
Vaccines Alone Cannot Slow the Evolution of SARS-CoV-2

 , , , and
, , , and 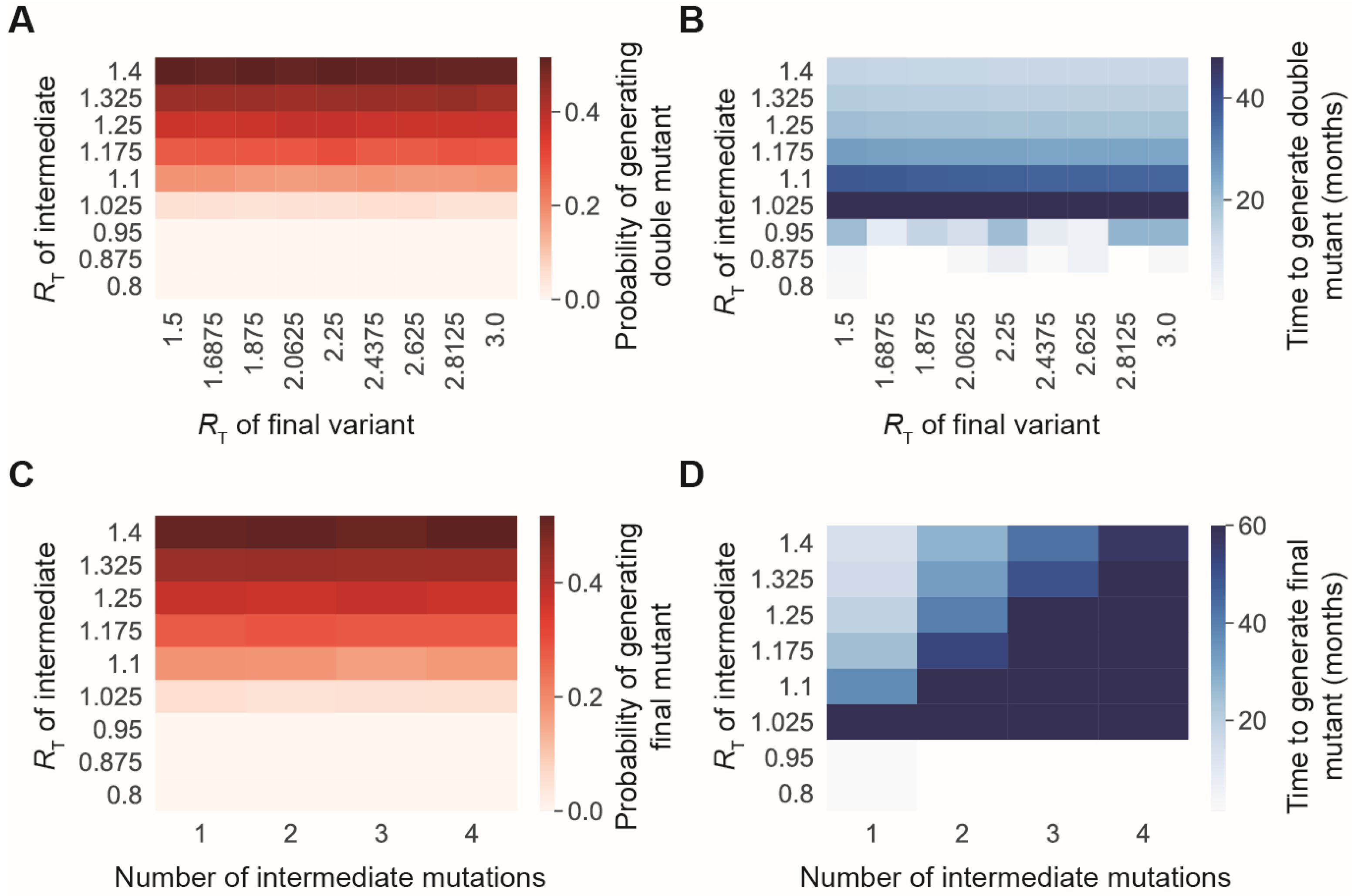{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Branching Process Model of Stochastic SARS-CoV-2 Transmission Dynamics
2.2. Extended SIR Model of Viral Transmission, Mutation, and Vaccination
2.3. Simulation of Intrahost Mutation Dynamics
3. Results
3.1. The Risk of Generating New SARS-CoV-2 Variants Strongly Depends on the Transmission Rate of Existing Variants
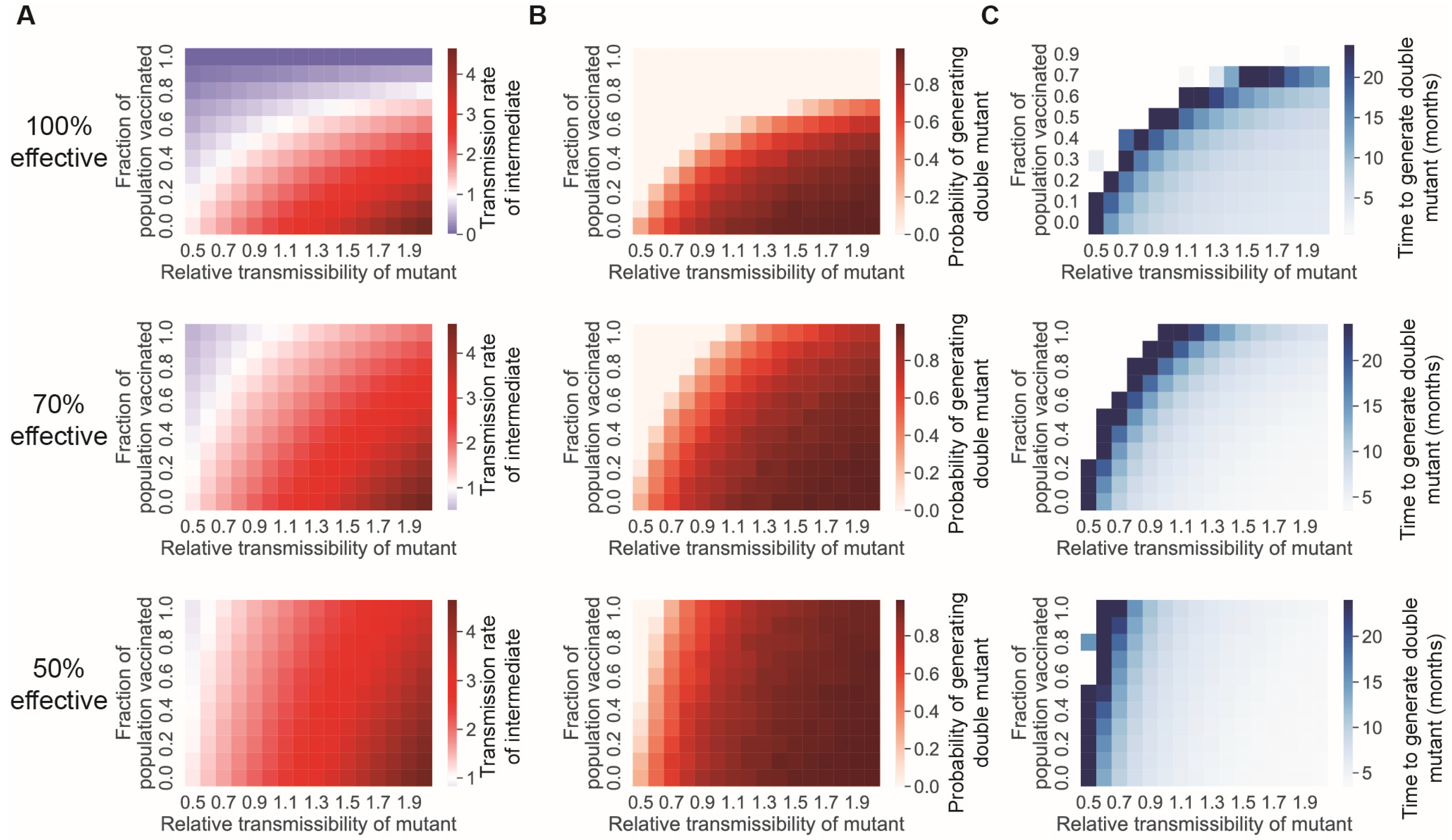
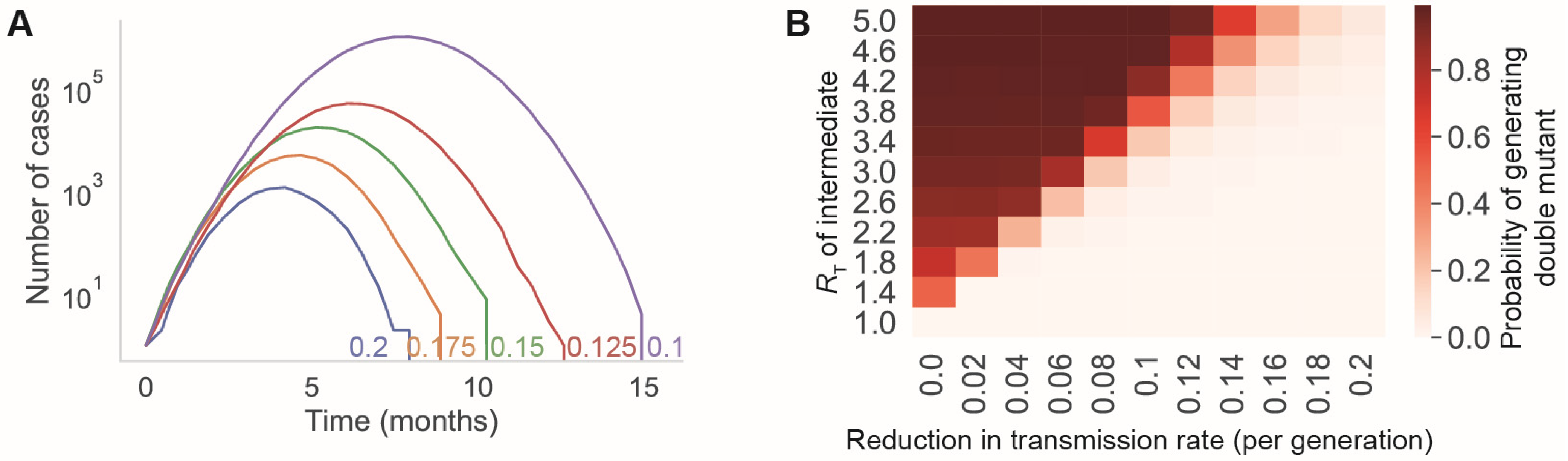
3.2. Reducing the Transmission Rate through Vaccination and/or Nonpharmaceutical Interventions Reduces the Rate at Which New Mutations Appear
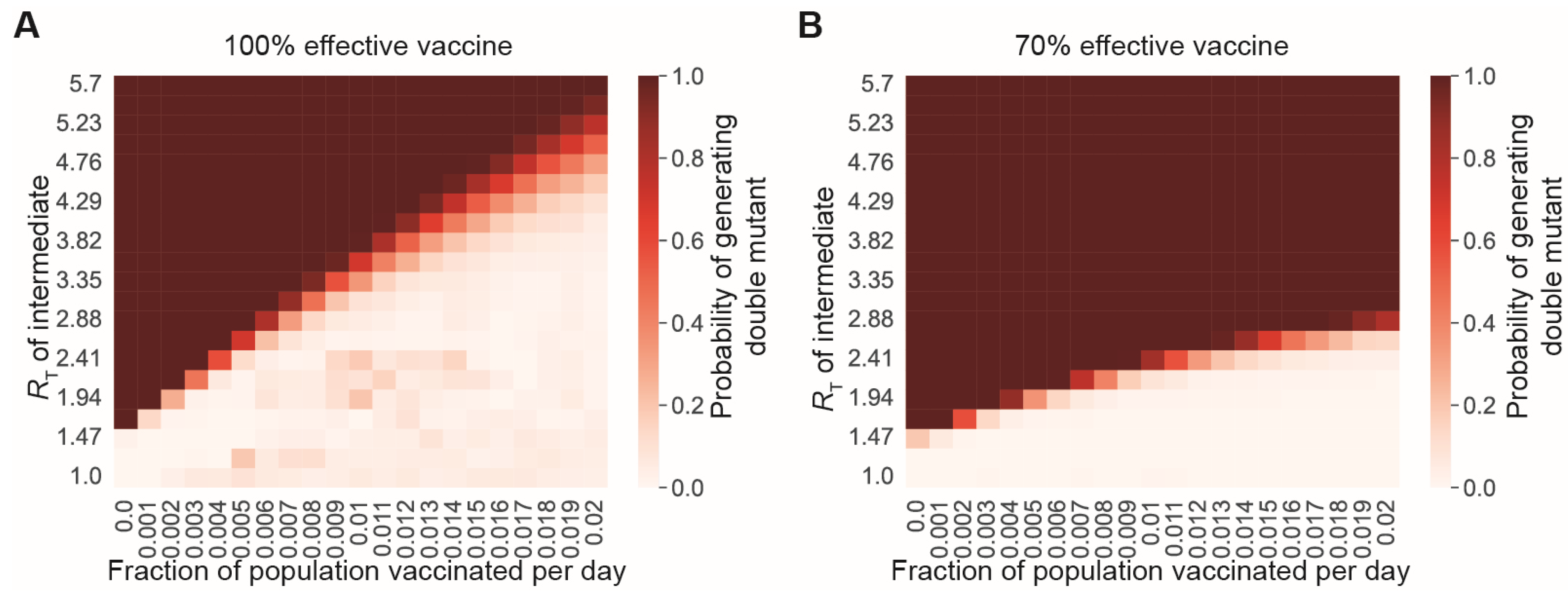
3.3. A Vaccine-Only Strategy for Controlling SARS-CoV-2 Spread Likely Cannot Prevent the Generation of New Immune-Evading Variants
