

Mathematical Modeling and Analysis of the Dynamics of RNA Viruses in Presence of Immunity and Treatment: A Case Study of SARS-CoV-2

 ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
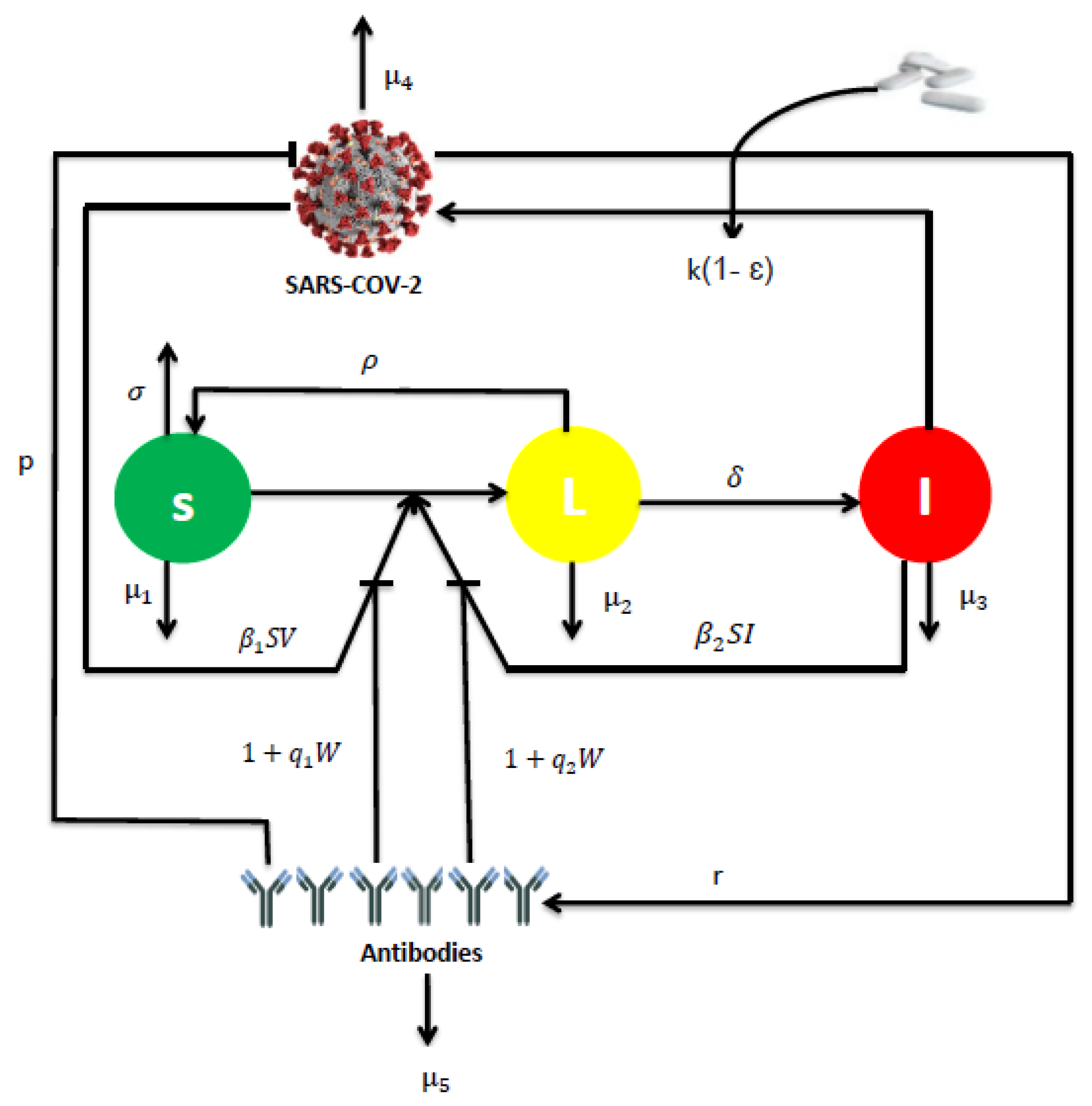
2.1. Mathematical Formulation
2.2. Equilibria and Threshold Parameters
- If , then , and
- (i)
- (ii)
- The threshold parameter is called the basic reproduction number. Biologically, this threshold parameter represents the average number of secondary infections produced by one productively infected cell at the beginning of infection. It can be rewritten as , where is the basic reproduction number of the virus-to-cell transmission mode and is the basic reproduction number of the cell-to-cell transmission mode.When , model (1) has another biological equilibrium called the infection equilibrium without humoral immunity of the form , where , , and .
- Since , we have . This implies that there is no biological equilibrium when or . Let and be the function defined on the closed interval as followswhere . On the other hand, we have andWhen the humoral immune response has not been established, we have . Hence, we define another threshold parameter called the reproduction number for humoral immunity as followswhere is the average life span of antibodies and is the quantity of viruses at the steady state . So, the number can biologically determine the average number of antibodies activated by viral particles.As if , we deduce that there exists a such that . Further, we have . This establishes the uniqueness of and therefore model (1) has an unique infection equilibrium point with humoral immunity when , where , , and .
3. Analytical Results
3.1. Well-Posedness
3.2. Stability Analysis
- (i)
- is globally asymptotically stable if and .
- (ii)
- is globally asymptotically stable if and δ is sufficiently large.
