
Bacterial Outer Membrane Vesicles as a Platform for the Development of a Broadly Protective Human Papillomavirus Vaccine Based on the Minor Capsid Protein L2

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Cultures
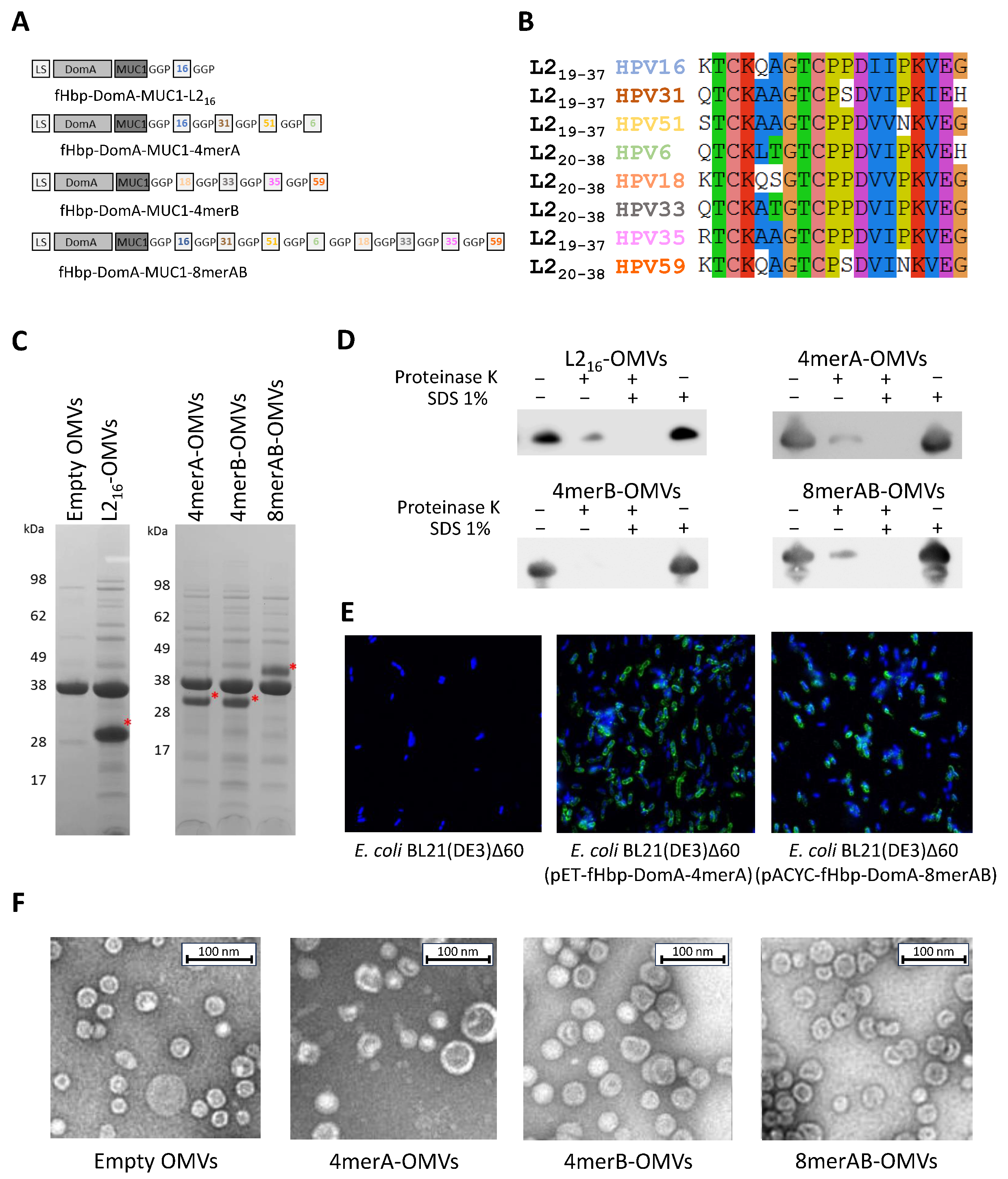
2.2. Cloning of the L2 Epitopes
2.3. OMV Preparation and Purification
2.4. Dynamic Light Scattering
2.5. Confocal Microscopy Analysis of Recombinant E. coli Strains
2.6. Analysis of Surface Localization of L2 Epitopes Using Proteinase K Assay
2.7. Negative Staining Electron Microscopy Analysis
2.8. Interleukin 6 (IL-6) Reactogenicity Assay
2.9. Animal Experiments
2.10. Enzyme-Linked Immunosorbent Assay (ELISA)
2.11. Eukaryotic Cell Cultures
2.12. Pseudovirions Preparation
2.13. Neutralization Assays
3. Results
3.1. L2 Epitope Is Efficiently Expressed on the Surface of E. coli and Accumulated in OMVs
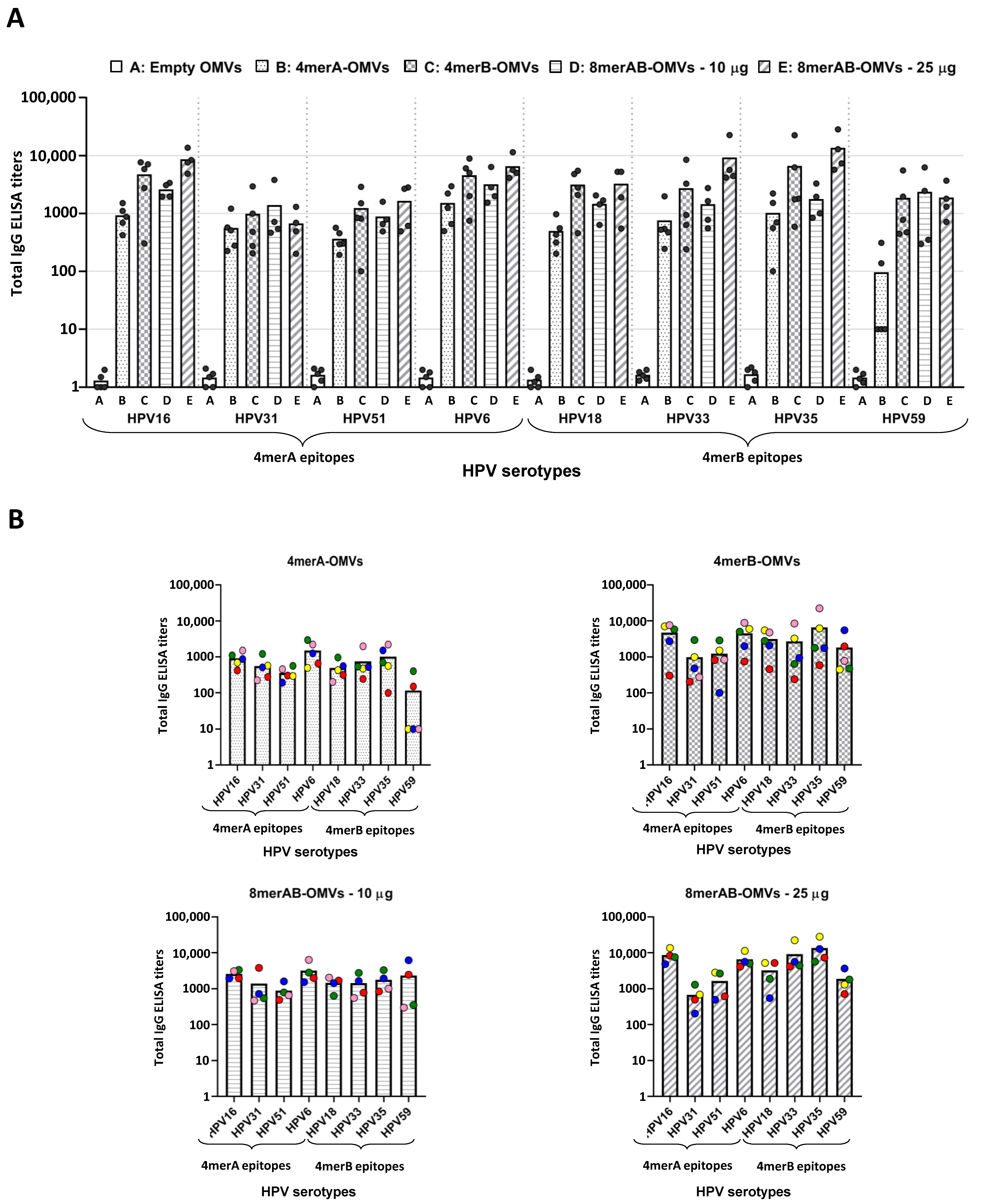
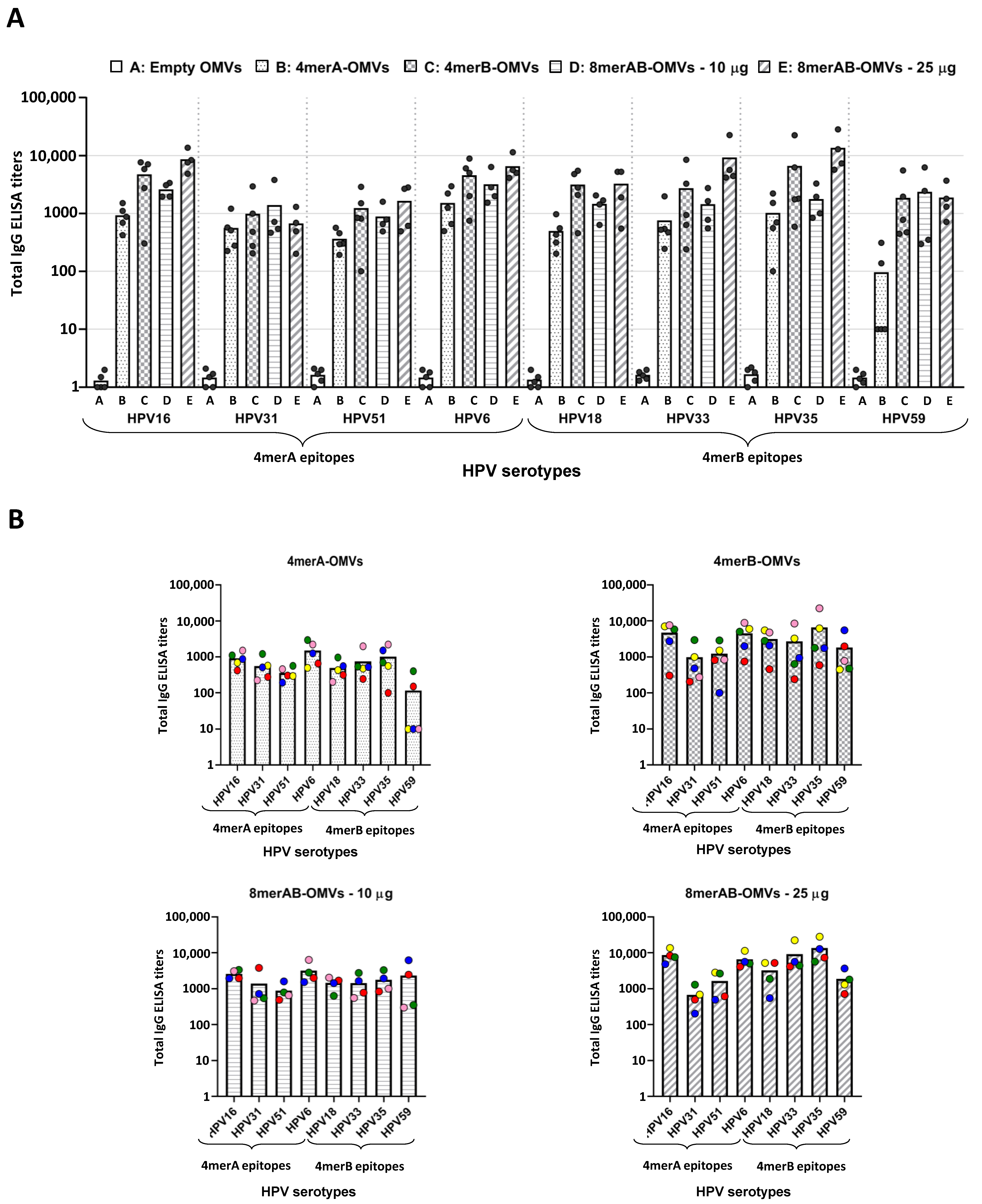
3.2. Immunization with L2-Engineered OMVs Elicits L2-Specific IgG Titers
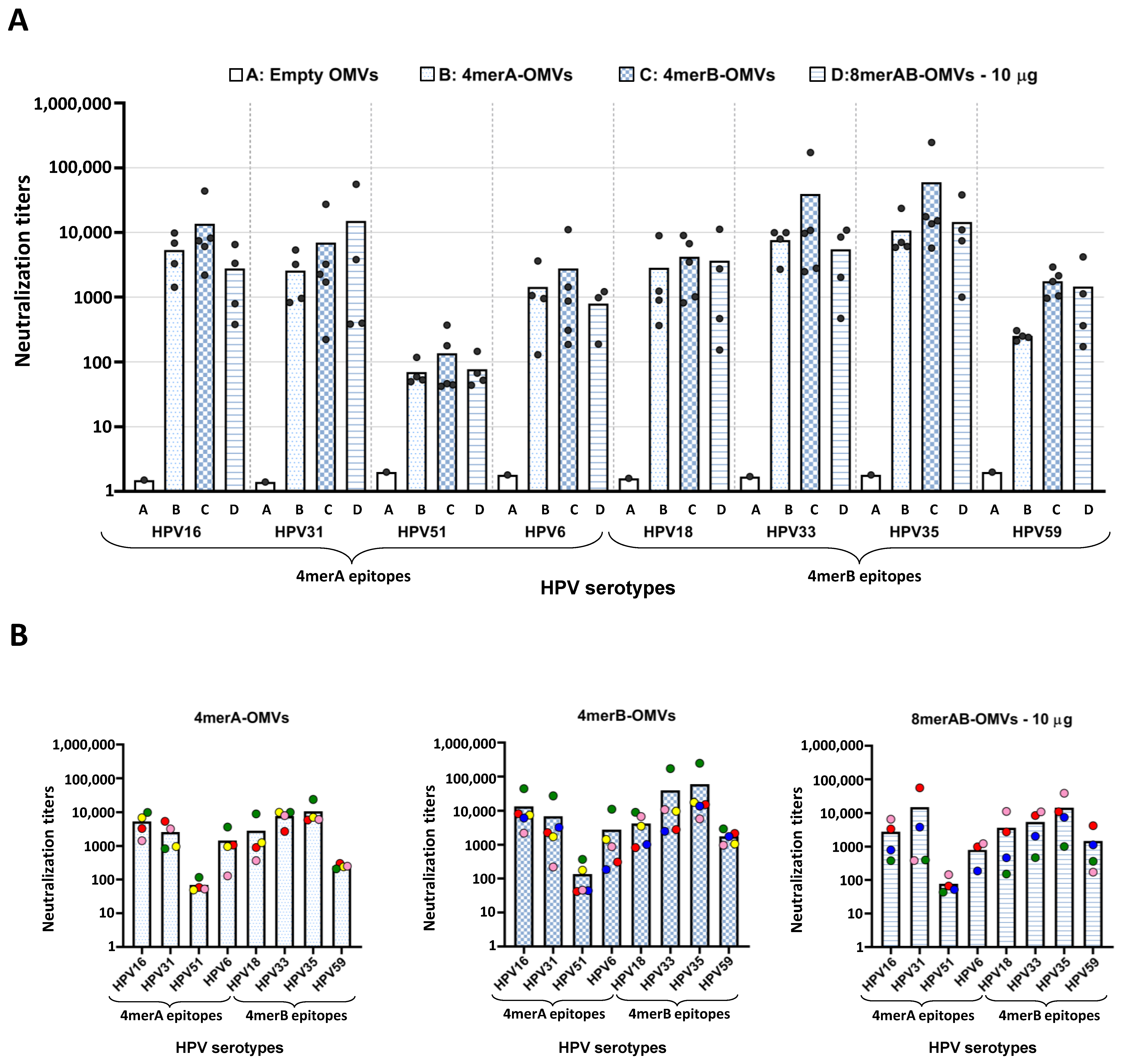
3.3. L2-Specific IgGs Elicited by OMV Immunization Neutralize HPV In Vitro
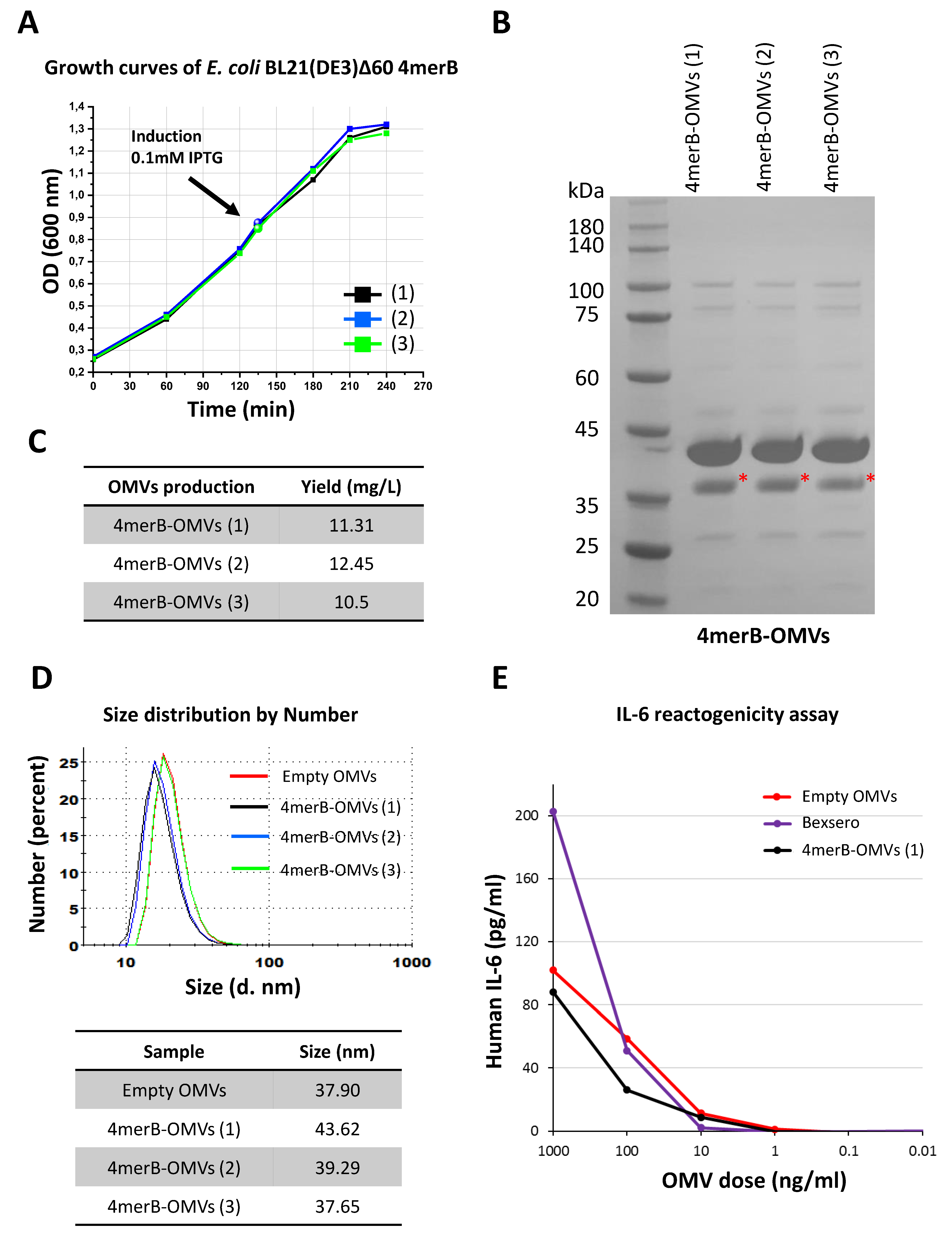
3.4. Set-Up of a Laboratory Scale Production Process of 4merB-OMVs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kulp, A.; Kuehn, M.J. Biological functions and biogenesis of secreted bacterial outer membrane vesicles. Annu. Rev. Microbiol. 2010, 64, 163–184. [Google Scholar] [CrossRef]
- Ellis, T.N.; Kuehn, M.J. Virulence and immunomodulatory roles of bacterial outer membrane vesicles. Microbiol. Mol. Biol. Rev. 2010, 74, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Osterrieder, N.; Metzger, S.M.; Buckles, E.; Doody, A.M.; DeLisa, M.P.; Putnam, D. Delivery of foreign antigens by engineered outer membrane vesicle vaccines. Proc. Natl. Acad. Sci. USA 2010, 107, 3099–3104. [Google Scholar] [CrossRef]