3.4. Vaccines Do Not Effectively Prevent the Generation and Selection of New Mutants within Individual Hosts
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lavine, J.S.; Bjornstad, O.N.; Antia, R. Immunological Characteristics Govern the Transition of COVID-19 to Endemicity. Science 2021, 371, 741–745. [Google Scholar] [CrossRef] [PubMed]
- Lavine, J.S.; Bjornstad, O.N.; Coombs, D.; Antia, R. Severity of SARS-CoV-2 Reinfections in Second Wave Determines Likelihood of Mild Endemicity. MedRxiv 2021. [Google Scholar]
- Phillips, S. The New Normal for COVID Calls for a New Narrative. Available online: https://www.scientificamerican.com/article/the-new-normal-for-covid-calls-for-a-new-narrative/ (accessed on 4 October 2022).
- Ghebreyesus, T.A. WHO Director-General’s Opening Remarks at the COVID-19 Media Briefing. Available online: https://www.who.int/director-general/speeches/detail/who-director-general-s-opening-remarks-at-the-covid-19-media-briefing--17-august-2022 (accessed on 4 October 2022).
- Trottier, H.; Moore, D. Managing Population Health Risks as We Learn to Live with COVID-19. Can. J. Public Health 2022, 113, 649–652. [Google Scholar] [CrossRef] [PubMed]
- The White House Press Briefing by White House COVID-19 Response Team and Public Health Officials. Available online: https://www.whitehouse.gov/briefing-room/press-briefings/2021/12/17/press-briefing-by-white-house-covid-19-response-team-and-public-health-officials-74/ (accessed on 4 October 2022).
- Cao, Y.; Yisimayi, A.; Jian, F.; Song, W.; Xiao, T.; Wang, L.; Du, S.; Wang, J.; Li, Q.; Chen, X.; et al. BA.2.12.1, BA.4 and BA.5 Escape Antibodies Elicited by Omicron Infection. Nature 2022, 608, 593–602. [Google Scholar] [CrossRef]
- Katzourakis, A. COVID-19: Endemic Doesn’t Mean Harmless. Nature 2022, 601, 485. [Google Scholar] [CrossRef]
- Prater, E. You’re Probably Going to Get COVID at Least Once a Year. Available online: https://fortune.com/2022/05/25/how-often-can-you-get-covid-yearly-modeling-shows/ (accessed on 4 October 2022).
- Constantino, A.K. Dr. Fauci: BA.5-Specific Booster Shots Are the Country’s “Best Guess” for Dealing with COVID This Fall. Available online: https://www.cnbc.com/2022/07/27/fauci-ba5-boosters-are-best-guess-for-dealing-with-covid-this-fall.html (accessed on 4 October 2022).
- WHO Interim Statement on the Use of Additional Booster Doses of Emergency Use Listed MRNA Vaccines against COVID-19. Available online: https://www.who.int/news/item/17-05-2022-interim-statement-on-the-use-of-additional-booster-doses-of-emergency-use-listed-mrna-vaccines-against-covid-19 (accessed on 4 October 2022).