4. Numerical Results
4.1. Parameters Estimation
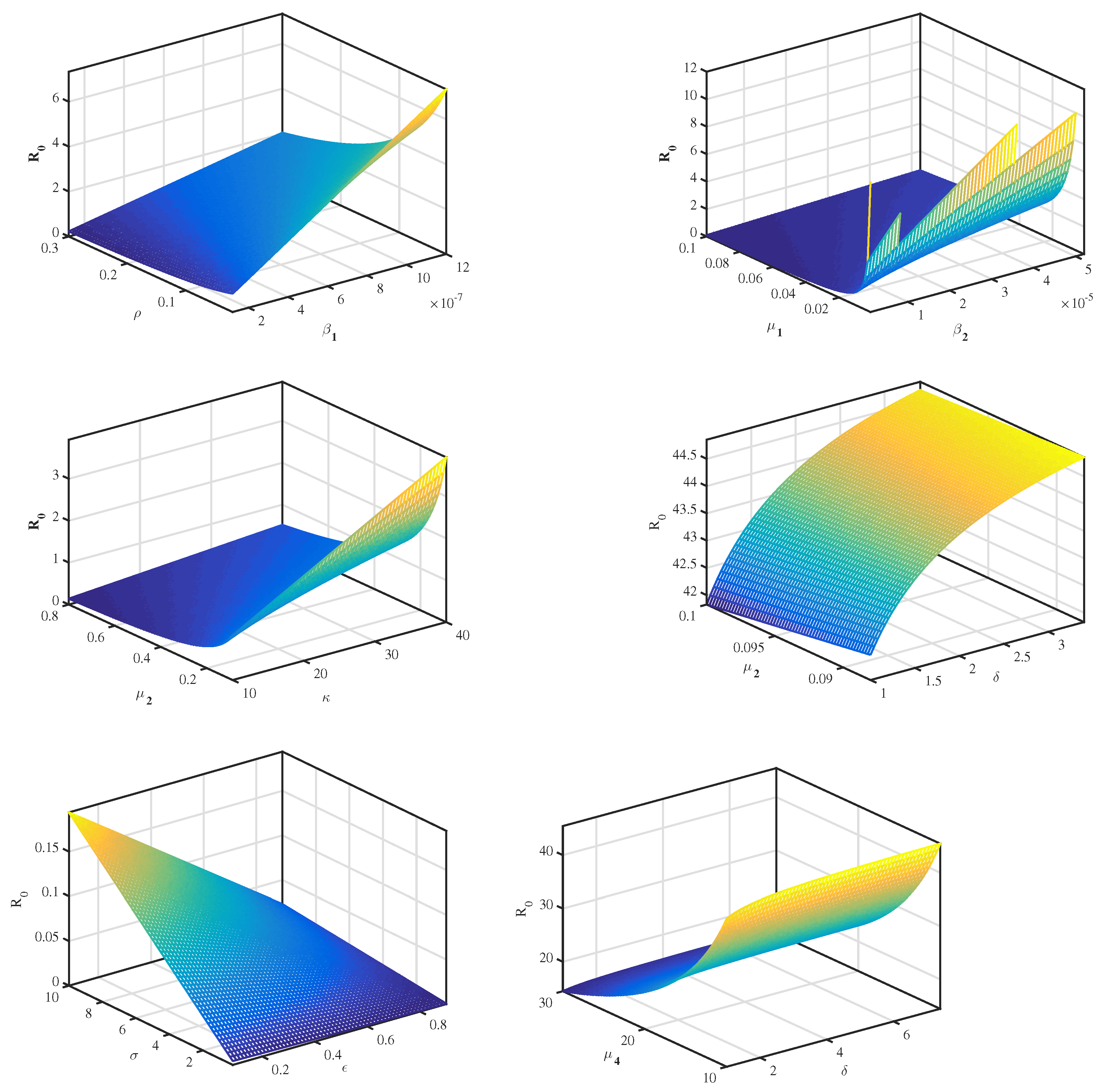
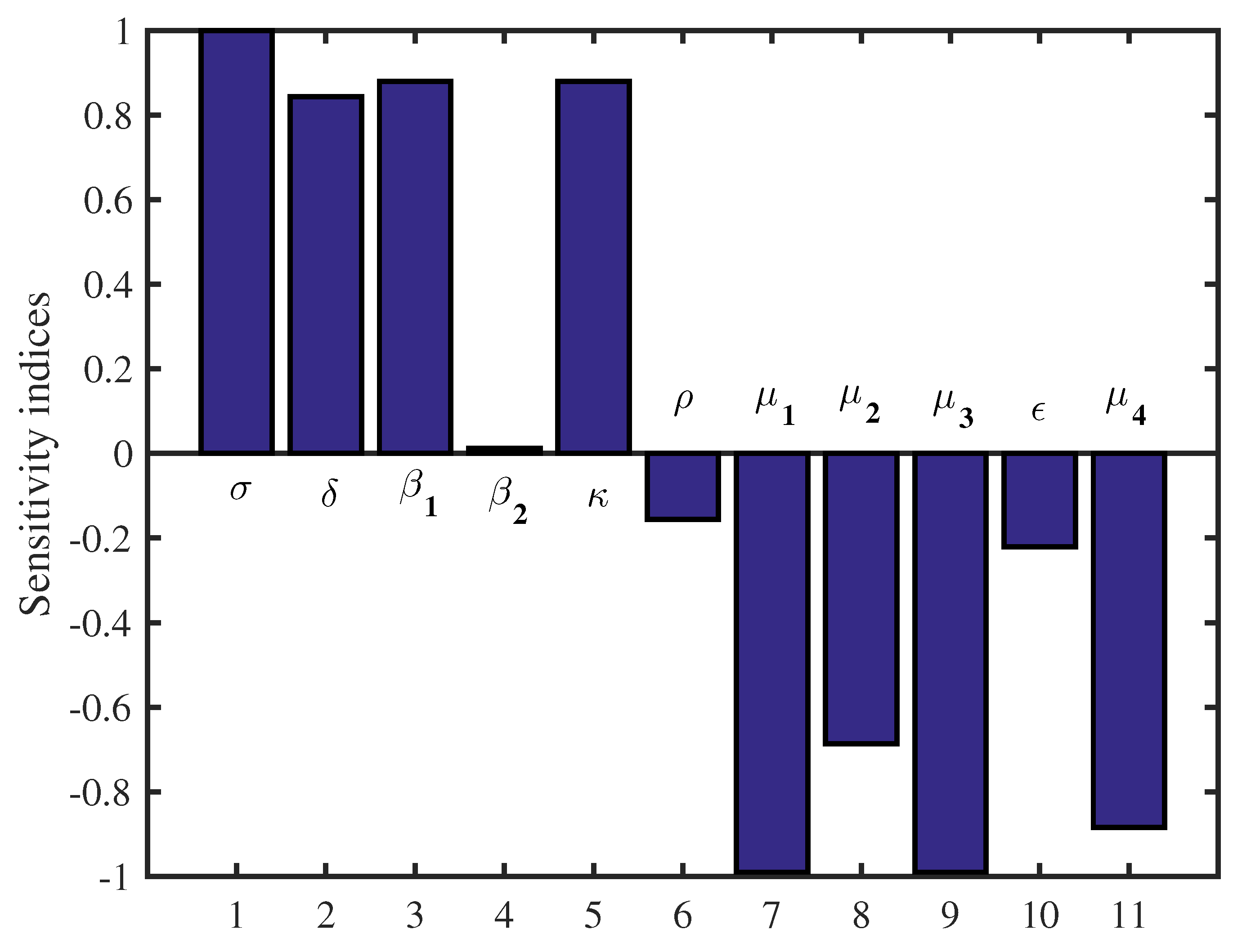
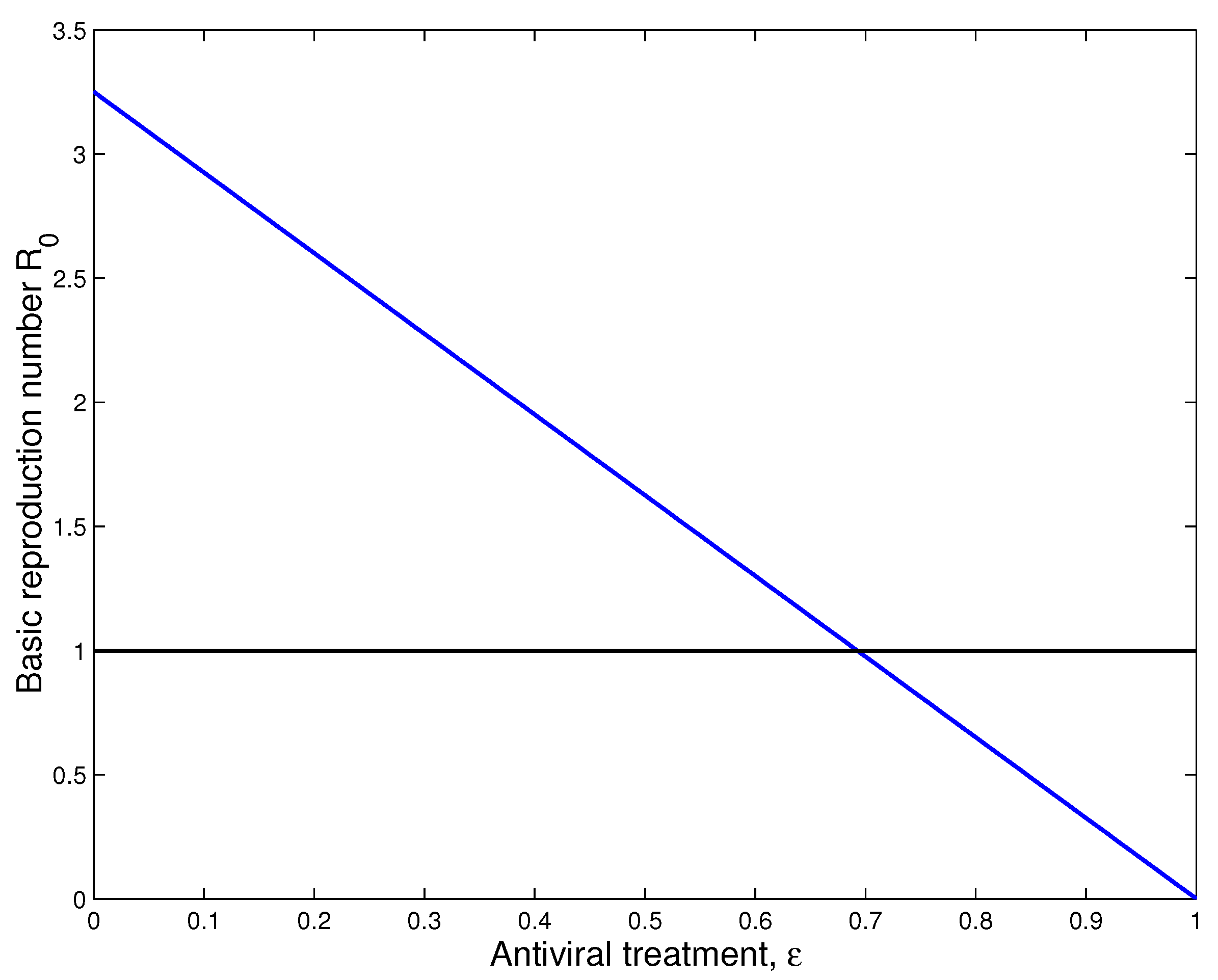
4.2. Sensitivity Analysis
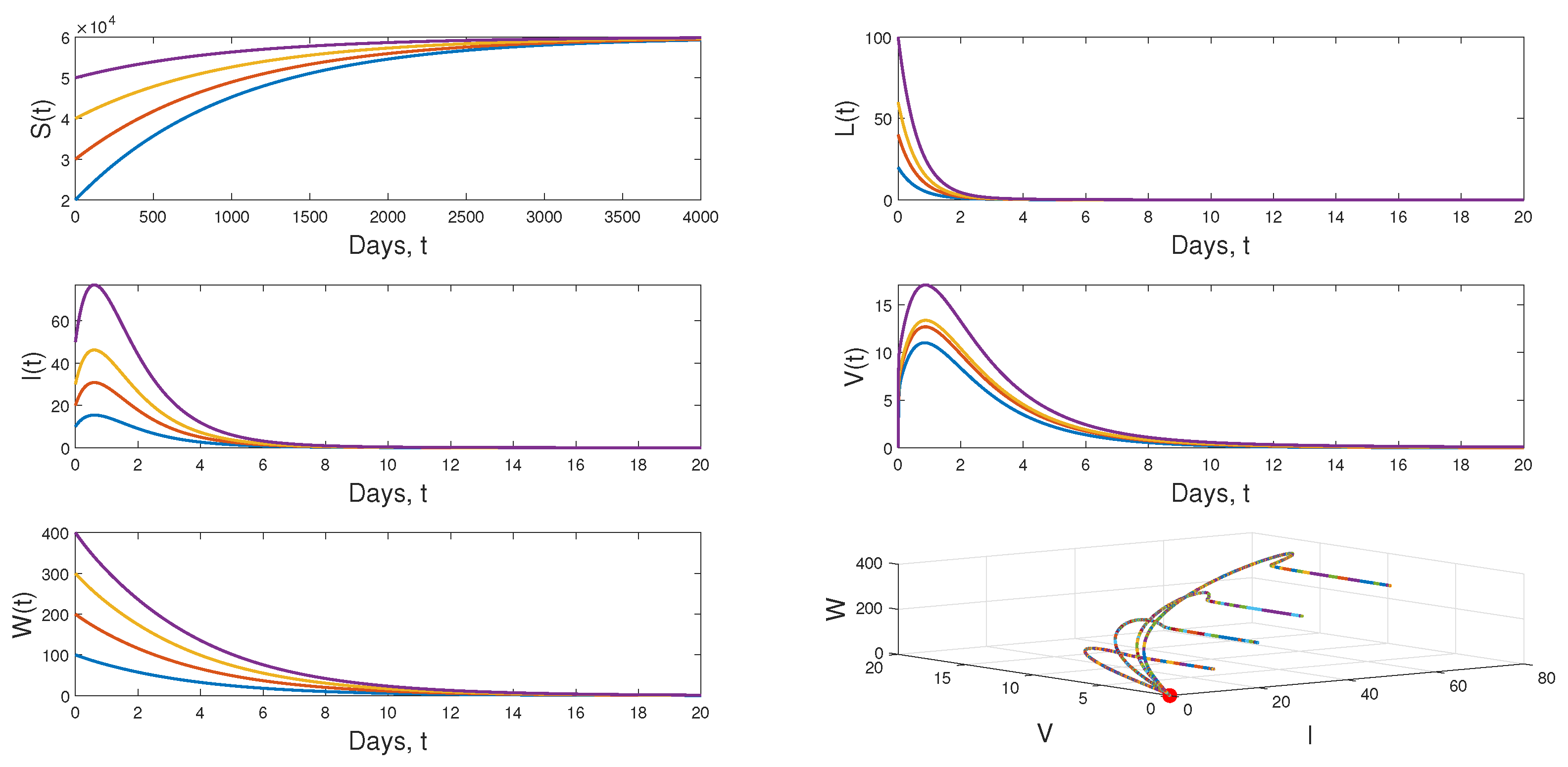
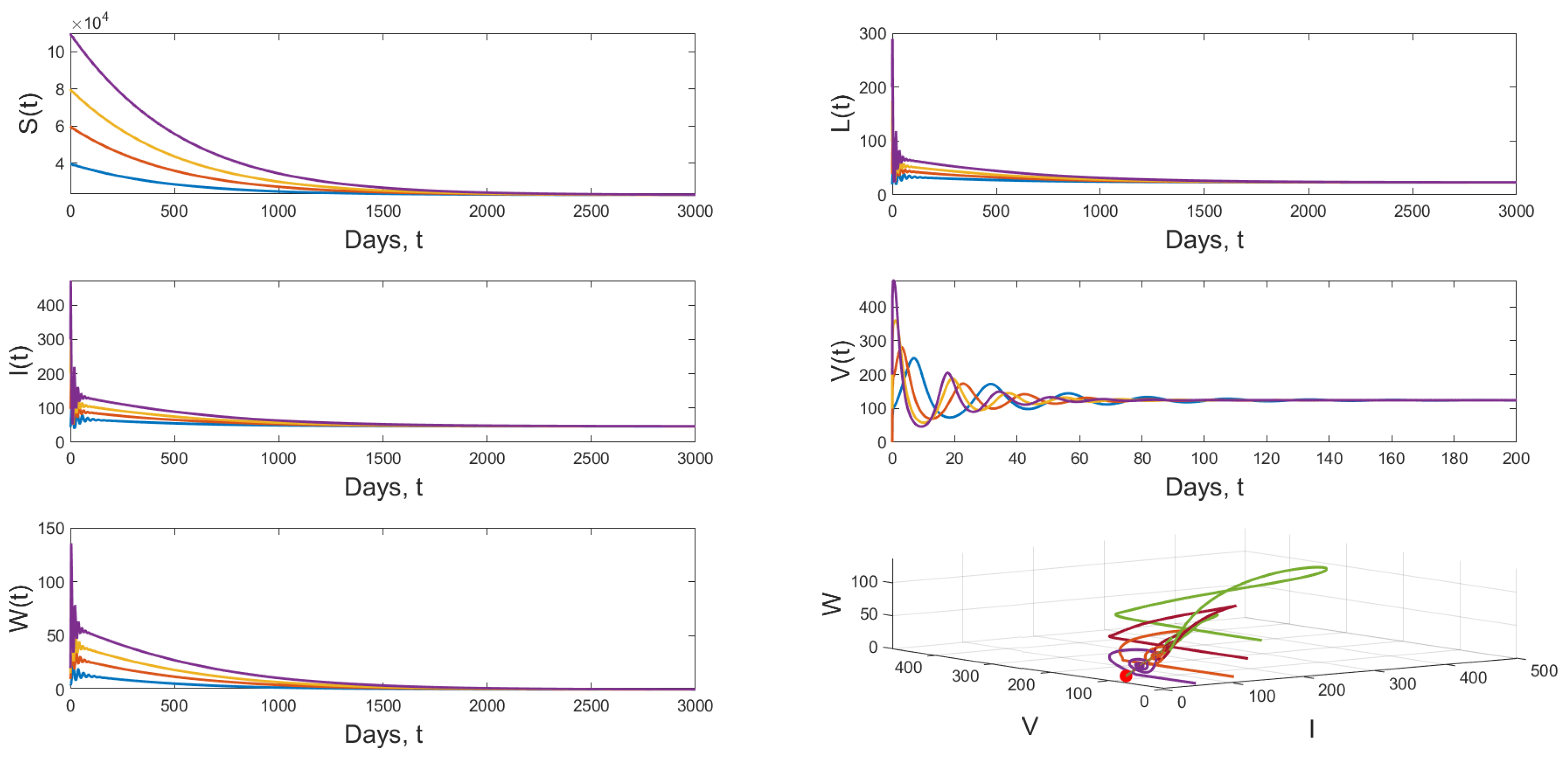
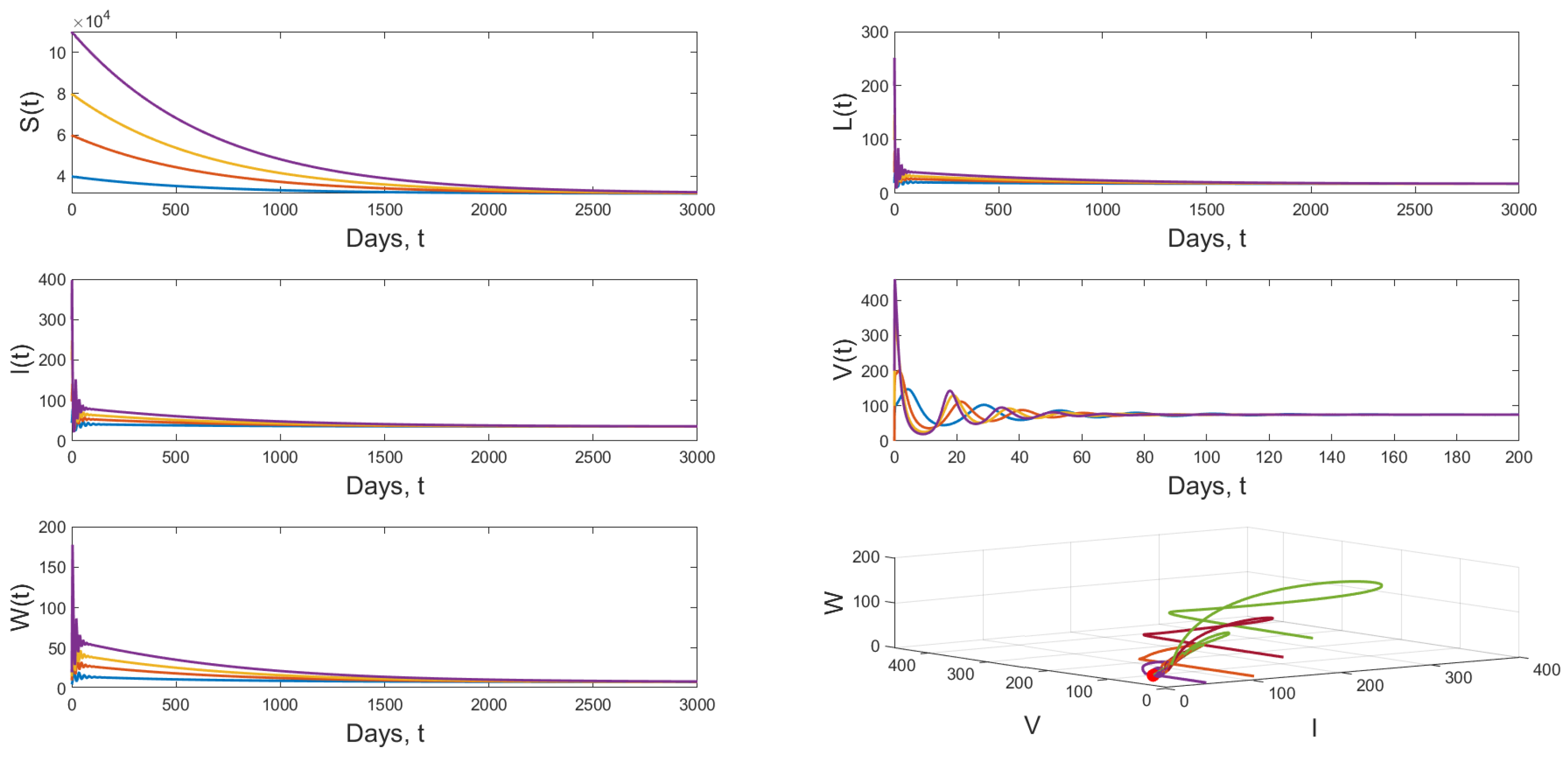
4.3. Numerical Simulation
5. Conclusions and Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- ICTV. The species Severe acute respiratory syndromerelated coronavirus: Classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar] [CrossRef]
- HWO. Weekly Epidemiological Update on COVID-19, 21 September 2022. Available online: https://www.who.int/publications/m/item/weekly-epidemiological-update-on-covid-19 (accessed on 21 September 2021).
- Mahase, E. Covid-19: Pfizer’s paxlovid is 89% effective in patients at risk of serious illness, company reports. Br. Med. J. (BMJ) 2021, 375, n2713. [Google Scholar] [CrossRef] [PubMed]
- Hattaf, K.; Mohsen, A.A.; Harraq, J.; Achtaich, N. Modeling the dynamics of COVID-19 with carrier effect and environmental contamination. Int. J. Model. Simul. Sci. Comput. 2021, 12, 2150048. [Google Scholar] [CrossRef]
- Hattaf, K.; Mohsen, A.A. Dynamics of a generalized fractional epidemic model of COVID-19 with carrier effect. Adv. Syst. Sci. Appl. 2022, 22, 36–48. [Google Scholar]
- Mohsen, A.A.; Al-Husseiny, H.F.; Zhou, X.; Hattaf, K. Global stability of COVID-19 model involving the quarantine strategy and media coverage effects. AIMS Public Health J. 2020, 7, 587–605. [Google Scholar] [CrossRef] [PubMed]
- Denu, D.; Kermausuor, S. Analysis of a Fractional-Order COVID-19 Epidemic Model with Lockdown. Vaccines 2022, 10, 1773. [Google Scholar] [CrossRef]