- Kesty, N.C.; Kuehn, M.J. Incorporation of heterologous outer membrane and periplasmic proteins into Escherichia coli Outer Membrane Vesicles. J. Biol. Chem. 2004, 279, 2069–2076. [Google Scholar] [CrossRef] [PubMed]
- Gerritzen MJ, H.; Martens, D.E.; Wijffels, R.H.; van der Pol, L.; Stork, M. Bioengineering bacterial outer membrane vesicles as vaccine platform. Biotechnol. Adv. 2017, 35, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Irene, C.; Fantappiè, L.; Caproni, E.; Zerbini, F.; Anesi, A.; Tomasi, M.; Zanella, I.; Stupia, S.; Prete, S.; Valensin, S.; et al. Bacterial outer membrane vesicles engineered with lipidated antigens as a platform for Staphylococcus aureus vaccine. Proc. Natl. Acad. Sci. USA 2019, 116, 21780–21788. [Google Scholar] [CrossRef]
- Berlanda Scorza, F.; Colucci, A.M.; Maggiore, L.; Sanzone, S.; Rossi, O.; Ferlenghi, I.; Pesce, I.; Caboni, M.; Norais, N.; Di Cioccio, V.; et al. High yield production process for Shigella outer membrane particles. PLoS ONE 2012, 7, e35616. [Google Scholar] [CrossRef]
- Ladhani, S.N.; Ramsay, M.; Borrow, R.; Riordan, A.; Watson, J.M.; Pollard, A.J. Enter B and W: Two new meningococcal vaccine programmes launched. Arch. Dis. Child. 2016, 101, 91–95. [Google Scholar] [CrossRef]
- Serruto, D.; Bottomley, M.J.; Ram, S.; Giuliani, M.M.; Rappuoli, R. The new multicomponent vaccine against meningococcal serogroup B, 4CMenB: Immunological, functional and structural characterization of the antigens. Vaccine 2012, 30, B87–B97. [Google Scholar] [CrossRef]
- Gerke, C.; Colucci, A.M.; Giannelli, C.; Sanzone, S.; Vitali, C.G.; Sollai, L.; Rossi, O.; Martin, L.B.; Auerbach, J.; Di Cioccio, V.; et al. Production of a Shigella sonnei vaccine based on generalized modules for membrane antigens (GMMA), 1790GAHB. PLoS ONE 2015, 10, e0134478. [Google Scholar] [CrossRef]
