1. Introduction
The dramatic SARS-CoV-2 pandemic exploded worldwide at the beginning of 2020 and has triggered an unprecedented race to the development of effective vaccines. In less than a three-year timeframe, hundreds of vaccines have been designed and tested in preclinical settings, more than 100 have reached the clinic, and 24 are currently authorized for human use [
1]. It is estimated that more than 9 billion doses have been administered so far worldwide, saving approximately 1 million lives.
Despite this spectacular success of modern vaccinology, the race against COVID-19 is not over. Mostly because of costs and logistic issues, vaccine distribution is heavily unbalanced, with half of the planet still waiting for a dose and with only 4% of populations in low-income countries being vaccinated [
2]. Moreover, SARS-CoV-2 has the extraordinary capacity to continuously accumulate mutations, which allow the virus to escape, at least partially, host immune responses while preserving infectivity and virulence [
3].
To overcome such challenges and to provide a sustainable long-term prophylaxis, a “pan-vaccine” capable of eliciting a broad, cross-protective immune response should become available. The ideal vaccine would negate the need of booster immunizations using vaccines tailored for the emerging variants of concern (VOCs). In addition, the vaccine should rely on easily scalable, low-cost production processes, while not requiring the cold chain to simplify world-wide distribution.
Among the several technologies available for vaccine development, outer membrane vesicles (OMVs) have emerged in recent years as an attractive tool capable of coupling excellent built-in adjuvanticity provided by the microbe-associated-molecular patterns (MAMPs) embedded in the vesicles, and an easily scalable production and purification process [
4]. Anti-Neisseria OMV-based vaccines are currently available for human use [
5], and others against Shigella and Salmonella are in advanced clinical phases [
6,
7].
We have recently developed a platform based on “proteome minimized”
E. coli OMVs selectively loaded with heterologous antigens [
8]. The platform has been successfully applied to design prophylactic vaccines against infectious diseases [
9] and was shown to stimulate potent anti-tumour activity in different mouse models [
10,
11].
Because OMVs are readily phagocytosed, the associated antigens are efficiently presented by professional antigen-presenting cells, eliciting both antibody- and T-cell responses [
12]. The reaction is coupled to the production of IFN-γ, ensuring a sustained Th1 response as well as an optimal humoral response. Clinical evidence demonstrates that an accelerated induction of a Th1 cell response is associated with less severe cases of COVID-19 [
13,
14]. Moreover, convalescent individuals tend to develop strong memory CD4+ and CD8+ T cells [
15]. Therefore, the ability of OMVs to trigger Th1 represents a desired feature. Crucially, in addition to the simplicity and cost-effectiveness of OMV production pipelines [
16], the antigen-decorated vesicles are extremely stable for long-term storage at room temperature, potentially making them ideal vaccine candidates for world-wide distribution.
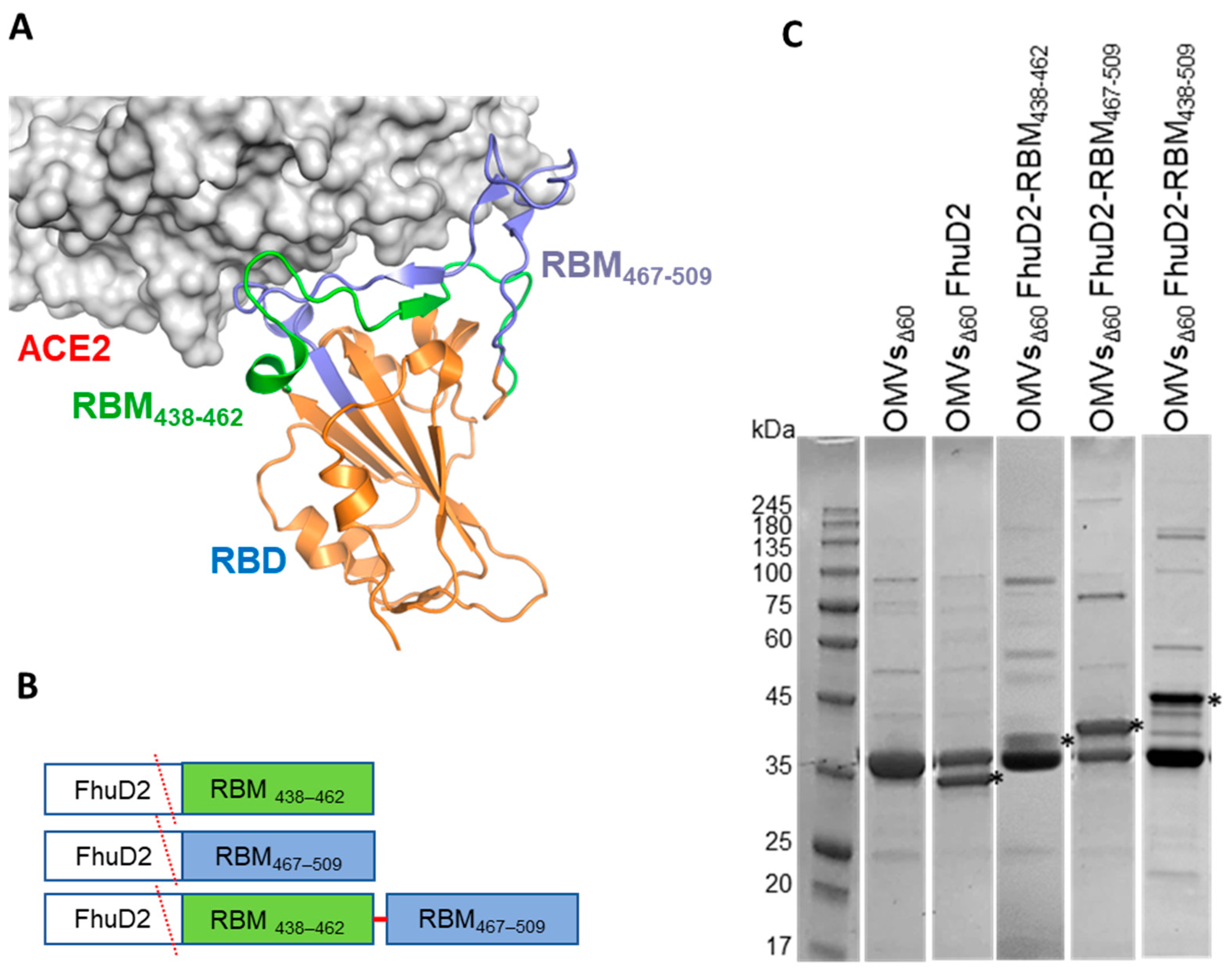
