
Using an Aluminum Hydroxide–Chitosan Matrix Increased the Vaccine Potential and Immune Response of Mice against Multi-Drug-Resistant Acinetobacter baumannii

 , , , , , , ,
, , , , , , ,  and
and
Abstract
1. Introduction
2. Materials and methods
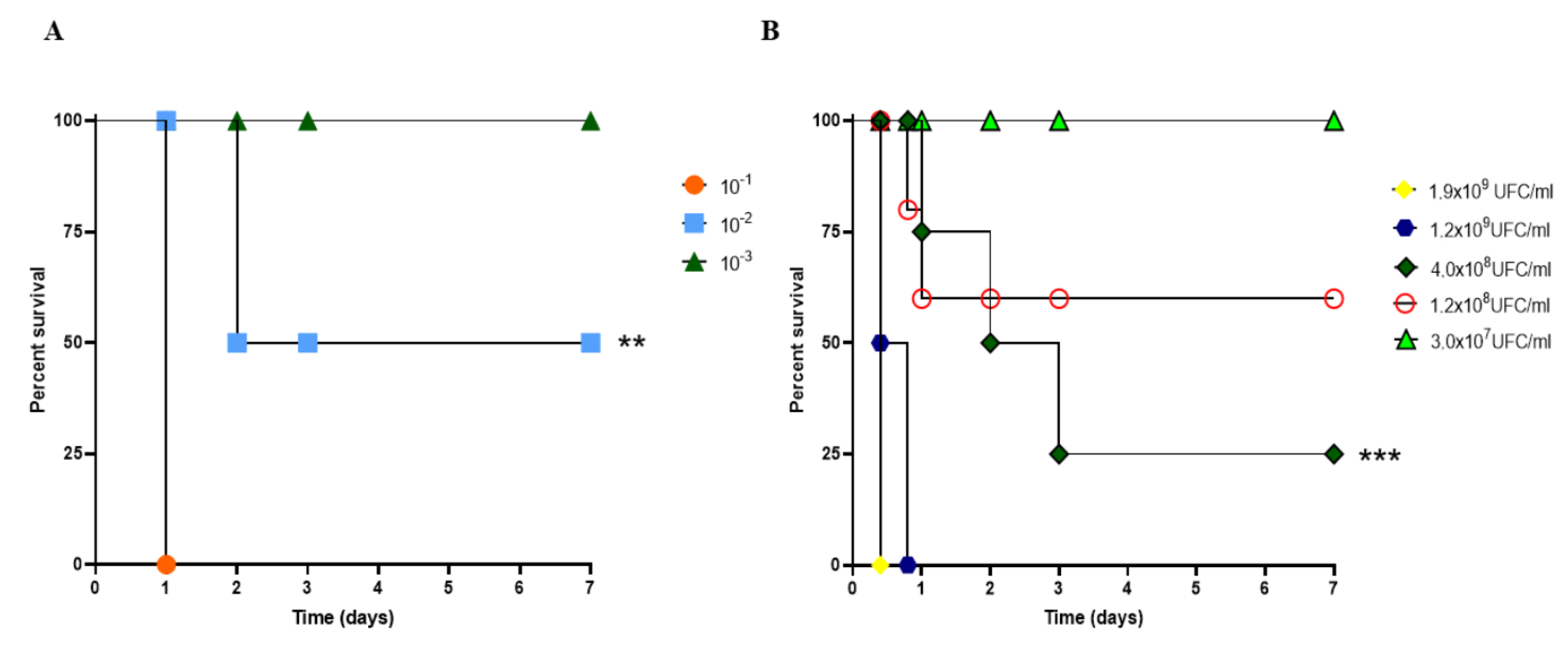
2.1. A. baumannii Strain and Lethal Dose Determination
2.2. Inactivation of A. baumannii Culture
2.3. mAhC and Vaccine Formulation
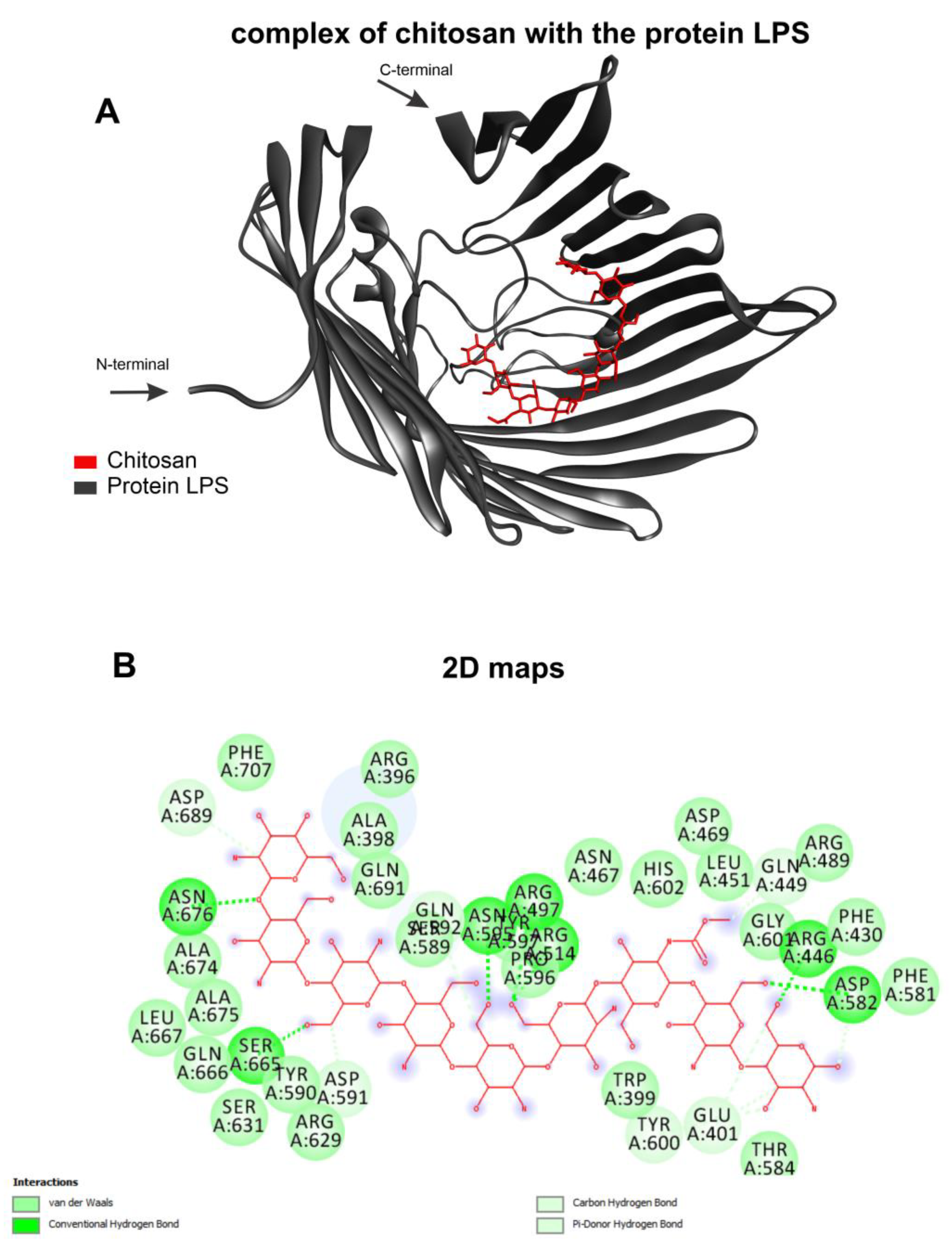
2.4. mAhC and Molecular Docking Calculations
2.5. Ethical/Legal Requirements and Use of Animals in Experiments
2.6. Experimental Design
2.7. Enzyme-Linked Immunosorbent Assay Using A. baumannii Cultures
2.8. Histopathological Analysis
2.9. Statistical Analysis
3. Results
3.1. Sepsis Model of Infection Using A. baumannii in CY-Untreated Mice
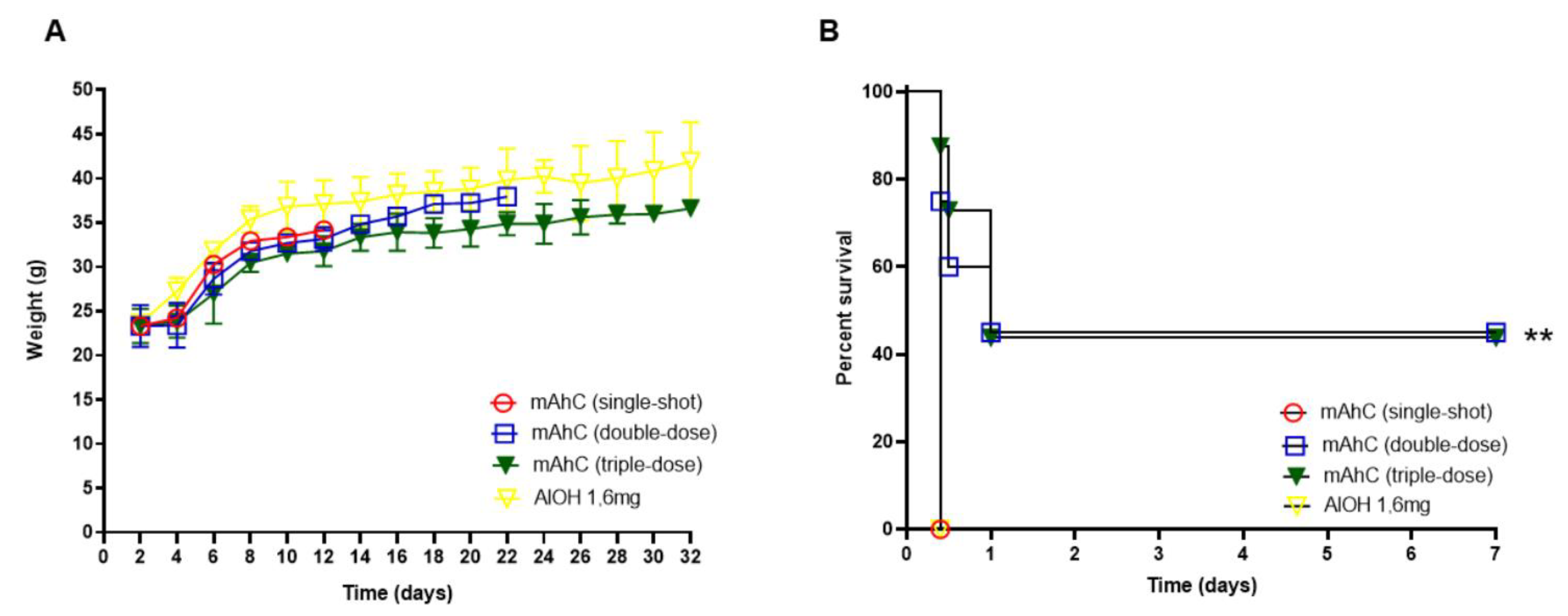
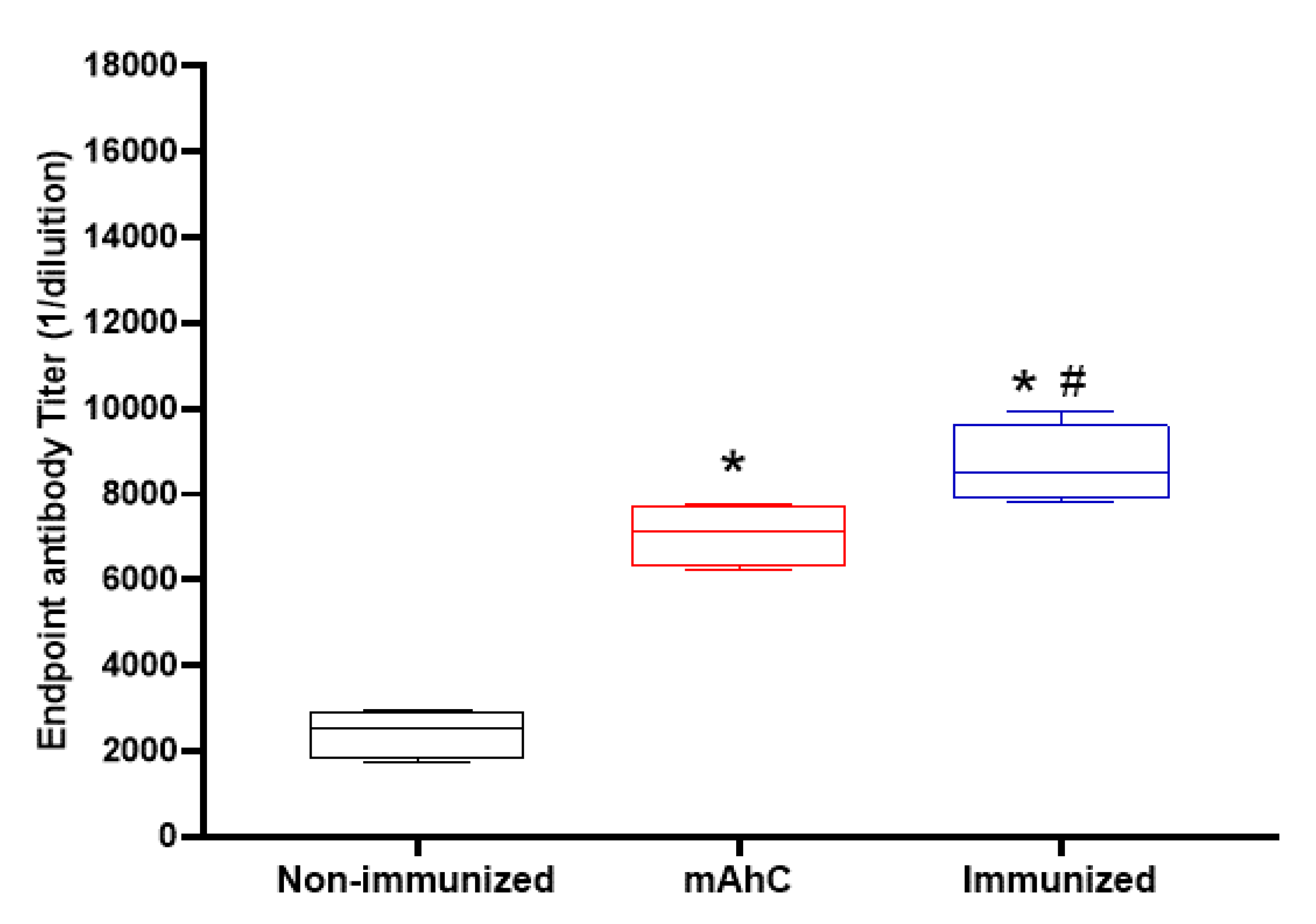
3.2. Determination of mAhC Dose in CY-Untreated Mice