- Rossi, O.; Caboni, M.; Negrea, A.; Necchi, F.; Alfini, R.; Micoli, F.; Saul, A.; MacLennan, C.A.; Rondini, S.; Gerke, C. Toll-Like receptor activation by generalized modules for membrane antigens from Lipid A mutants of Salmonella enterica Serovars Typhimurium and Enteritidis. Clin. Vaccine Immunol. 2016, 23, 304–314. [Google Scholar] [CrossRef]
- Zanella, I.; König, E.; Tomasi, M.; Gagliardi, A.; Frattini, L.; Fantappiè, L.; Irene, C.; Zerbini, F.; Caproni, E.; Isaac, S.J.; et al. Proteome-minimized outer membrane vesicles from Escherichia coli as a generalized vaccine platform. J. Extracell. Vesicles 2021, 10, e12066. [Google Scholar] [CrossRef] [PubMed]
- Fantappie, L.; Irene, C.; De Santis, M.; Armini, A.; Gagliardi, A.; Tomasi, M.; Parri, M.; Cafardi, V.; Bonomi, S.; Ganfini, L.; et al. Some Gram-negative lipoproteins keep their surface topology when transplanted from one species to another and deliver foreign polypeptides to the bacterial surface. Mol. Cell. Proteom. 2017, 16, 1348–1364. [Google Scholar] [CrossRef] [PubMed]
- König, E.; Gagliardi, A.; Riedmiller, I.; Andretta, C.; Tomasi, M.; Irene, C.; Frattini, L.; Zanella, I.; Berti, F.; Grandi, A.; et al. Multi-Antigen Outer Membrane Vesicle Engineering to Develop Polyvalent Vaccines: The Staphylococcus aureus Case. Front. Immunol. 2021, 12, 752168. [Google Scholar] [CrossRef] [PubMed]
- Grandi, A.; Tomasi, M.; Accordini, S.; Bertelli, C.; Vanzo, T.; Gagliardi, A.; Caproni, E.; Tamburini, S.; Fantappie, L.; Di Lascio, G.; et al. Immunogenicity and pre-clinical efficacy of an OMV-based SARS-CoV-2 vaccine. Vaccines 2023, 11, 1546. [Google Scholar] [CrossRef]
- McBride, A.A. Human papillomaviruses: Diversity, infection and host interactions. Nat. Rev. Microbiol. 2021, 20, 95–108. [Google Scholar] [CrossRef]