- Adams, K.; Rhoads, J.P.; Surie, D.; Gaglani, M.; Ginde, A.A.; McNeal, T.; Ghamande, S.; Huynh, D.; Talbot, H.K.; Casey, J.D.; et al. Vaccine Effectiveness of Primary Series and Booster Doses against Omicron Variant COVID-19-Associated Hospitalization in the United States. medRxiv 2022. [Google Scholar]
- Feikin, D.R.; Abu-Raddad, L.J.; Andrews, N.; Davies, M.-A.; Higdon, M.M.; Orenstein, W.A.; Patel, M.K. Assessing Vaccine Effectiveness against Severe COVID-19 Disease Caused by Omicron Variant. Report from a Meeting of the World Health Organization. Vaccine 2022, 40, 3516–3527. [Google Scholar] [CrossRef] [PubMed]
- Lauring, A.S.; Tenforde, M.W.; Chappell, J.D.; Gaglani, M.; Ginde, A.A.; McNeal, T.; Ghamande, S.; Douin, D.J.; Talbot, H.K.; Casey, J.D.; et al. Clinical Severity of, and Effectiveness of MRNA Vaccines against, Covid-19 from Omicron, Delta, and Alpha SARS-CoV-2 Variants in the United States: Prospective Observational Study. BMJ 2022, 376, e069761. [Google Scholar] [CrossRef]
- Eyre, D.W.; Taylor, D.; Purver, M.; Chapman, D.; Fowler, T.; Pouwels, K.B.; Walker, A.S.; Peto, T.E.A. Effect of Covid-19 Vaccination on Transmission of Alpha and Delta Variants. New Engl. J. Med. 2022, 386, 744–756. [Google Scholar] [CrossRef]
- Kislaya, I.; Rodrigues, E.F.; Borges, V.; Gomes, J.P.; Sousa, C.; Almeida, J.P.; Peralta-Santos, A.; Nunes, B. Comparative Effectiveness of Coronavirus Vaccine in Preventing Breakthrough Infections among Vaccinated Persons Infected with Delta and Alpha Variants. Emerg. Infect Dis. 2022, 28, 331–337. [Google Scholar] [CrossRef]
- Gram, M.A.; Emborg, H.-D.; Schelde, A.B.; Friis, N.U.; Nielsen, K.F.; Moustsen-Helms, I.R.; Legarth, R.; Lam, J.U.H.; Chaine, M.; Malik, A.Z.; et al. Vaccine Effectiveness against SARS-CoV-2 Infection and COVID-19-Related Hospitalization with the Alpha, Delta and Omicron SARS-CoV-2 Variants: A Nationwide Danish Cohort Study. PLoS Med. 2022, 19, e1003992. [Google Scholar] [CrossRef] [PubMed]
- Puhach, O.; Adea, K.; Hulo, N.; Sattonnet, P.; Genecand, C.; Iten, A.; Jacquérioz, F.; Kaiser, L.; Vetter, P.; Eckerle, I.; et al. Infectious Viral Load in Unvaccinated and Vaccinated Individuals Infected with Ancestral, Delta or Omicron SARS-CoV-2. Nat. Med. 2022, 28, 1491–1500. [Google Scholar] [CrossRef]
- Kissler, S.M.; Fauver, J.R.; Mack, C.; Tai, C.G.; Breban, M.I.; Watkins, A.E.; Samant, R.M.; Anderson, D.J.; Metti, J.; Khullar, G.; et al. Viral Dynamics of SARS-CoV-2 Variants in Vaccinated and Unvaccinated Persons. New Engl. J. Med. 2021, 385, 2489–2491. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.; Kim, J.Y.; Park, H.; Park, S.; Lim, J.S.; Lim, S.Y.; Bae, S.; Lim, Y.-J.; Kim, E.O.; Kim, J.; et al. Transmission and Infectious SARS-CoV-2 Shedding Kinetics in Vaccinated and Unvaccinated Individuals. JAMA Netw. Open 2022, 5, e2213606. [Google Scholar] [CrossRef] [PubMed]
- Singanayagam, A.; Hakki, S.; Dunning, J.; Madon, K.J.; Crone, M.A.; Koycheva, A.; Derqui-Fernandez, N.; Barnett, J.L.; Whitfield, M.G.; Varro, R.; et al. Community Transmission and Viral Load Kinetics of the SARS-CoV-2 Delta (B.1.617.2) Variant in Vaccinated and Unvaccinated Individuals in the UK: A Prospective, Longitudinal, Cohort Study. Lancet Infect. Dis. 2022, 22, 183–195. [Google Scholar] [CrossRef]