3.3. Molecular Docking from MAhC and Bacterial Lipopolysaccharide
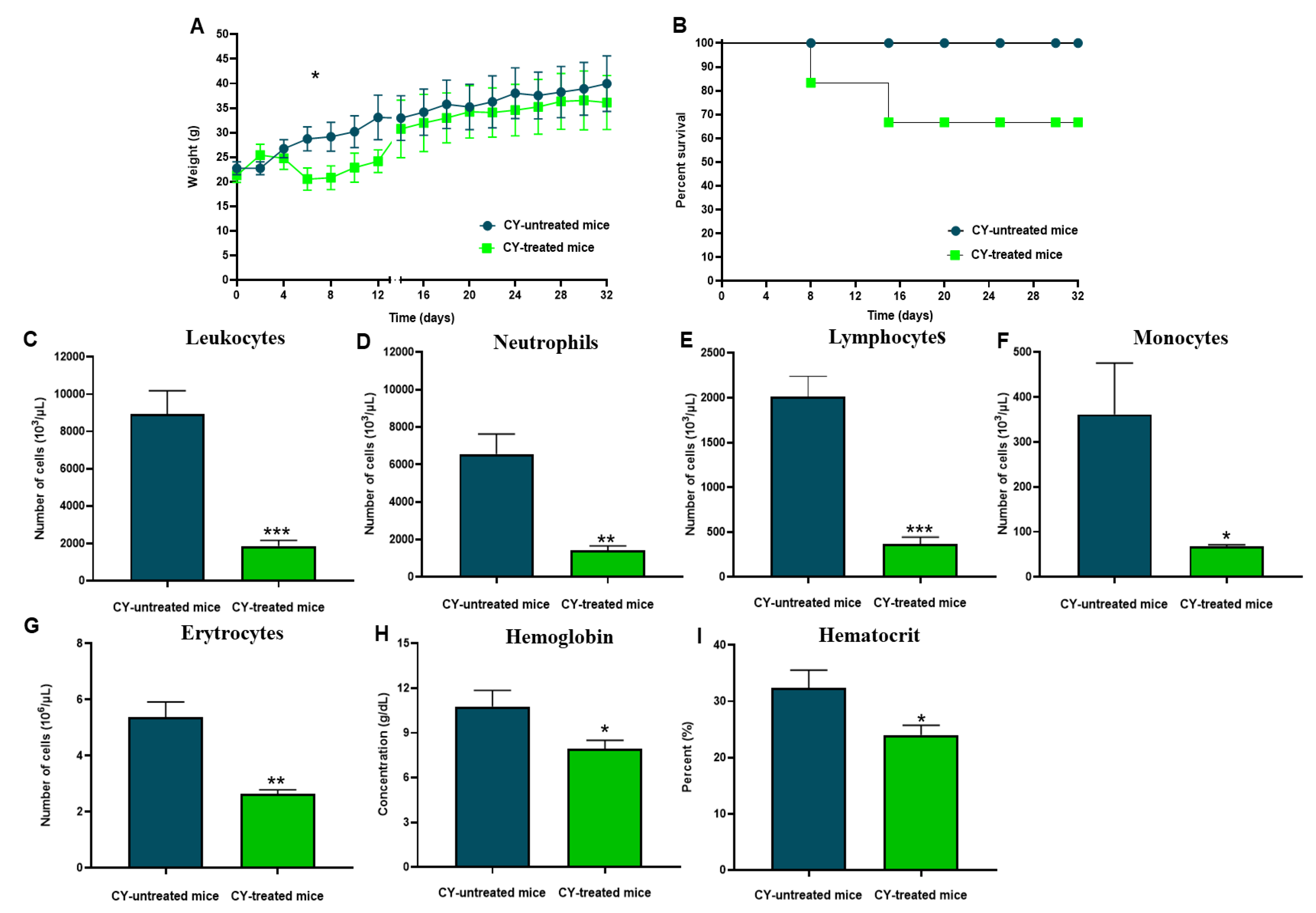
3.4. Immunosuppressive Model
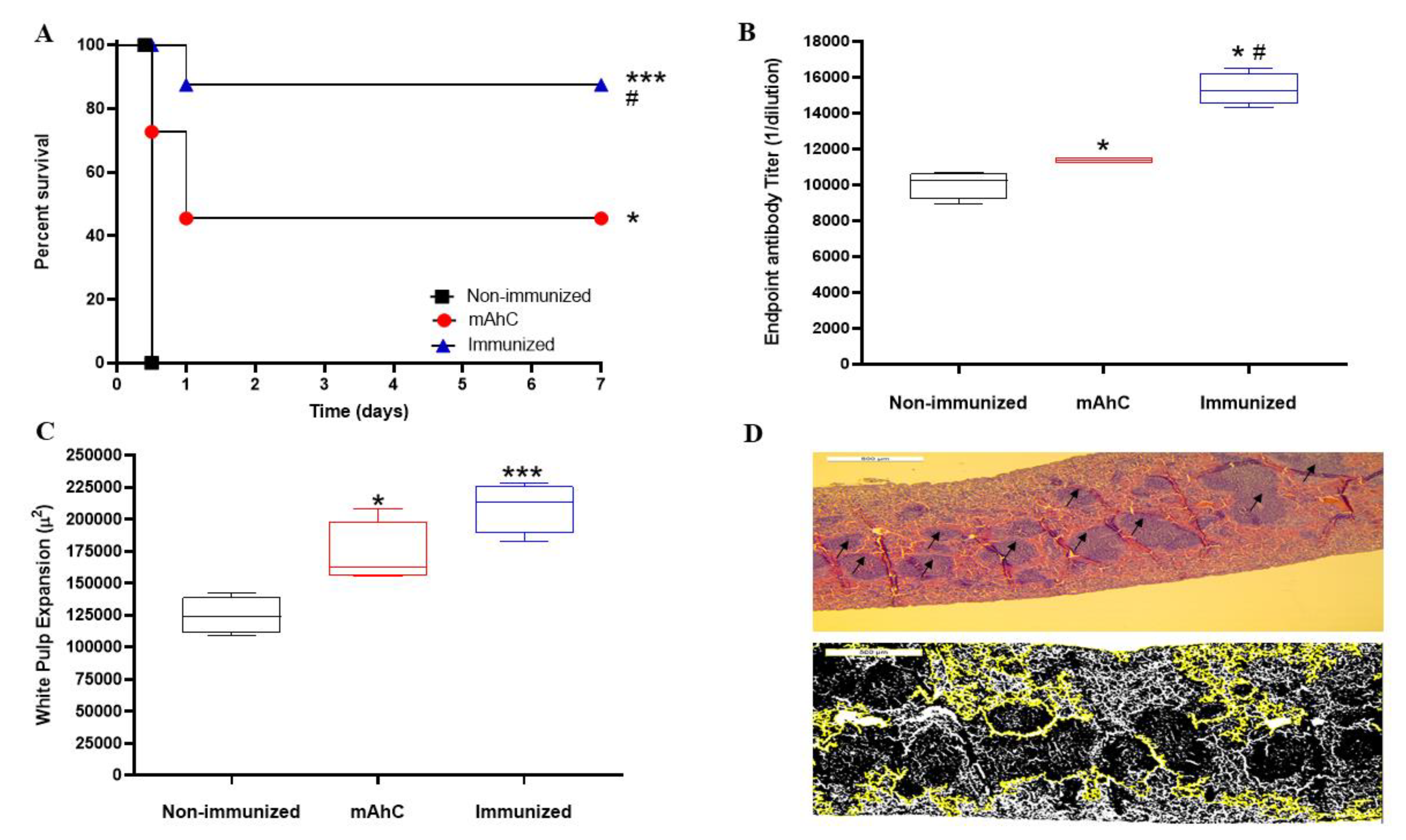
3.5. Protective Effect of A. baumannii Whole-Cell Vaccine Inactivated and Adsorbed on mAhC in CY-Treated Mice
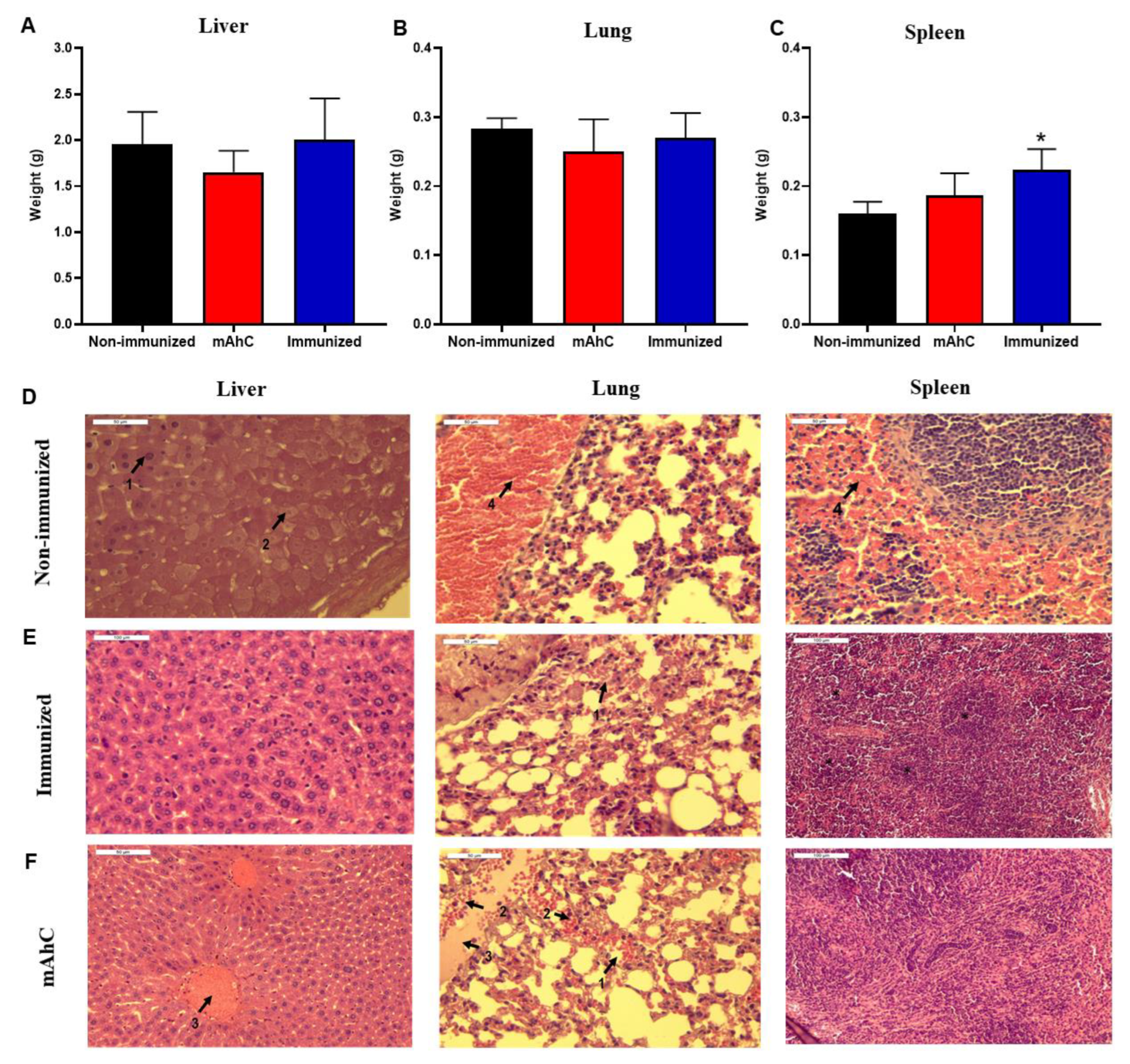
3.6. Histopathological Analysis of the Sepsis Model of A. baumannii Infection in CY-Treated Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Giamarellou, H.; Antoniadou, A.; Kanellakopoulou, K. Acinetobacter baumannii: A universal threat to public health? Int. J. Antimicrob. Agents 2008, 3, 106–119. [Google Scholar] [CrossRef] [PubMed]
- Lesho, E.; Eun-Jeong, Y.; McGann, P.; Snesrud, E.; Kwak, Y.; Milillo, M.; Onmus-Leone, F.; Preston, L.; St Clair, K.; Nikolich, M.; et al. Emergence of colistin-resistence in extremely drug-resistant Acinetobacter baumannii containing a novel pmrCAB operon during colistin therapy of wound infections. J. Infect. Dis. 2013, 208, 1142–1151. [Google Scholar] [CrossRef] [PubMed]