- DarAssi, M.H.; Safi, M.A.; Khan, M.A.; Beigi, A.; Aly, A.A.; Alshahrani, M.Y. A mathematical model for SARS-CoV-2 in variable-order fractional derivative. Eur. Phys. J. Spec. Top. 2022, 231, 1905–1914. [Google Scholar] [CrossRef]
- Hattaf, K.; Yousfi, N. Dynamics of SARS-CoV-2 infection model with two modes of transmission and immune response. Math. Biosci. Eng. 2020, 5, 5326–5340. [Google Scholar] [CrossRef]
- Chatterjee, A.N.; Basir, F.A.; Almuqrin, M.A.; Mondal, J. SARS-CoV-2 infection with lytic and non-lytic immune responses: A fractional order optimal control theoretical study. Results Phys. 2021, 26, 104260. [Google Scholar] [CrossRef]
- Chatterjee, A.N.; Basir, F.A. A model for SARS-CoV-2 infection with treatment. Comput. Math. Methods Med. 2020, 2020, 1–11. [Google Scholar] [CrossRef]
- Liu, X.; Wang, J.; Xu, X.; Liao, G.; Chen, Y.; Hu, C.H. Patterns of IgG and IgM antibody response in COVID-19 patients. Emerg. Microbes Infect. 2020, 9, 1269–1274. [Google Scholar] [CrossRef] [PubMed]
- Assadiasl, S.; Fatahi, Y.; Zavvard, M.; Nicknam, M.H. COVID-19: Significance of antibodies. Hum. Antibodies 2020, 8, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Valdez-Cruz, N.A.; Garcia-Hernandez, E.; Espitia, C.; Cobos-Marin, L.; Altamirano, C.; Bando-Campos, C.G.; Trujillo-Roldan, M.A. Integrative overview of antibodies against SARS-CoV-2 and their possible applications in COVID-19 prophylaxis and treatment. Microb. Cell Factories 2021, 20, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Seow, J.; Graham, C.; Merrick, B.; Acors, S.; Pickering, S.; Steel, K.J.; Doores, K.J. antibody responses in the three months following SARS-CoV-2 infection in humans. Nat. Microbiol. 2020, 5, 1598–1607. [Google Scholar] [CrossRef] [PubMed]