- Madewell, Z.J.; Yang, Y.; Longini, I.M.; Halloran, M.E.; Dean, N.E. Household Secondary Attack Rates of SARS-CoV-2 by Variant and Vaccination Status. JAMA Netw. Open 2022, 5, e229317. [Google Scholar] [CrossRef]
- Hagan, L.M. Outbreak of SARS-CoV-2 B.1.617.2 (Delta) Variant Infections Among Incarcerated Persons in a Federal Prison—Texas, July–August 2021. MMWR Morb. Mortal Wkly Rep. 2021, 70, 1349–1354. [Google Scholar] [CrossRef]
- Stoddard, M.; Novokhodko, A.; Sarkar, S.; Egeren, D.V.; White, L.F.; Hochberg, N.S.; Rogers, M.; Zetter, B.; Joseph-McCarthy, D.; Chakravarty, A. Endemicity Is Not a Victory: The Unmitigated Downside Risks of Widespread SARS-CoV-2 Transmission. COVID 2022, 2, 1689–1709. [Google Scholar] [CrossRef]
- Sauer, M.A.; Truelove, S.; Gerste, A.K.; Limaye, R.J. A Failure to Communicate? How Public Messaging Has Strained the COVID-19 Response in the United States. Health Secur. 2021, 19, 65–74. [Google Scholar] [CrossRef]
- LaFraniere, S.; Weiland, N. Walensky, Citing Botched Pandemic Response, Calls for C.D.C. Reorganization; The New York Times: New York, NY, USA, 2022. [Google Scholar]
- Kupferschmidt, K. Viral Evolution May Herald New Pandemic Phase. Science 2021, 371, 108–109. [Google Scholar] [CrossRef] [PubMed]
- Van Egeren, D.; Novokhodko, A.; Stoddard, M.; Tran, U.; Zetter, B.; Rogers, M.; Pentelute, B.L.; Carlson, J.M.; Hixon, M.; Joseph-McCarthy, D.; et al. Risk of Rapid Evolutionary Escape from Biomedical Interventions Targeting SARS-CoV-2 Spike Protein. PLoS ONE 2021, 16, e0250780. [Google Scholar] [CrossRef]
- Saad-Roy, C.M.; Morris, S.E.; Metcalf, C.J.E.; Mina, M.J.; Baker, R.E.; Farrar, J.; Holmes, E.C.; Pybus, O.G.; Graham, A.L.; Levin, S.A.; et al. Epidemiological and Evolutionary Considerations of SARS-CoV-2 Vaccine Dosing Regimes. Science 2021, 372, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, D.; Toy, S.; Douglas, J. The Covid Race to Watch: Vaccines vs. Variants. Available online: https://www.wsj.com/articles/covid-race-vaccine-vs-variants-11641915035 (accessed on 4 October 2022).
- Sah, P.; Vilches, T.N.; Moghadas, S.M.; Fitzpatrick, M.C.; Singer, B.H.; Hotez, P.J.; Galvani, A.P. Accelerated Vaccine Rollout Is Imperative to Mitigate Highly Transmissible COVID-19 Variants. eClinicalMedicine 2021, 35, 100865. [Google Scholar] [CrossRef] [PubMed]
- van Dorp, L.; Acman, M.; Richard, D.; Shaw, L.P.; Ford, C.E.; Ormond, L.; Owen, C.J.; Pang, J.; Tan, C.C.S.; Boshier, F.A.T.; et al. Emergence of Genomic Diversity and Recurrent Mutations in SARS-CoV-2. Infect. Genet. Evol. 2020, 83, 104351. [Google Scholar] [CrossRef] [PubMed]
- Karlin, S. A First Course in Stochastic Processes; Academic Press: Cambridge, MA, USA, 1966; pp. 291–295. [Google Scholar]
- Van Egeren, D.; Stoddard, M.; Novokhodko, A.; Rogers, M.S.; Joseph-McCarthy, D.; Zetter, B.; Chakravarty, A. Rapid Relaxation of Pandemic Restrictions after Vaccine Rollout Favors Growth of SARS-CoV-2 Variants: A Model-Based Analysis. PLoS ONE 2021, 16, e0258997. [Google Scholar] [CrossRef] [PubMed]
- Van Egeren, D.; Novokhodko, A.; Stoddard, M.; Tran, U.; Zetter, B.; Rogers, M.S.; Joseph-McCarthy, D.; Chakravarty, A. Controlling Long-Term SARS-CoV-2 Infections Can Slow Viral Evolution and Reduce the Risk of Treatment Failure. Sci. Rep. 2021, 11, 22630. [Google Scholar] [CrossRef]