- zur Hausen, H. Papillomaviruses Causing Cancer: Evasion From Host-Cell Control in Early Events in Carcinogenesis. J. Natl. Cancer Inst. 2020, 92, 690–698. [Google Scholar] [CrossRef]
- Bernard, H.U.; Burk, R.D.; Chen, Z.; Van Doorslaer, K.; zur Hausen, H.; de Villiers, E.-M. Classification of papillomaviruses (PVs) based on 189 PV types and proposal of taxonomic amendments. Virology 2010, 401, 70–79. [Google Scholar] [CrossRef]
- Munoz, N.; Bosch, F.X.; Castellsagué, X.; Díaz, M.; de Sanjose, S.; Hammouda, D.; Shah, K.V.; Meijer, C.J. Against which human papillomavirus types shall we vaccinate and screen? The international perspective. Int. J. Cancer 2004, 111, 278–285. [Google Scholar] [CrossRef]
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. Human papillomaviruses. IARC Monogr. Eval. Carcinog. Risks Hum. 2007, 90, 1–636. [Google Scholar]
- zur Hausen, H. Papillomaviruses in the causation of human cancers—A brief historical account. Virology 2009, 384, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Winer, R.L.; Lee, S.K.; Hughes, J.P.; Adam, D.E.; Kiviat, N.B.; Koutsky, L.A. Genital human papillomavirus infection: Incidence and risk factors in a cohort of female university students. Am. J. Epidemiol. 2003, 157, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Dunne, E.F.; Sternberg, M.; Markowitz, L.E.; McQuillan, G.; Swan, D.; Patel, S.; Unger, E.R. Human papillomavirus (HPV) 6, 11, 16, and 18 prevalence among females in the United States—National Health And Nutrition Examination Survey, 2003–2006: Opportunity to measure HPV vaccine impact? J. Infect. Dis. 2001, 204, 562–565. [Google Scholar] [CrossRef]
- Rodriguez, A.C.; Schiffman, M.; Herrero, R.; Wacholder, S.; Hildesheim, A.; Castle, P.E.; Solomon, D.; Burk, R.; On behalf of the Proyecto Epidemiologico Guanacaste Group. Rapid clearance of human papillomavirus and implications for clinical focus on persistent infections. J. Natl. Cancer Inst. 2008, 100, 513–517. [Google Scholar] [CrossRef]