- Cervia, C.; Nilsson, J.; Zurbuchen, Y.; Valaperti, A.; Schreiner, J.; Wolfensberger, A.; Raeber, M.E.; Adamo, S.; Weigang, S.; Emmenegger, M.; et al. Systemic and mucosal antibody responses specific to SARS-CoV-2 during mild versus severe COVID-19. J. Allergy Clin. Immunol. 2021, 147, 545–557. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, L.; Sang, L.; Ye, F.; Ruan, S.; Zhong, B.; Song, T.; Alshukairi, A.N.; Chen, R.; Zhang, Z.; et al. Kinetics of viral load and antibody response in relation to COVID-19 severity. J. Clin. Investig. 2020, 130, 5235–5244. [Google Scholar] [CrossRef]
- Pang, N.Y.L.; Pang, A.S.R.; Chow, V.T.; Wang, D.Y. Understanding neutralising antibodies against SARS.CoV.2 and their implications in clinical practice. Mil. Med. Res. 2021, 8, 1–17. [Google Scholar]
- Deans, J.A.; Cohen, S. Immunology of malaria. Annu. Rev. Microbiol. 1983, 37, 25–50. [Google Scholar] [CrossRef]
- Pan, S.; Chakrabarty, S.P. Threshold dynamics of HCV model with cell-to-cell transmission and a non-cytolytic cure in the presence of humoral immunity. Commun. Nonlinear Sci. Numer. Simul. 2018, 61, 180–197. [Google Scholar] [CrossRef]
- Rong, L.; Gilchrist, M.A.; Feng, Z.; Perelson, A.S. Modeling within-host HIV-1 dynamics and the evolution of drug resistance: Trade-offs between viral enzyme function and drug susceptibilitye. J. Theor. Biol. 2007, 247, 804–818. [Google Scholar] [CrossRef]
- Maziane, M.; Lotfi, E.M.; Hattaf, K.; Yousfi, N. Dynamics of a class of HIV infection models with cure of infected cells in eclipse stage. Acta Biotheor. 2015, 63, 363–380. [Google Scholar] [CrossRef] [PubMed]