- Rochman, N.D.; Wolf, Y.I.; Faure, G.; Mutz, P.; Zhang, F.; Koonin, E.V. Ongoing Global and Regional Adaptive Evolution of SARS-CoV-2. Proc. Natl. Acad. Sci. 2021, 118, e2104241118. [Google Scholar] [CrossRef]
- Scherer, E.M.; Babiker, A.; Adelman, M.W.; Allman, B.; Key, A.; Kleinhenz, J.M.; Langsjoen, R.M.; Nguyen, P.-V.; Onyechi, I.; Sherman, J.D.; et al. SARS-CoV-2 Evolution and Immune Escape in Immunocompromised Patients. New Engl. J. Med. 2022, 386, 2436–2438. [Google Scholar] [CrossRef]
- Oz, T.; Guvenek, A.; Yildiz, S.; Karaboga, E.; Tamer, Y.T.; Mumcuyan, N.; Ozan, V.B.; Senturk, G.H.; Cokol, M.; Yeh, P.; et al. Strength of Selection Pressure Is an Important Parameter Contributing to the Complexity of Antibiotic Resistance Evolution. Mol. Biol. Evol. 2014, 31, 2387–2401. [Google Scholar] [CrossRef]
- REX Consortium Heterogeneity of Selection and the Evolution of Resistance. Trends Ecol. Evol. 2013, 28, 110–118. [CrossRef]
- Huisman, J.S.; Scire, J.; Angst, D.C.; Li, J.; Neher, R.A.; Maathuis, M.H.; Bonhoeffer, S.; Stadler, T. Estimation and Worldwide Monitoring of the Effective Reproductive Number of SARS-CoV-2. eLife 2022, 11, e71345. [Google Scholar] [CrossRef]
- Yang, Z.-R.; Jiang, Y.-W.; Li, F.-X.; Liu, D.; Lin, T.-F.; Zhao, Z.-Y.; Wei, C.; Jin, Q.-Y.; Li, X.-M.; Jia, Y.-X.; et al. Efficacy of SARS-CoV-2 Vaccines and the Dose–Response Relationship with Three Major Antibodies: A Systematic Review and Meta-Analysis of Randomised Controlled Trials. Lancet Microbe 2023, 4, e236–e246. [Google Scholar] [CrossRef] [PubMed]
- Stoddard, M.; Yuan, L.; Sarkar, S.; Mangalaganesh, S.; Nolan, R.P.; Bottino, D.; Hather, G.; Hochberg, N.S.; White, L.F.; Chakravarty, A. Heterogeneity in Vaccinal Immunity to SARS-CoV-2 Can Be Addressed by a Personalized Booster Strategy. Vaccines 2023, 11, 806. [Google Scholar] [CrossRef]
- Van Egeren, D.; Stoddard, M.; Malakar, A.; Ghosh, D.; Acharya, A.; Mainuddin, S.; Majumdar, B.; Luo, D.; Nolan, R.; Joseph-McCarthy, D.; et al. No Magic Bullet: Limiting In-School Transmission in the Face of Variable SARS-CoV-2 Viral Loads. Front. Public Health 2022, 10, 941773. [Google Scholar] [CrossRef]
- Chaguza, C.; Hahn, A.M.; Petrone, M.E.; Zhou, S.; Ferguson, D.; Breban, M.I.; Pham, K.; Peña-Hernández, M.A.; Castaldi, C.; Hill, V.; et al. Accelerated SARS-CoV-2 Intrahost Evolution Leading to Distinct Genotypes during Chronic Infection. MedRxiv 2022. [Google Scholar] [CrossRef]
- Callaway, E. The Coronavirus Is Mutating—Does It Matter? Nature 2020, 585, 174–177. [Google Scholar] [CrossRef]
- Harris, R. Vaccines Could Drive The Evolution Of More COVID-19 Mutants. NPR 2021. [Google Scholar]
- WHO. The Effects of Virus Variants on COVID-19 Vaccines. Available online: https://www.who.int/news-room/feature-stories/detail/the-effects-of-virus-variants-on-covid-19-vaccines (accessed on 4 October 2022).
- Feuer, W. WHO Says Coronavirus Has Not Meaningfully Mutated to a More Lethal or Contagious Form. Available online: https://www.cnbc.com/2020/06/03/who-says-coronavirus-has-not-meaningfully-mutated-to-a-more-lethal-or-contagious-form.html (accessed on 4 October 2022).
- Reed, J. Anthony Fauci’s Boss on Why Things Could Be Much Better Soon. Available online: https://nymag.com/intelligencer/2020/07/anthony-faucis-boss-on-why-things-could-be-much-better-soon.html (accessed on 4 October 2022).