- Wong, D.; Nielsen, T.B.; Bonomo, R.A.; Pantapalangkoor, P.; Luna, B.; Spellberg, B. Clinical and pathophysiological overview of Acinetobacter infections: A century of challenges. Clin. Microbiol. Rev. 2017, 30, 409–447. [Google Scholar] [CrossRef] [PubMed]
- Castilho, S.R.A.; Godoy, C.S.D.M.; Guilarde, A.O.; Cardoso, J.L.; André, M.C.P.; Junqueira-Kipnis, A.P.; Kipnis, A. Acinetobacter baumannii strains isolated from patients in intensive care units in Goiania, Brazil: Molecular and drug susceptibility profiles. PLoS ONE 2017, 12, e0176790. [Google Scholar] [CrossRef] [PubMed]
- Munier, A.L.; Biard, L.; Legrand, M.; Rousseau, C.; Lafaurie, M.; Donay, J.L.; Molina, J.M. Incidence, risk factors and outcome of multi-drug resistant Acinetobacter baumannii nosocomial infections during an outbreak in a burn unit. International J. Infect. Dis. 2019, 79, 179–184. [Google Scholar] [CrossRef]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Monnet, D.L. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Zarrilli, R.; Pournaras, S.; Giannouli, M.; Tsakris, A. Global evolution of multidrug-resistant Acinetobacter baumannii clonal lineages. Int. J. Antimicrob. Agents 2013, 41, 11–19. [Google Scholar] [CrossRef]
- Mancuso, G.; Midiri, A.; Gerace, E.; Biondo, C. Bacterial Antibiotic Resistance: The Most Critical Pathogens. Pathogens 2021, 10, 1310. [Google Scholar] [CrossRef]
- Kyriakidis, I.; Vasileiou, E.; Pana, Z.D.; Tragiannidis, A. Acinetobacter baumannii Antibiotic Resistance Mechanisms. Pathogens 2021, 10, 373. [Google Scholar] [CrossRef]
- Lin, C.Y.; Chen, Y.M.; Lin, M.C.; Chang, Y.P.; Chao, T.Y.; Wang, C.C.; Tsai, Y.L.; Shen, L.S.; Li, C.L.; Lin, A.S. Risk factors of multidrug-resistant Acinetobacter baumannii recurrence after successful eradication in ventilated patients. Biomed. J. 2016, 39, 130–138. [Google Scholar] [CrossRef]
- Yang, Q.; Rui, Y. Two multiplex real-time PCR assays to detect and differentiate Acinetobacter baumannii and non-Baumannii Acinetobacter spp. carrying bla NDM, bla OXA-23-like, bla OXA-40-like, bla OXA-51-like, and bla OXA-58-like genes. PLoS ONE 2016, 11, e0158958. [Google Scholar] [CrossRef]
- Lee, C.R.; Lee, J.H.; Park, M.; Park, K.S.; Bae, I.K.; Kim, Y.B.; Cha, C.J.; Jeong, B.C.; Lee, S.H. Biology of Acinetobacter baumannii: Pathogenesis, Antibiotic Resistance Mechanisms, and Prospective Treatment Options. Front. Cell. Infect. Microbiol. 2017, 7, 55. [Google Scholar] [CrossRef] [PubMed]
- Brasil, Instrução normativa 23 M.A.P.A. Diário Oficial da União. Available online: https://www.in.gov.br/servicos/diario-oficial-da-uniao (accessed on 31 January 2023).