Essentially, all available vaccines and those under development are designed to induce antibodies specific for the spike (S) protein or its receptor binding domain (RBD). Neutralizing titres found in vaccinees or in convalescent individuals correlate strongly with antibody binding to the RBD [
17]. Moreover, the most potent monoclonal antibodies isolated from convalescent patients recognize epitopes located within the receptor binding motif (RBM), the RBD region which directly engages the viral receptor, angiotensin-converting enzyme 2 (ACE2) [
18]. The ability to specifically concentrate the immune response against these epitopes would exclusively elicit neutralizing antibodies, while minimizing the generation of non-neutralizing or poorly neutralizing immunoglobulins binding to irrelevant spike regions.
The OMVs offer the unique opportunity to display short and defined epitopes to B- and T-cells in a highly immunogenic context provided by the bacterial vesicle. Taking advantage of such potential and the availability of the crystal structure of the RBD in complex with ACE2, we engineered the OMVs with peptides from the SARS-CoV-2 ancestral RBM. Here, we show that RBM-derived peptides can be expressed in the OMV membrane and induce neutralizing antibodies sufficient to fully protect human ACE2 (hACE2)-transgenic mice from a challenge with SARS-CoV-2. We also show that the same region from the BA.1 variant can also be efficiently expressed in the OMVs. Moreover, the RBM438–509 ancestral-OMVs elicit antibodies which can neutralize in vitro both the homologous ancestral strain and the Omicron BA.1 and the BA.5 variants, indicating that OMV-RBM formulations elicit novel populations of antibodies with robust cross-reactive neutralization capacity.
Given the efficacy of the vaccine in the animal model, the cross-neutralization capacity, the ease of its engineering and the cost-effective production process, we propose the RBM438–509 ancestral-OMVs as a promising candidate to continue the vaccination campaign against SARS-CoV-2.
2. Materials and Methods
2.1. Engineering BL21(DE3) E. coli Strains with SARS-CoV-2 Neutralizing Epitopes
The pET21-FhuD2 plasmid carrying the
Staphylococcus aureus Ferric hydroxamate receptor 2 (FhuD2) was fused to one copy of RBM
438–462, RBM
467–509 and RBM
438–509 SARS-CoV-2 epitope, respectively. The three plasmids were assembled using the PIPE method [
19]. Briefly, pET21-FhuD2 was linearized by PCR, using FhuD2-v-R and pET-V-F primers (
Table 1). In parallel, the synthetic DNA encoding the RBM of SARS-CoV-2 was synthesized by GeneArt (Thermo Fisher Scientific, Waltham, MA, USA) and used as a template for the amplification of the three epitopes. In detail, RBM
438–462 and RBM
467–509 (
Table 2) were amplified by PCR with the forward 2-F and 1-F and the reverse 2-R and 1-R primers, respectively. The PCR products and the linearized plasmid were mixed together and used to transform the
E. coli HK100 strain.
The RBM438–509, the combination of the two epitopes, was assembled with two steps of PCR in succession. In the first step, two different fragments carrying an overlapping sequence were amplified with 2-F/R-1 and F-1/1-R primers. In the second step, the two fragments were eluted from Agarose gel, and mixed and used as template for a final PCR reaction with the primers 2-F and 1-R. This final product and the linearized plasmid were mixed and used to transform the E. coli HK100 strain.