- Lotfi, M.; Jabbari, A.; Rahbarghazi, R.; Kheiri, H. Innate immune response against COVID-19: The first report on the theory of COVID-19 treatment by a combined method of mathematics and medicine. Res. Sq. 2021, 1–21. [Google Scholar] [CrossRef]
- Sattentau, Q. Avoiding the void: Cell-to-cell spread of human viruses. Nat. Rev. Microbiol. 2008, 6, 815–826. [Google Scholar] [CrossRef]
- Marsh, M.; Helenius, A. Virus entry: Open sesame. Cell 2006, 124, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Mothes, W.; Sherer, N.M.; Jin, J.; Zhong, P. Virus cell-to-cell transmission. J. Virol. 2010, 84, 8360–8368. [Google Scholar] [CrossRef] [PubMed]
- Zeng, C.; Evans, J.P.; King, T.; Zheng, Y.M.; Oltz, E.M.; Whelan, S.P.J.; Saif, L.J.; Peeples, M.E.; Liu, S.L. SARS-CoV-2 spreads through cell-to-cell transmission. Proc. Natl. Acad. Sci. USA 2022, 119, e2111400119. [Google Scholar] [CrossRef]
- Wodarz, D.; Christensen, J.P.; Thomsen, A.R. The importance of lytic and nonlytic immune responses in viral infectionsn. Trends Immunol. 2002, 23, 194–200. [Google Scholar] [CrossRef]
- Wodarz, D.; Christensen, J.P.; Thomsen, A.R. Dynamical interactions between CTL and antibody responses. In Killer Cell Dynamics, Mathematical and Computational Approaches to Immunology; Wodarz, D., Ed.; Springer: New York, NY, USA, 2007; Volume 32, pp. 125–136. [Google Scholar]
- Buonomo, B.; Vargas-De-Leon, C. Global stability for an HIV-1 infection model including an eclipse stage of infected cells. J. Math. Anal. Appl. 2012, 385, 709–720. [Google Scholar] [CrossRef]
- Baccam, P.; Beauchemin, C.; Macken, C.A.; Hayden, F.G.; Perelson, A.S. Kinetics of Influenza A Virus Infection in Humans. J. Virol. 2006, 80, 7590–7599. [Google Scholar] [CrossRef]
