Nyblade, C.; Zhou, P.; Parreño, V.; Ramesh, A.; Frazier, A.; Frazier, M.; Garrison, S.; Fantasia-Davis, A.; Cai, R.; et al. Combined Live Oral Priming and Intramuscular Boosting Regimen with Rotarix® and a Nanoparticle-Based Trivalent Rotavirus Vaccine Evaluated in Gnotobiotic Pig Models of G4P[6] and G1P[8] Human Rotavirus Infection. Vaccines 2023, 11, 927. https://doi.org/10.3390/vaccines11050927
Hensley C, Nyblade C, Zhou P, Parreño V, Ramesh A, Frazier A, Frazier M, Garrison S, Fantasia-Davis A, Cai R, et al. Combined Live Oral Priming and Intramuscular Boosting Regimen with Rotarix® and a Nanoparticle-Based Trivalent Rotavirus Vaccine Evaluated in Gnotobiotic Pig Models of G4P[6] and G1P[8] Human Rotavirus Infection. Vaccines. 2023; 11(5):927. https://doi.org/10.3390/vaccines11050927
Chicago/Turabian StyleHensley, Casey, Charlotte Nyblade, Peng Zhou, Viviana Parreño, Ashwin Ramesh, Annie Frazier, Maggie Frazier, Sarah Garrison, Ariana Fantasia-Davis, Ruiqing Cai, and et al. 2023. "Combined Live Oral Priming and Intramuscular Boosting Regimen with Rotarix® and a Nanoparticle-Based Trivalent Rotavirus Vaccine Evaluated in Gnotobiotic Pig Models of G4P[6] and G1P[8] Human Rotavirus Infection" Vaccines 11, no. 5: 927. https://doi.org/10.3390/vaccines11050927
APA StyleHensley, C., Nyblade, C., Zhou, P., Parreño, V., Ramesh, A., Frazier, A., Frazier, M., Garrison, S., Fantasia-Davis, A., Cai, R., Huang, P.-W., Xia, M., Tan, M., & Yuan, L. (2023). Combined Live Oral Priming and Intramuscular Boosting Regimen with Rotarix® and a Nanoparticle-Based Trivalent Rotavirus Vaccine Evaluated in Gnotobiotic Pig Models of G4P[6] and G1P[8] Human Rotavirus Infection. Vaccines, 11(5), 927. https://doi.org/10.3390/vaccines11050927