The RBD-BA.1438–509 gene was synthesized by GeneArt (Thermo Fisher Scientific, Waltham, MA, USA) and used as template for the amplification of the RBM-BA.1438–509 using the CoSASta-F and CoSASta-R primers. The low copy number pACYC plasmid was linearized using the couple of primers FhuD2-v-R and PACYC-F The PCR product and the linearized plasmid were mixed and used to transform E. coli HK100 strain.
To confirm the correct gene fusions, plasmids were sequenced (Eurofins, Ebersberg, Germany) and the E. coli BL21(DE3)Δ60 strain was transformed with pET21-FhuD2-RBM438–462, pET21-FhuD2-RBM467–509, pET21-FhuD2-RBM438–509 and pACYC-FhuD2-RBM-BA.1438–509 plasmids and the derived recombinant strains were used for the production of engineered FhuD2-RBM438–462, FhuD2-RBM467–509, FhuD2-RBM438–509 (RBMancestral-OMVs) and FhuD2-RBM-BA.1438–509 (RBMBA.1-OMVs) OMVs, respectively.
2.2. OMV Purification
OMVs from K-12
E. coli BL21(DE3)
Δ60-pET-FhuD2-RBM
438–509, BL21(DE3)
Δ60-pET-FhuD2-RBM
438–462, BL21(DE3)
Δ60-pET-FhuD2-RBM
467–509 and BL21(DE3)
Δ60-pACYC-FhuD2-RBMBA.1
438–509 were purified in an EZ control bioreactor (Applikon Biotechnology, Schiedam, The Netherlands) as previously described [
8]. Cultures were started at an OD
600 of 0.1 and grown in LB medium at 30 °C, pH 6.8 (±0.2), dO
2 > 30%, 280–500 rpm until OD
600 = 0.5, then the temperature was lowered to 25 °C. The expression of the recombinant protein was induced when the culture reached an OD
600 of 1 with 0.1 mM IPTG and a feed made of 50 mg/L ampicillin, 15 g/L glycerol, 0.25 g/L MgSO
4 was added to the culture medium. The fermentation was carried out until the end of the exponential phase at 25 °C. OMVs were then purified and quantified as previously described [
8]. Culture supernatants were separated from biomass by centrifugation at 4000×
g for 20 min. After filtration through a 0.22-μm pore size filter (Millipore, Burlington, MA, USA), OMVs were isolated, concentrated and diafiltrated from the supernatants using Tangential Flow Filtration (TFF) with a Cytiva Äkta Flux system. OMVs were quantified using a DC protein assay (Bio-Rad, Hercules, CA, USA) and their protein content analysed by loading 20 μg of total protein on SDS-PAGE.
2.3. Studies with CD1 Mice
Mice were monitored twice per day to evaluate early signs of pain and distress, such as respiration rate, posture, and loss of weight (more than 20%) according to humane endpoints. Animals showing such conditions were anesthetized and subsequently sacrificed in accordance with experimental protocols, which were reviewed and approved by the Animal Ethical Committee of The University of Trento and the Italian Ministry of Health.
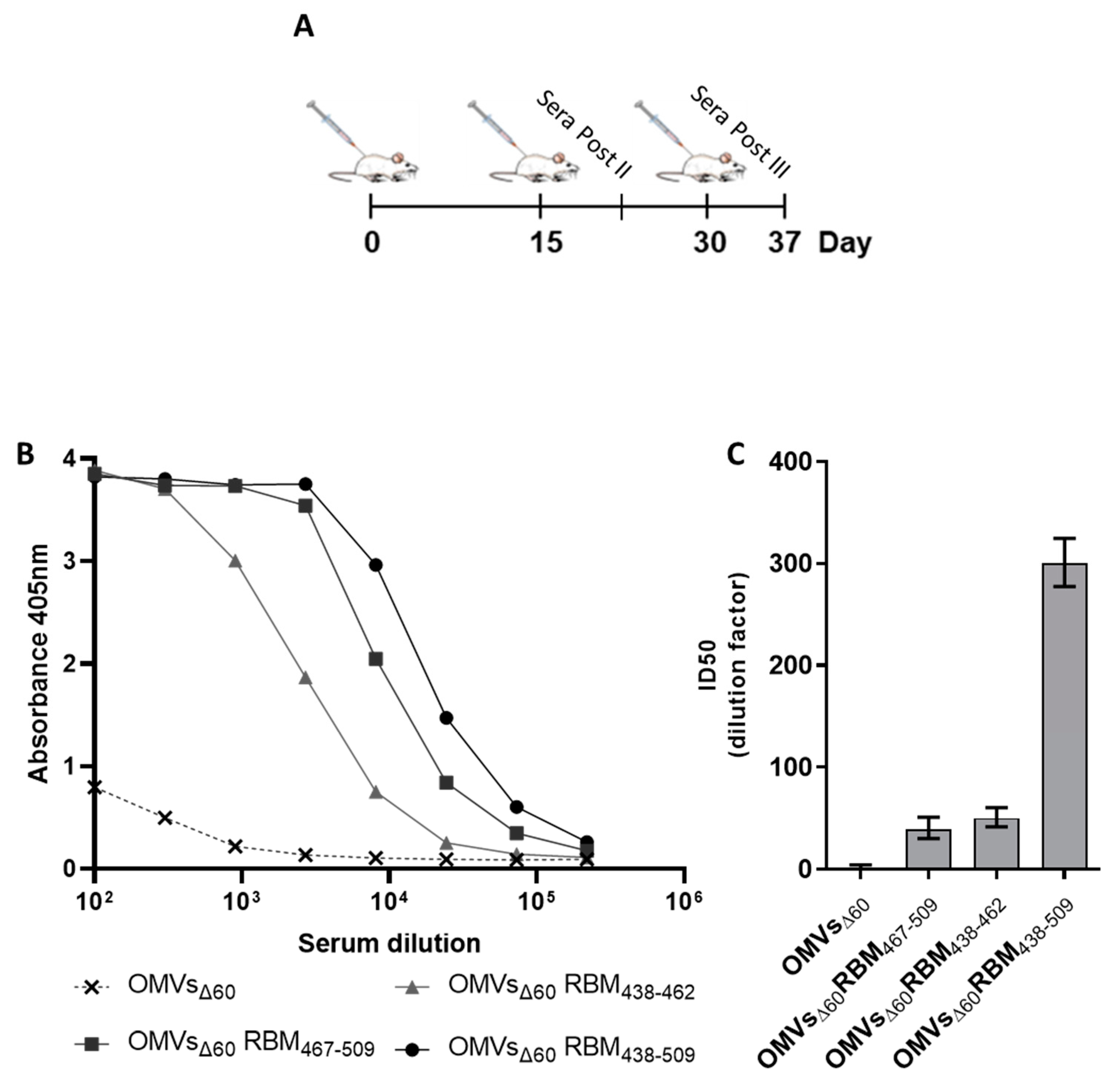
Five-week-old CD1 female mice were immunized intraperitoneally (i.p.) on day 0, 14 and 28 with 10 μg of OMVs together with 2 mg/mL Aluminium hydroxide in a final volume of 200 µL. Sera were collected 7 days after the last immunization. Alternatively, mice received intramuscularly (i.m.) 50 µL of AstraZeneca ChAdOx1 nCoV-19 vaccine (AZ), consisting of roughly 2.5 × 107 infectious units.