- Hildesheim, A.; Hadjimichael, O.; Schwartz, P.E.; Wheeler, C.M.; Barnes, W.; Lowell, D.M.; Willett, J.; Schiffman, M. Risk factors for rapid-onset cervical cancer. Am. J. Obstet. Gynecol. 1999, 180, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Kirnbauer, R.; Booy, F.; Cheng, N.; Lowy, D.R.; Schiller, J. Papillomavirus L1 major capsid protein self-assembles into virus-like particles that are highly immunogenic. Proc. Natl. Acad. Sci. USA 1992, 89, 12180–12184. [Google Scholar] [CrossRef]
- Day, P.M.; Gambhira, R.; Roden, R.B.; Lowy, D.R.; Schiller, J.T. Mechanisms of human papillomavirus type 16 neutralization by L2 cross-neutralizing and L1 type-specific antibodies. J. Virol. 2008, 82, 4638–4646. [Google Scholar] [CrossRef]
- Rubio, I.; Seitz, H.; Canali, E.; Sehr, P.; Bolchi, A.; Tommasino, M.; Ottonello, S.; Müller, M. The N-terminal region of the human papillomavirus L2 protein contains overlapping binding sites for neutralizing, cross-neutralizing and non-neutralizing antibodies. Virology 2011, 409, 348–359. [Google Scholar] [CrossRef]
- Seitz, H.; Ribeiro-Müller, L.; Canali, E.; Bolchi, A.; Tommasino, M.; Ottonello, S.; Müller, M. Robust In vitro and in vivo neutralization against multiple high-risk HPV types induced by a thermostable thioredoxin-L2 vaccine. Cancer Prev. Res. 2015, 8, 932–941. [Google Scholar] [CrossRef]
- Seitz, H.; Dantheny, T.; Burkart, F.; Ottonello, S.; Muller, M. Influence of oxidation and multimerization on the immunogenicity of a thioredoxin-L2 prophylactic papillomavirus vaccine. Clin. Vaccine Immunol. 2013, 20, 1061–1069. [Google Scholar] [CrossRef]