- Rodriguez, T.; Dobrovolny, H.M. Estimation of viral kinetics model parameters in young and aged SARS-CoV-2 infected macaques. R. Soc. Open Sci. 2021, 8, 1–10. [Google Scholar] [CrossRef]
- Smith, H.L.; Waltman, P. The Theory of the Chemostat: Dynamics of Microbial Competition; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- LaSalle, J.P. The Stability of Dynamical Systems, Regional Conference Series in Applied Mathematics; SIAM: Philadelphia, PA, USA, 1976. [Google Scholar]
- Lee, H.Y.; Topham, D.J.; Park, S.Y.; Hollenbaugh, J.; Treanor, J.; Mosmann, T.R.; Jin, X.; Ward, B.M.; Miao, H.; Holden-Wiltse, J.; et al. Simulation and prediction of the adaptive immune response to influenza A virus infection. J. Virol. 2009, 83, 7151–7165. [Google Scholar] [CrossRef] [PubMed]
- Heitzman-Breen, N.; Ciupe, S.M. Modeling within-host and aerosol dynamics of SARS-CoV-2: The relationship with infectiousness. PLoS Comput. Biol. 2022, 8, e1009997. [Google Scholar] [CrossRef] [PubMed]
- Ke, R.; Zitzmann, C.; Ho, D.D.; Ribeiro, R.M.; Perelson, A.S. In vivo kinetics of SARS-CoV-2 infection and its relationship with a person’s infectiousness. Proc. Natl. Acad. Sci. USA 2021, 49, e2111477118. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, A.; Bertr, J.; Ke, R.; Comets, E.; De Lamballerie, X.; Malvy, D.; Pizzorno, A.; Terrier, O.; Rosa Calatrava, M.; Mentré, F.; et al. Timing of Antiviral Treatment Initiation is Critical to Reduce SARS-CoV-2 Viral Load. CPT Pharmacometrics Syst. Pharmacol. 2020, 9, 509–514. [Google Scholar] [CrossRef]
- Vaidya, N.K.; Bloomquist, A.; Perelson, A.S. Modeling Within-Host Dynamics of SARS-CoV-2 Infection: A Case Study in Ferrets. Viruses 2021, 8, 1635. [Google Scholar] [CrossRef]