2.4. Studies with the ACE2 Mouse Model
B6.Cg-Tg(K18-hACE2)
2Prlmn/J mice [
1] were purchased from The Jackson Laboratory. Mice were housed under specific pathogen-free conditions and heterozygous mice were used at 8–10 weeks of age. All experimental animal procedures were approved by the Institutional Animal Committee of the San Raffaele Scientific Institute and all infectious work was performed in designed BSL-3 workspaces. The hCoV-19/Italy/LOM-UniSR-1/2020 (EPI_ISL_413489) isolate of SARS-CoV-2 was obtained from the Unit of Microbiology and Virology of San Raffaele Scientific Institute and grown in Vero E6 cells. K18-hACE2-transgenic mice were i.p. immunized twice, 14 days apart, with 10 µg vaccine in 200 μL of PBS or with PBS alone. Virus infection was performed via intranasal administration of 1 × 10
5 TCID
50 SARS-CoV-2 per mouse. Tissue homogenates were prepared by homogenizing perfused lung using gentleMACS Octo Dissociator (Miltenyi Biotec, #130-096-427, Bergisch Gladbach, Germany) in M tubes (#130-093-335) containing 1 mL of DMEM. Samples were homogenized three times with the program m_Lung_01_02 (34 s, 164 rpm). The homogenates were centrifuged at 3.500 rpm for 5 min at 4 °C. The supernatant was collected and stored at −80 °C for viral isolation and viral load detection. Viral titres were calculated by 50% tissue culture infectious dose (TCID
50). Briefly, Vero E6 cells were seeded at a density of 1.5 × 10
4 cells per well in flat-bottom 96-well tissue-culture plates. The following day, 2-fold dilutions of the homogenized tissue were applied to confluent cells and incubated for 1 h at 37 °C. Then, cells were washed with phosphate-buffered saline (PBS) and incubated for 72 h at 37 °C in DMEM 2% FBS. Cells were fixed with 4% paraformaldehyde for 20 min and stained with 0.05% (
w/
v) crystal violet in 20% methanol.
2.5. RNA Extraction and qPCR
Tissue homogenates were prepared by homogenizing perfused lungs using gentleMACS dissociator (Miltenyi Biotec, #130-096-427, Bergisch Gladbach, Germany ) with program RNA_02 in M tubes (#130-096-335) in 1 mL of Trizol (Invitrogen, #15596018, Waltham, MA, USA). The homogenates were centrifuged at 2000× g for 1 min at 4 °C and the supernatant was collected. RNA extraction was performed by combining phenol/guanidine-based lyisis with silica membrane-based purification. Briefly, 100 µL of Chloroform was added in 500 mL of homogenized sample and the total RNA was extracted using a ReliaPrep™ RNA Tissue Miniprep column (Promega, Cat #Z6111, Madison, WI, USA). Total RNA was isolated according to the manufacturer’s instructions. qPCR was performed using TaqMan Fast virus 1 Step PCR Master Mix (Life Technologies #4444434, Carlsbad, CA, USA), the standard curve was drawn with 2019_nCOV_N Positive control (IDT#10006625), and the primers used are 2019-nCoV_N1-Forward Primer (5′-GAC CCC AAA ATC AGC GAA AT-3′), 2019-nCoV_N1-Reverse Primer (5′-TCT GGT TAC TGC CAG TTG AAT CTG-3′) 2019-nCoV_N1-Probe (5′-FAM-ACC CCG CAT TAC GTT TGG TGG ACC-BHQ1-3′) (Centers for Disease Control and Prevention (CDC), Atlanta, GA, USA). All experiments were performed in duplicate.