- Seitz, H.; Canali, E.; Ribeiro-Müller, L.; Pàlfi, A.; Bolchi, A.; Tommasino, M.; Ottonello, S.; Müller, M. A three component mix of thioredoxin-L2 antigens elicits broadly neutralizing responses against oncogenic human papillomaviruses. Vaccine 2014, 32, 2610–2617. [Google Scholar] [CrossRef]
- Rubio, I.; Bolchi, A.; Moretto, N.; Canali, E.; Gissmann, L.; Tommasino, M.; Müller, M.; Ottonello, S. Potent anti-HPV immune responses induced by tandem repeats of the HPV16 L2 (20–38) peptide displayed on bacterial thioredoxin. Vaccine 2009, 27, 1949–1956. [Google Scholar] [CrossRef]
- Canali, E.; Bolchi, A.; Spagnoli, G.; Seitz, H.; Rubio, I.; Pertinhez, T.A.; Müller, M.; Ottonello, S. A high-performance thioredoxin-based scaffold for peptide immunogen construction: Proof-of-concept testing with a human papillomavirus epitope. Sci. Rep. 2014, 4, 4729. [Google Scholar] [CrossRef]
- Pouyanfard, S.; Spagnoli, G.; Bulli, L.; Balz, K.; Yang, F.; Odenwald, C.; Seitz, H.; Mariz, F.C.; Bolchi, A.; Ottonello, S.; et al. Minor Capsid Protein L2 Polytope Induces Broad Protection against Oncogenic and Mucosal Human Papillomaviruses. J. Virol. 2018, 92, e01930-17. [Google Scholar] [CrossRef]
- Klock, H.E.; Lesley, S.A. The Polymerase Incomplete Primer Extension (PIPE) method applied to high-throughput cloning and site-directed mutagenesis. Methods Mol. Biol. 2009, 498, 91–103. [Google Scholar] [PubMed]
- Bartolomé, B.; Jubete, Y.; Martínez, E.; de la Cruz, F. Construction and properties of a family of pACYC184-derived cloning vectors compatible with pBR322 and its derivatives. Gene 1991, 102, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Christopher, B.; Buck, C.D.T. Production of Papillomavirus-Based Gene Transfer Vectors. Curr. Protoc. Cell Biol. 2007, 37, 26.1.1–26.1.19. [Google Scholar]