- Wang, S.; Pan, Y.; Wang, Q.; Miao, H.; Brown, A.N.; Rong, L. Modeling the viral dynamics of SARS-CoV-2 infection. Math. Biosci. 2020, 328, 108438. [Google Scholar] [CrossRef]
- Tao, X.; Xu, A. Basic knowledge of immunology. In Amphioxus Immunity; Academic Press: Cambridge, MA, USA, 2016; pp. 15–42. [Google Scholar]
- Hattaf, K. On the stability and numerical scheme of fractional differential equations with application to biology. Computation 2022, 10, 97. [Google Scholar] [CrossRef]
- Hyder, A.; Barakat, M.A.; Rizk, D.; Shah, R.; Nonlaopon, K. Study of HIV model via recent improved fractional differential and integral operators. AIMS Math. 2022, 8, 1656–1671. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Definition | Value | Source |
|---|---|---|---|
| Epithelial cells | – | [9] | |
| production rate | cells mL day | ||
| Death rate of uninfected | day | [35] | |
| epithelial cells | |||
| Virus-to-cell infection rate | – | Estimated | |
| mL virion day | |||
| Cell-to-cell infection rate | 0–1 mL cell day | Assumed | |
| Death rate of latently | – day | Assumed | |
| infected epithelial cells | |||
| Rate to become productively | 1– day | Estimated | |
| infected cells | |||
| Death rate of productively | – day | Estimated | |
| infected epithelial cells | |||
| k | Virion production rate per | –580 | Estimated |
| infected epithelial cell | virions cell day | ||
| Virus clearance rate | –20 day | Estimated | |
| r | Activation rate of | 0–1 mL virion day | Assumed |
| antibodies | |||
| Death rate of antibodies | 0–1 day | Assumed | |
| p | Neutralization rate of | 0–1 | Assumed |
| virus by antibodies | mL molecules day | ||
| Non-lytic strength against | 0–1 mL molecules | Assumed | |
| virus-to-cell infection | |||