2.6. Cell Isolation and Flow Cytometry
Mice were euthanized by cervical dislocation. Lungs were perfused through the right ventricle with PBS at the time of autopsy. Lung tissue was digested in RPMI 1640 containing 3.2 mg/mL Collagenase IV (Sigma, #C5138, St. Louis, MO, USA) and 25 U/mL DNAse I (Sigma-Aldrich, #D4263, St. Louis, MO, USA) for 30 min at 37 °C. Homogenized lungs were passed through a 70 μm nylon mesh to obtain a single-cell suspension. Cells were resuspended in 36% percoll solution (Sigma #P4937, St. Louis, MO, USA) and centrifuged for 20 min at 2000 rpm (light acceleration and low brake). The remaining red blood cells were removed with ACK lysis. Cell viability was assessed by staining with Viobility™ 405/520 fixable dye (Miltenyi Biotec, Cat #130-109-814, Bergisch Gladbach, Germany). The antibodies (Abs) used are presented in
Table S1. Flow cytometry analysis was performed on a BD FACSymphony A5 SORP and analysed with FlowJo software V10 6.1 (Treestar).
2.7. Confocal Immunofluorescence Histology and Histochemistry
Lungs of infected mice were collected and fixed in 4% paraformaldehyde (PFA). Samples were then dehydrated in 30% sucrose prior to embedding in OCT freezing media (Bio-Optica, Milan, Italy). Twenty micrometre sections were cut on a CM1520 cryostat (Leica, Wetzlar, Germany) and adhered to Superfrost Plus slides (Thermo Fisher Scientific, Waltham, MA, USA). Sections were then permeabilized and blocked in PBS containing 0.3% Triton X-100 (Sigma-Aldrich, St. Louis, MO, USA) and 5% FBS followed by staining in PBS containing 0.3% Triton X-100 and 1% FBS. Slides were stained for SARS-CoV-2 nucleocapsid (GeneTex, Irvine, CA, USA) for 1 h RT. Then, slides were stained with Alexa Fluor 568 Goat Anti-Rabbit antibody for 2 h RT. All slides were analysed by confocal fluorescence microscopy (Leica TCS SP5 Laser Scanning Confocal). For N-SARS-CoV-2 immunohistochemistry, mice were perfused with PBS and lungs were collected in Zn-formalin and transferred into 70% ethanol 24 h later. Tissue was then processed, embedded in paraffin and automatically stained for SARS-CoV-2 (2019-nCoV) Nucleocapsid Antibody (SINO BIO, 40143-R019, Beijing, China) through LEICA BOND RX 1 h RT and developed with Bond Polymer Refine Detection (Leica Biosystem, DS9800, Mannheim, Germany). Brightfield images were acquired through an Aperio Scanscope System CS2 microscope and an ImageScope program (Leica Biosystem, Mannheim, Germany) following the manufacturer’s instructions. In both immunofluorescence and histochemistry, the N-SARS-CoV-2 percentage of positive area was determined by QuPath V0.4.4 (Quantitative Pathology & Bioimage Analysis) software.
2.8. Clinical Score
Mice were observed daily for clinical symptoms. Disease severity was scored as reported in
Table 3.
2.9. Studies with hACE2-Transgenic B6 Mice Using Bioluminescent Reporter Viruses
All animals were maintained in the (SPF-free) barrier facility of the Yale University Animal Resource Centre (YARC) in a 14:10 light/dark cycle. All SARS-CoV-2-infected animals were housed in an animal room under BSL3 containment. The cages, animal waste, bedding, and carcasses of animals were disposed of and decontaminated in accordance with the guidelines set by Environmental Health Services at Yale. The replication-competent virus-infected animals were handled under ABSL3 conditions. All experiments described here were approved by Institutional Animal Care and Use Committee (IACUC) as well as SOPs approved by the Institutional Environmental Health and Biosafety committee. hACE2-transgenic B6 mice (heterozygous) were obtained from Jackson Laboratory. Six-to-eight-week-old male and female mice were used for all the experiments. The heterozygous mice were crossed and genotyped to select heterozygous mice for experiments by using the primer sets recommended by Jackson Laboratory. Each cohort size was n = 5 to allow for statistical testing. We calculated the number of animals (n = 4–8 per cohort) needed to achieve statistically significant results using an a priori power analysis. We calculated power and sample sizes required based on data from pilot experiments and previous studies [
20,
21,
22]. Animals with sex- and age-matched littermates were included randomly in the experiments. Animals were not excluded due to illness after the experiments. To ensure that the sex of the animals does not constitute a biological variable during analysis, equal numbers of male and female mice were included whenever possible.