- Kennedy, D.A.; Read, A.F. Monitor for COVID-19 Vaccine Resistance Evolution during Clinical Trials. PLoS Biol. 2020, 18, e3001000. [Google Scholar] [CrossRef]
- Starr, T.N.; Greaney, A.J.; Hilton, S.K.; Ellis, D.; Crawford, K.H.D.; Dingens, A.S.; Navarro, M.J.; Bowen, J.E.; Tortorici, M.A.; Walls, A.C.; et al. Deep Mutational Scanning of SARS-CoV-2 Receptor Binding Domain Reveals Constraints on Folding and ACE2 Binding. Cell 2020, 182, 1295–1310. [Google Scholar] [CrossRef]
- Greaney, A.J.; Starr, T.N.; Gilchuk, P.; Zost, S.J.; Binshtein, E.; Loes, A.N.; Hilton, S.K.; Huddleston, J.; Eguia, R.; Crawford, K.H.D.; et al. Complete Mapping of Mutations to the SARS-CoV-2 Spike Receptor-Binding Domain That Escape Antibody Recognition. Cell Host Microbe 2021, 29, 44–57.e9. [Google Scholar] [CrossRef] [PubMed]
- Baum, A.; Fulton, B.O.; Wloga, E.; Copin, R.; Pascal, K.E.; Russo, V.; Giordano, S.; Lanza, K.; Negron, N.; Ni, M.; et al. Antibody Cocktail to SARS-CoV-2 Spike Protein Prevents Rapid Mutational Escape Seen with Individual Antibodies. Science 2020, 369, 1014–1018. [Google Scholar] [CrossRef] [PubMed]
- Begley, S. Experts Envision Two Scenarios If the New Coronavirus Isn’t Contained. Available online: https://www.statnews.com/2020/02/04/two-scenarios-if-new-coronavirus-isnt-contained/ (accessed on 4 October 2022).
- Aktipis, A.; Alcock, J. How the Coronavirus Escapes an Evolutionary Trade-Off That Helps Keep Other Pathogens in Check. Available online: http://theconversation.com/how-the-coronavirus-escapes-an-evolutionary-trade-off-that-helps-keep-other-pathogens-in-check-140706 (accessed on 4 October 2022).
- CDC. COVID Data Tracker. Available online: https://covid.cdc.gov/covid-data-tracker (accessed on 12 May 2021).
- Tian, D.; Sun, Y.; Xu, H.; Ye, Q. The Emergence and Epidemic Characteristics of the Highly Mutated SARS-CoV-2 Omicron Variant. J. Med. Virol. 2022, 94, 2376–2383. [Google Scholar] [CrossRef] [PubMed]
- Harvey, W.T.; Carabelli, A.M.; Jackson, B.; Gupta, R.K.; Thomson, E.C.; Harrison, E.M.; Ludden, C.; Reeve, R.; Rambaut, A.; Peacock, S.J.; et al. SARS-CoV-2 Variants, Spike Mutations and Immune Escape. Nat. Rev. Microbiol. 2021, 19, 409–424. [Google Scholar] [CrossRef] [PubMed]
- Sneppen, K.; Nielsen, B.F.; Taylor, R.J.; Simonsen, L. Overdispersion in COVID-19 Increases the Effectiveness of Limiting Nonrepetitive Contacts for Transmission Control. Proc. Natl. Acad. Sci. USA 2021, 118, e2016623118. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, B.F.; Eilersen, A.; Simonsen, L.; Sneppen, K. Lockdowns Exert Selection Pressure on Overdispersion of SARS-CoV-2 Variants. Epidemics 2022, 40, 100613. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Egeren, D.; Stoddard, M.; White, L.F.; Hochberg, N.S.; Rogers, M.S.; Zetter, B.; Joseph-McCarthy, D.; Chakravarty, A. Vaccines Alone Cannot Slow the Evolution of SARS-CoV-2. Vaccines 2023, 11, 853. https://doi.org/10.3390/vaccines11040853
Van Egeren D, Stoddard M, White LF, Hochberg NS, Rogers MS, Zetter B, Joseph-McCarthy D, Chakravarty A. Vaccines Alone Cannot Slow the Evolution of SARS-CoV-2. Vaccines. 2023; 11(4):853. https://doi.org/10.3390/vaccines11040853
Chicago/Turabian StyleVan Egeren, Debra, Madison Stoddard, Laura F. White, Natasha S. Hochberg, Michael S. Rogers, Bruce Zetter, Diane Joseph-McCarthy, and Arijit Chakravarty. 2023. "Vaccines Alone Cannot Slow the Evolution of SARS-CoV-2" Vaccines 11, no. 4: 853. https://doi.org/10.3390/vaccines11040853
APA StyleVan Egeren, D., Stoddard, M., White, L. F., Hochberg, N. S., Rogers, M. S., Zetter, B., Joseph-McCarthy, D., & Chakravarty, A. (2023). Vaccines Alone Cannot Slow the Evolution of SARS-CoV-2. Vaccines, 11(4), 853. https://doi.org/10.3390/vaccines11040853