- Sobrinho, E.M.; Cangussu, A.S.R.; Brandi, I.V.; Sari, R.S.; Almeida, A.C.; Colen, F.; Quintilio, W.; Santos, H.O. Modified toxin-binding inhibition (ToBI) test for epsilon antitoxin determination in serum of immunized rabbits. Vet. Immunol. Immunopathol. 2010, 138, 129–133. [Google Scholar] [CrossRef] [PubMed]
- McConnell, M.J.; Actis, L.; Pachón, J. Acinetobacter baumannii: Human infections, factors contributing to pathogenesis and animal models. FEMS Microbiol. Rev. 2013, 37, 130–155. [Google Scholar] [CrossRef] [PubMed]
- Brandi, I.V.; Mozzer, O.D.; Vander, J.E.; Passos, F.J.V.; Passos, F.M.L.; Cangussu, A.S.R.; Sobrinho, E.M. Growth conditions of clostridium perfringens type B for production of toxins used to obtain veterinary vaccines. Bioprocess Biosyst. Eng. 2014, 37, 1737–1742. [Google Scholar] [CrossRef]
- Cangussu, A.S.R.; Mariúba, L.A.M.; Lalwani, P.; Pereira, K.D.; Astolphi-Filho, E.S.; Orlandi, P.P.; Nogueira, P.A. A hybrid protein containing MSP1a repeats and Omp7, Omp8 and Omp9 epitopes protect immunized BALB/c mice against anaplasmosis. Vet. Res. 2018, 49, 6. [Google Scholar] [CrossRef]
- Santos, E.M.S.; Almeida, A.C.; Santos, H.O.; Cangussu, A.S.R.; Costa, K.S.; Alves, J.N.; Barbosa, L.C.B.; Aguiar, R.W.S. Mechanism of Brassica oleracea performance in bovine infectious mastitis by bioinformatic analysis. Microb. Pathog. 2019, 129, 19–29. [Google Scholar] [CrossRef]
- Felix, M.K.C.; Deusdará, T.T.; Santos, L.S.S.; Aguiar, R.W.S.; Corrêa, R.F.T.; Brandi, I.V.; Cangussu, A.S.R. Inactivated alpha toxin from Clostridium novyi type B in nano-emulsion protect partially protects Swiss mice from lethal alpha toxin challenge. Sci. Rep. 2019, 9, 14082. [Google Scholar] [CrossRef]
- Brito, H.S.; Alencar, F.C.; Alburquerque, B.; Silva, M.G.; Félix, M.K.C.; Mulholland, D.S.; Oliveira, E.E.; Mariúba, L.A.M.; Santos, E.M.S.; Brandi, I.V.; et al. Pathologic findings on ruminant enteric clostridial diseases reveal specificities and differences among iota and iota-like toxins. Rev. Cienc. Agríc. 2021, 38, 157–174. [Google Scholar] [CrossRef]
- Chen, W. Host innate immune responses to Acinetobacter baumannii infection. Front. Cell. Infect. Microbiol. 2020, 10, 486. [Google Scholar] [CrossRef]
- Cabral, M.P.; García, P.; Beceiro, A.; Rumbo, C.; Pérez, A.; Moscoso, M.; Bou, G. Design of live attenuated bacterial vaccines based on D-glutamate auxotrophy. Nat. Commun. 2017, 8, 15480. [Google Scholar] [CrossRef]
- Dollery, S.J.; Zurawski, D.V.; Gaidamakova, E.K.; Matrosova, V.Y.; Tobin, J.K.; Wiggins, T.J.; Tobin, G.J. Radiation-Inactivated Acinetobacter baumannii Vaccine Candidates. Vaccines 2021, 9, 96. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.Y.; Hsieh, S.C.; Liu, C.W.; Lu, C.S.; Wu, C.H.; Liao, H.T.; Chen, M.H.; Li, K.J.; Shen, C.Y.; Kuo, Y.M.; et al. Cross-Talk among Polymorphonuclear Neutrophils, Immune, and Non-Immune Cells via Released Cytokines, Granule Proteins, Microvesicles, and Neutrophil Extracellular Trap Formation: A Novel Concept of Biology and Pathobiology for Neutrophils. Int. J. Mol. Sci. 2021, 22, 3119. [Google Scholar] [CrossRef]
- Diehl, R.; Ferrara, F.; Müller, C.; Dreyer, A.Y.; McLeod, D.D.; Fricke, S.; Boltze, J. Immunosuppression for in vivo research: State-of-the-art protocols and experimental approaches. Cell. Mol. Immunol. 2017, 14, 146–179. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.R.; Du, W.J.; Xie, J.F.; Xu, J.Y.; Huang, Y.Z.; Qiu, H.B.; Yang, Y. Role of immunodeficiency in Acinetobacter baumannii associated pneumonia in mice. Chin. Med. 2020, 133, 2161. [Google Scholar] [CrossRef] [PubMed]
- Ikezawa, Y.; Nakazawa, M.; Tamura, C.; Takahashi, K.; Minami, M.; Ikezawa, Z. Cyclophosphamide decreases the number, percentage and the function of CD25+ CD4+ regulatory T cells, which suppress induction of contact hypersensitivity. J. Dermatol. Sci. 2005, 39, 105–112. [Google Scholar] [CrossRef]
- McConnell, M.J.; Pachón, J. Active and passive immunization against Acinetobacter baumannii using an inactivated whole cell vaccine. Vaccine 2010, 29, 1–5. [Google Scholar] [CrossRef]
- Jin, P.; Sun, F.; Liu, Q.; Wang, Q.; Zhang, Y.; Liu, X. An oral vaccine based on chitosan/aluminum adjuvant induces both local and systemic immune responses in turbot (Scophthalmus maximus). Vaccine 2021, 39, 7477–7484. [Google Scholar] [CrossRef]