2.10. Cell and Viruses
Vero E6 (CRL-1586, American Type Culture Collection (ATCC), were cultured at 37 °C in RPMI supplemented with 10% foetal bovine serum (FBS), 10 mM HEPES pH 7.3, 1 mM sodium pyruvate, 1× non-essential amino acids, and 100 U/mL of penicillin–streptomycin. SARS-CoV-2/USA_WA1/2019 isolate expressing nanoluc luciferase (nLuc) was obtained from Craig B Wilen, Yale University and generously provided by K. Plante and Pei-Yong Shi, World Reference Center for Emerging Viruses and Arboviruses, University of Texas Medical Branch [
23]. Viruses were propagated in Vero E6 TMPRSS2 by infecting them in T150 cm
2 flasks at an MOI of 0.1. The culture supernatants were collected after 18 h when cytopathic effects were clearly visible. The cell debris was removed by sedimentation and filtered through 0.45 micron filter to generate virus stocks. Viruses were concentrated by adding one volume of cold (4 °C) 4× PEG-it Virus Precipitation Solution (40% (
w/
v) PEG-8000 and 1.2 M NaCl; System Biosciences) to three volumes of virus-containing supernatant. The solution was mixed by inverting the tubes several times and then incubated at 4 °C overnight. The precipitated virus was harvested by centrifugation at 1500×
g for 60 min at 4 °C. The concentrated virus was then resuspended in PBS then aliquoted for storage at −80 °C. All work with infectious SARS-CoV-2 reporter viruses was performed in Institutional Biosafety Committee of Yale University (IBCYU)-approved BSL3 and A-BSL3 facilities at Yale University School of Medicine using appropriate positive pressure air respirators and protective equipment.