| Non-lytic strength against | 0–1 mL molecules | Assumed | |
| cell-to-cell infection | |||
| Cure rate of latently | 0–1 day | Assumed | |
| infected cells | |||
| Effectiveness of | 0–1 | Assumed | |
| antiviral treatment |
| Parameter | Value | Sensitivity Index |
|---|---|---|
| 500 | 1 | |
| 0.001 | −1 | |
| 0.0000011 | 0.88 | |
| 0.00000012 | 0.0115 | |
| 0.088 | −0.687 | |
| 4.5 | 0.844 | |
| 0.088 | −1 | |
| k | 88 | 0.88 |
| 10 | −0.885 | |
| 0.02 | −0.156 | |
| 0.2 | −0.2212 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hattaf, K.; El Karimi, M.I.; Mohsen, A.A.; Hajhouji, Z.; El Younoussi, M.; Yousfi, N. Mathematical Modeling and Analysis of the Dynamics of RNA Viruses in Presence of Immunity and Treatment: A Case Study of SARS-CoV-2. Vaccines 2023, 11, 201. https://doi.org/10.3390/vaccines11020201
Hattaf K, El Karimi MI, Mohsen AA, Hajhouji Z, El Younoussi M, Yousfi N. Mathematical Modeling and Analysis of the Dynamics of RNA Viruses in Presence of Immunity and Treatment: A Case Study of SARS-CoV-2. Vaccines. 2023; 11(2):201. https://doi.org/10.3390/vaccines11020201
Chicago/Turabian StyleHattaf, Khalid, Mly Ismail El Karimi, Ahmed A. Mohsen, Zakaria Hajhouji, Majda El Younoussi, and Noura Yousfi. 2023. "Mathematical Modeling and Analysis of the Dynamics of RNA Viruses in Presence of Immunity and Treatment: A Case Study of SARS-CoV-2" Vaccines 11, no. 2: 201. https://doi.org/10.3390/vaccines11020201
APA StyleHattaf, K., El Karimi, M. I., Mohsen, A. A., Hajhouji, Z., El Younoussi, M., & Yousfi, N. (2023). Mathematical Modeling and Analysis of the Dynamics of RNA Viruses in Presence of Immunity and Treatment: A Case Study of SARS-CoV-2. Vaccines, 11(2), 201. https://doi.org/10.3390/vaccines11020201