- Luo, G.; Lin, L.; Ibrahim, A.S.; Baquir, B.; Pantapalangkoor, P.; Bonomo, R.A.; Spellberg, B. Active and passive immunization protects against lethal, extreme drug resistant-Acinetobacter baumannii infection. PLoS ONE 2012, 7, e29446. [Google Scholar] [CrossRef]
- García-Quintanilla, M.; Pulido, M.R.; Pachón, J.; McConnell, M.J. Immunization with lipopolysaccharide-deficient whole cells provides protective immunity in an experimental mouse model of Acinetobacter baumannii infection. PLoS ONE 2014, 9, e114410. [Google Scholar] [CrossRef]
- Shu, M.H.; MatRahim, N.; NorAmdan, N.; Pang, S.P.; Hashim, S.H.; Phoon, W.H.; AbuBakar, S. An inactivated antibiotic-exposed whole-cell vaccine enhances bactericidal activities against multidrug-resistant Acinetobacter baumannii. Sci. Rep. 2016, 6, 22332. [Google Scholar] [CrossRef]
- Ramezanalizadeh, F.; Owlia, P.; Rasooli, I. Type I pili, CsuA/B and FimA induce a protective immune response against Acinetobacter baumannii. Vaccine 2020, 38, 5436–5446. [Google Scholar] [CrossRef] [PubMed]
- Sari, R.S.; Almeida, A.C.; Cangussu, A.S.R.; Jorge, E.V.; Mozzer, D.O.; Santos, H.O.; Quintilio, W.; Brandi, I.V.; Andrade, V.A.; Miguel, A.S.M.; et al. Anti-botulism single-shot vaccine using chitosan for protein encapsulation by simple coacervation. Anaerobe 2016, 42, 182–187. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, 296–303. [Google Scholar] [CrossRef]
- Ramachandran, G.N.; Sasisekharan, V. Conformation of polypeptides and proteins. Adv. Protein Chem. 1968, 23, 283–437. [Google Scholar] [CrossRef] [PubMed]
- Haas, J.; Barbato, A.; Behringer, D.; Studer, G.; Roth, S.; Bertoni, M.; Mostaguir, K.; Gumienny, R.; Schwede, T. Continuous Automated Model Evaluation (CAMEO) complementing the critical assessment of structure prediction in CASP12. Proteins 2018, 86, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Benkert, P.; Biasini, M.; Schwede, T. Toward the estimation of the absolute quality of individual protein structure models. Bioinformatics 2011, 27, 343–350. [Google Scholar] [CrossRef]
- Sanner, M.F. Python: A programming language for software integration and development. J. Mol. Graph. Model. 1999, 17, 57–61. [Google Scholar]
- Moura, W.S.; Souza, S.R.; Campos, F.S.; Cangussu, A.S.R.; Santos, E.M.S.; Andrade, B.S.; Viana, K.F.; Haddi, K.; Oliveira, E.E.; Nascimento, V.L.; et al. Antibacterial activity of Siparuna guianensis essential oil mediated by impairment of membrane permeability and replication of pathogenic bacteria. Ind. Crops Prod. 2020, 46, 112142. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Schrodinger, LLC. The PyMOL Molecular Graphics System, Version 2.0; Schrodinger, LLC: New York, NY, USA, 2018. [Google Scholar]
- Biovia, D.S. Discovery Studio Modeling Environment, Release 2017; Dassault Systemes: San Diego, CA, USA, 2017. [Google Scholar]
- Papadopulos, F.; Spinelli, M.; Valente, S.; Foroni, L.; Orrico, C.; Pasquinelli, G. Common tasks in microscopic and ultrastructural image analysis using imageJ. Ultrastruct. Pathol. 2007, 31, 401–407. [Google Scholar] [CrossRef]
- Alos, J.I. Antibiotic resistance: A global crisis. Enferm. Infecc. Microbiol. Clin. 2014, 33, 692–699. [Google Scholar] [CrossRef]
- Borek, A.J.; Wanat, M.; Sallis, A.; Ashiru-Oredope, D.; Atkins, L.; Beech, E.; Tonkin-Crine, S. How can national antimicrobial stewardship interventions in primary care be improved? A stakeholder consultation. Antibiotics 2019, 8, 207. [Google Scholar] [CrossRef]
- Miranda, C.; Silva, V.; Igrejas, P.G.P. Impact of European pet antibiotic use on enterococci and staphylococci antimicrobial resistance and human health. Fut. Microbiol. 2021, 16, 185–203. [Google Scholar] [CrossRef]
- Rodríguez-Hernández, M.J.; Pachón, J.; Pichardo, C.; Cuberos, L.; Ibáñez-Martínez, J.; García-Curiel, A.; Jiménez-Mejías, M.E. Imipenem, doxycycline and amikacin in monotherapy and in combination in Acinetobacter baumannii experimental pneumonia. J. Antimicrob. Chemother. 2000, 45, 493–501. [Google Scholar] [CrossRef]
- Harris, G.; KuoLee, R.; Lam, C.K.; Kanzaki, G.; Patel, G.B.; Xu, H.H.; Chen, W. A mouse model of Acinetobacter baumannii-associated pneumonia using a clinically isolated hypervirulent strain. Antimicrob. Agents Chemother. 2013, 57, 3601–3613. [Google Scholar] [CrossRef] [PubMed]
- Suttie, A.W. Histopathology of the spleen. Toxicol. Pathol. 2006, 34, 466–503. [Google Scholar] [CrossRef] [PubMed]