2.11. OMV Vaccination and SARS-CoV-2 Infection
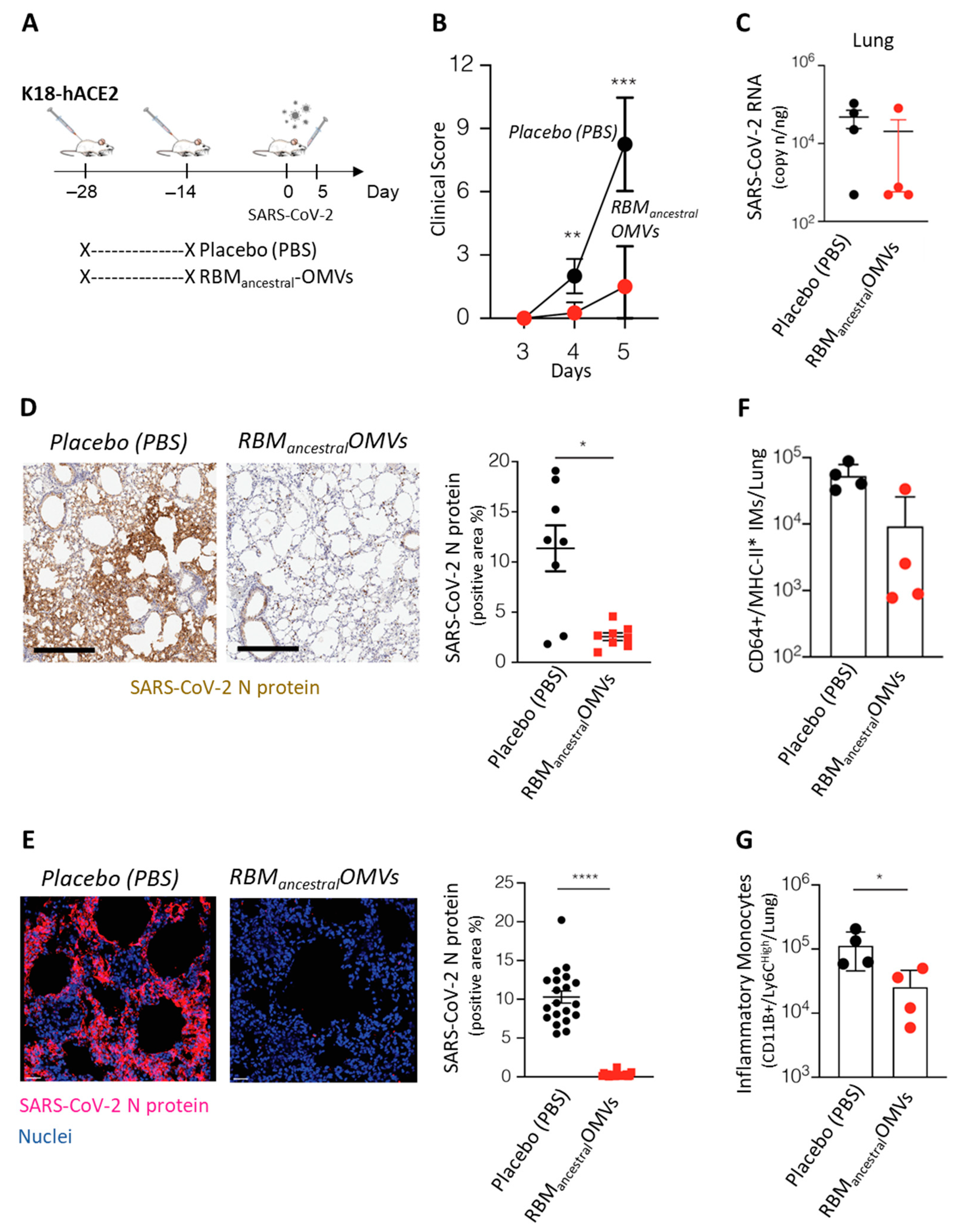
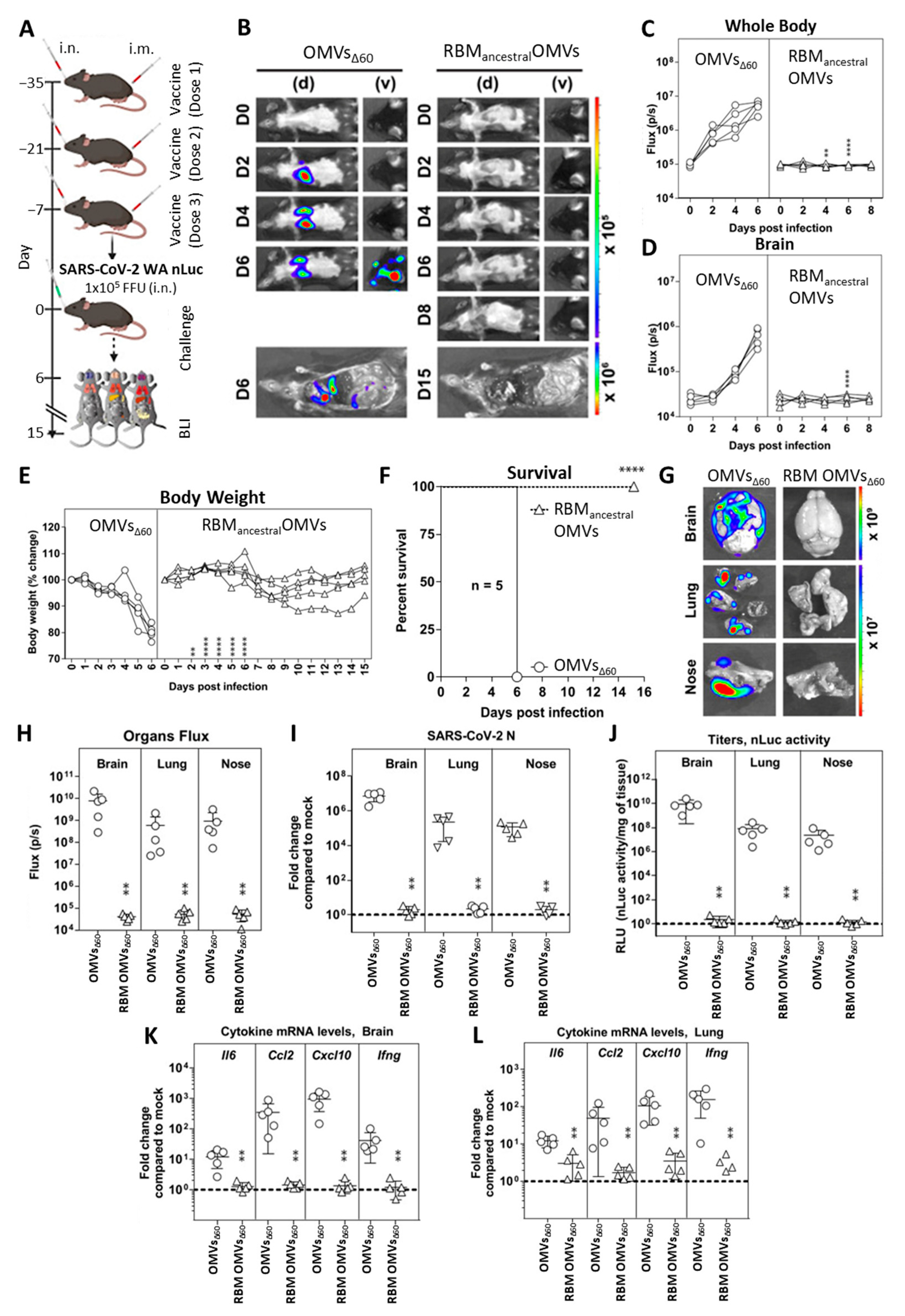
For all bioluminescence imaging-based in vivo experiments, groups of five 6-to-8-week-old K18 hACE-transgenic mice (both male and female) were immunized with 10 µg of either “empty OMVs” or RBMancestral-OMVs both intramuscularly (i.m., 50 µL per mouse; mixed with 2 mg/mL aluminium hydroxide (Alum) on the day of immunization, day 0) and intranasally (i.n., 25 µL, without any adjuvant). The animals were then boosted on day 14 and day 28 both i.m and i.n in a similar manner as primary vaccination. On day 35, the mice were intranasally challenged with 1 × 105 FFU SARS-CoV-2_WA1_nLuc in 25–30 µL volume under anaesthesia (0.5–5% isoflurane delivered using precision Dräger vaporizer with an oxygen flow rate of 1 L/min). The starting body weight was set to 100%. For survival experiments, mice were monitored every 8–12 h starting six days after virus challenge. Lethargic and moribund mice or mice that had lost more than 20% of their body weight were sacrificed and considered to have succumbed to infection for Kaplan–Meier survival plots. Mice were considered to have recovered if they gained back all the lost weight (experimental endpoint).