- Konar, M.; Rossi, R.; Walter, H.; Pajon, R.; Beernink, P.T. A mutant library approach to identify improved meningococcal factor H binding protein vaccine antigens. PLoS ONE 2015, 10, e0128185. [Google Scholar] [CrossRef]
- Hooda, Y.; Lai, C.C.-L.; Judd, A.; Buckwalter, C.M.; Shin, H.E.; Gray-Owen, S.D.; Moraes, T.F. Slam is an outer membrane protein that is required for the surface display of lipidated virulence factors in Neisseria. Nat. Microbiol. 2016, 1, 160. [Google Scholar] [CrossRef]
- Valentini, S.; Santoro, G.; Baffetta, F.; Franceschi, S.; Paludi, M.; Brandini, E.; Gherardini, L.; Serruto, D.; Capecchi, B. Monocyte-activation test to reliably measure the pyrogenic content of a vaccine: An in vitro pyrogen test to overcome in vivo limitations. Vaccine 2019, 37, 3754–3760. [Google Scholar] [CrossRef]
- Spagnoli, G.; Pouyanfard, S.; Cavazzini, D.; Canali, E.; Maggi, S.; Tommasino, M.; Bolchi, A.; Müller, M.; Ottonello, S. Broadly neutralizing antiviral responses induced by a single molecule HPV vaccine based on thermostable thioredoxin-L2 multiepitope nanoparticles. Sci. Rep. 2017, 7, 1800. [Google Scholar] [CrossRef] [PubMed]
- Romanowski, B.; de Borba, P.C.; Naud, P.; Roteli-Martins, C.M.; De Carvalho, N.S.; Teixeira, J.C.; Aoki, F.; Ramjattan, B.; Shier, R.M.; Somani, R.; et al. Sustained efficacy and immunogenicity of the human papillomavirus (HPV)-16/18 AS04-adjuvanted vaccine: Analysis of a randomised placebo-controlled trial up to 6.4 years. Lancet 2009, 374, 1975–1985. [Google Scholar] [PubMed]
- Beatty, P.L.; Finn, O.J. Preventing cancer by targeting abnormally expressed self-antigens: MUC1 vaccines for prevention of epithelial adenocarcinomas. Ann. N. Y. Acad. Sci. 2013, 1284, 52–56. [Google Scholar] [CrossRef]