- Elmore, S.A. Enhanced histopathology of the spleen. Toxicol. Pathol. 2006, 34, 648–655. [Google Scholar] [CrossRef] [PubMed]
- He, S.; He, H.; Chen, Y.; Chen, Y.; Wang, W.; Yu, D. In vitro and in vivo analysis of antimicrobial agents alone and in combination against multi-drug resistant Acinetobacter baumannii. Front. Microbiol. 2015, 6, 507. [Google Scholar] [CrossRef]
- Na, S.H.; Oh, M.H.; Jeon, H.; Lee, Y.K.; Lee, B.; Shin, M.; Lee, J.C. Imaging of bioluminescent Acinetobacter baumannii in a mouse pneumonia model. Microb. Pathog. 2019, 137, 103784. [Google Scholar] [CrossRef]
- Ketter, P.M.; Guentzel, M.N.; Schaffer, B.; Herzig, M.; Wu, X.; Montgomery, R.K.; Arulanandam, B.P. Severe Acinetobacter baumannii sepsis is associated with elevation of pentraxin 3. Infect. Immun. 2014, 82, 3910–3918. [Google Scholar] [CrossRef]
- KuoLee, R.; Harris, G.; Yan, H.; Xu, H.H.; Conlan, W.J.; Patel, G.B.; Chen, W. Intranasal immunization protects against Acinetobacter baumannii-associated pneumonia in mice. Vaccine 2015, 33, 260–267. [Google Scholar] [CrossRef]
- Bergamini, G.; Perico, M.E.; Di Palma, S.; Sabatini, D.; Andreetta, F.; Defazio, R.; Ferrari, L. Mouse pneumonia model by Acinetobacter baumannii multidrug resistant strains: Comparison between intranasal inoculation, intratracheal instillation and oropharyngeal aspiration techniques. PLoS ONE 2021, 16, e0260627. [Google Scholar] [CrossRef] [PubMed]
- Gilavand, F.; Marzban, A.; Ebrahimipour, G.; Soleimani, N.; Goudarzi, M. Designation of chitosan nano-vaccine based on MxiH antigen of Shigella flexneri with increased immunization capacity. Carbohydr. Polym. 2020, 232, 115813. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Xue, J.; Jiang, M.; Lin, S.; Huang, Y.; Deng, K.; Feng, G. A multiepitope peptide, rOmp22, encapsulated in chitosan-PLGA nanoparticles as a candidate vaccine against Acinetobacter baumannii infection. Int. J. Nanomed. 2021, 16, 1819. [Google Scholar] [CrossRef] [PubMed]
- Pulido, M.R.; García-Quintanilla, M.; Pachón, J.; McConnell, M.J. A Lipopolysaccharide-free outer membrane vesicle vaccine protects against Acinetobacter baumannii infection. Vaccine 2020, 38, 719–724. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism | Target (Unipot Database) | Template | Identity (%) | Ramachandran Favored (%) | QMEAN |
|---|---|---|---|---|---|
| A. baumannii | LPS (D0C7T1) | 5IV8 | 34.02% | 90.54% | 0.56 |
| Organism | Ligand | Affinity Energy (kcal/mol) |
|---|---|---|
| A. baumannii | Chitosan | −8.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deusdará, T.T.; Félix, M.K.C.; de S. Brito, H.; Cangussu, E.W.S.; de S. Moura, W.; Albuquerque, B.; Silva, M.G.; dos Santos, G.R.; de Morais, P.B.; da Silva, E.F.; et al. Using an Aluminum Hydroxide–Chitosan Matrix Increased the Vaccine Potential and Immune Response of Mice against Multi-Drug-Resistant Acinetobacter baumannii. Vaccines 2023, 11, 669. https://doi.org/10.3390/vaccines11030669
Deusdará TT, Félix MKC, de S. Brito H, Cangussu EWS, de S. Moura W, Albuquerque B, Silva MG, dos Santos GR, de Morais PB, da Silva EF, et al. Using an Aluminum Hydroxide–Chitosan Matrix Increased the Vaccine Potential and Immune Response of Mice against Multi-Drug-Resistant Acinetobacter baumannii. Vaccines. 2023; 11(3):669. https://doi.org/10.3390/vaccines11030669
Chicago/Turabian StyleDeusdará, Túllio T., Mellanie K. C. Félix, Helio de S. Brito, Edson W. S. Cangussu, Wellington de S. Moura, Benedito Albuquerque, Marcos G. Silva, Gil R. dos Santos, Paula B. de Morais, Elizangela F. da Silva, and et al. 2023. "Using an Aluminum Hydroxide–Chitosan Matrix Increased the Vaccine Potential and Immune Response of Mice against Multi-Drug-Resistant Acinetobacter baumannii" Vaccines 11, no. 3: 669. https://doi.org/10.3390/vaccines11030669
APA StyleDeusdará, T. T., Félix, M. K. C., de S. Brito, H., Cangussu, E. W. S., de S. Moura, W., Albuquerque, B., Silva, M. G., dos Santos, G. R., de Morais, P. B., da Silva, E. F., Chaves, Y. O., Mariúba, L. A. M., Nogueira, P. A., Astolfi-Filho, S., Assunção, E. N., Epiphanio, S., Marinho, C. R. F., Brandi, I. V., Viana, K. F., ... Cangussu, A. S. R. (2023). Using an Aluminum Hydroxide–Chitosan Matrix Increased the Vaccine Potential and Immune Response of Mice against Multi-Drug-Resistant Acinetobacter baumannii. Vaccines, 11(3), 669. https://doi.org/10.3390/vaccines11030669