- Kimura, T.; Finn, O.J. MUC1 immunotherapy is here to stay. Expert Opin. Biol. Ther. 2013, 13, 35–49. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; McKolanis, J.R.; Dzubinski, L.A.; Islam, K.; Potter, D.M.; Salazar, A.M.; Schoen, R.E.; Finn, O.J. MUC1 Vaccine for Individuals with Advanced Adenoma of the Colon: A Cancer Immunoprevention Feasibility Study. Cancer Prev. Res. 2013, 6, 18–26. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tamburini, S.; Zhang, Y.; Gagliardi, A.; Di Lascio, G.; Caproni, E.; Benedet, M.; Tomasi, M.; Corbellari, R.; Zanella, I.; Croia, L.; et al. Bacterial Outer Membrane Vesicles as a Platform for the Development of a Broadly Protective Human Papillomavirus Vaccine Based on the Minor Capsid Protein L2. Vaccines 2023, 11, 1582. https://doi.org/10.3390/vaccines11101582
Tamburini S, Zhang Y, Gagliardi A, Di Lascio G, Caproni E, Benedet M, Tomasi M, Corbellari R, Zanella I, Croia L, et al. Bacterial Outer Membrane Vesicles as a Platform for the Development of a Broadly Protective Human Papillomavirus Vaccine Based on the Minor Capsid Protein L2. Vaccines. 2023; 11(10):1582. https://doi.org/10.3390/vaccines11101582
Chicago/Turabian StyleTamburini, Silvia, Yueru Zhang, Assunta Gagliardi, Gabriele Di Lascio, Elena Caproni, Mattia Benedet, Michele Tomasi, Riccardo Corbellari, Ilaria Zanella, Lorenzo Croia, and et al. 2023. "Bacterial Outer Membrane Vesicles as a Platform for the Development of a Broadly Protective Human Papillomavirus Vaccine Based on the Minor Capsid Protein L2" Vaccines 11, no. 10: 1582. https://doi.org/10.3390/vaccines11101582
APA StyleTamburini, S., Zhang, Y., Gagliardi, A., Di Lascio, G., Caproni, E., Benedet, M., Tomasi, M., Corbellari, R., Zanella, I., Croia, L., Grandi, G., Müller, M., & Grandi, A. (2023). Bacterial Outer Membrane Vesicles as a Platform for the Development of a Broadly Protective Human Papillomavirus Vaccine Based on the Minor Capsid Protein L2. Vaccines, 11(10), 1582. https://doi.org/10.3390/vaccines11101582