2.12. Bioluminescence Imaging (BLI) of SARS-CoV-2 Infection
All standard operating procedures and protocols for IVIS imaging of SARS-CoV-2-infected animals under ABSL-3 conditions were approved by IACUC, IBSCYU and Yale Animal Research Center (YARC). All the imaging was carried out using IVIS Spectrum® (Perkin Elmer, Waltham, MA, USA) in XIC-3 animal isolation chamber (Perkin Elmer, Waltham, MA, USA) that provided biological isolation of anesthetized mice or individual organs during the imaging procedure. All K18 hACE-transgenic mice were anesthetized via isoflurane inhalation (3–5% isoflurane, oxygen flow rate of 1.5 L/min) prior and during BLI using the XGI-8 Gas Anesthesia System. Prior to imaging, 100 µL of nanoluc luciferase (nLuc) substrate, furimazine (NanoGloTM, Promega, Madison, WI, USA) diluted 1:40 in endotoxin-free PBS was retro-orbitally administered to mice under anaesthesia. The mice were placed into XIC-3 animal isolation chamber (Perkin Elmer, Waltham, MA, USA) pre-saturated with isothesia and oxygen mix. The mice were imaged in both dorsal and ventral position at indicated days post infection. The animals were then imaged again after euthanasia and necropsy by spreading an additional 200 µL of substrate onto the exposed intact organs. Infected areas identified by carrying out whole-body imaging after necropsy were isolated, washed in PBS to remove residual blood and placed onto a clear plastic plate. Additional droplets of furimazine in PBS (1:40) were added to organs and soaked in substrate for 1–2 min before BLI. Images were acquired and analysed with Living Image v4.7.3 in vivo software package (Perkin Elmer, Waltham, MA, USA). Image acquisition exposures were set to auto, with imaging parameter preferences set in order of exposure time, binning, and f/stop, respectively. Images were acquired with luminescent f/stop of 2, photographic f/stop of 8. Binning was set to medium. Comparative images were compiled and batch-processed using the image browser with collective luminescent scales. Photon flux was measured as luminescent radiance (p/sec/cm2/sr). During luminescent threshold selection for image display, luminescent signals were regarded as background when minimum threshold setting resulted in displayed radiance above non-tissue-containing or known uninfected regions.
2.13. Focus Forming Assay
Titres of virus stocks were determined using a standard plaque assay. Briefly, the 4 × 105 Vero-E6 cells were seeded on a 12-well plate. Then, 24 h later, the cells were infected with 200 µL of serially diluted virus stock. After 1 h, the cells were overlayed with 1 mL of pre-warmed 0.6% Avicel (RC-581 FMC BioPolymer, Philadelphia, PA, USA) made in complete RPMI medium. Plaques were resolved at 48 h post infection by fixing in 10% paraformaldehyde for 15 min followed by staining for 20 min with 0.2% crystal violet made in 20% ethanol. Plates were rinsed in water to visualize plaques.
2.14. Measurement of Viral Burden
Indicated organs (nasal cavity, brain, and lungs) from infected or uninfected mice were collected, weighed, and homogenized in 1 mL of serum-free RPMI media containing penicillin–streptomycin and homogenized in a 2 mL tube containing 1.5 mm Zirconium beads with BeadBug 6 homogenizer (Benchmark Scientific, TEquipment Inc., Long Branch, NY, USA). Virus titres were measured using three highly correlative methods. First, the total RNA was extracted from homogenized tissues using an RNeasy plus Mini kit (Qiagen Cat # 74136, Hilden, Germany), reverse-transcribed with an iScript advanced cDNA kit (Bio-Rad Cat #1725036, Hercules, CA, USA) followed by a SYBR Green Real-time PCR assay for determining copies of SARS-CoV-2 N gene RNA using primers SARS-CoV-2 N F: 5′-ATGCTGCAATCGTGCTACAA-3′ and SARS-CoV-2 N R: 5′-GACTGCCGCCTCTGCTC-3′. All our real-time PCR assays based on SYBR Green had a built-in melt-curve that was checked to ensure estimation of only specific PCR products and not false positives. Second, we used Nanoluc (nLuc) luciferase activity as a shorter surrogate for plaque assay. Infected cells were washed with PBS and then lysed using 1× Passive lysis buffer. The lysates transferred into a 96-well solid white plate (Costar Inc., Washington, DC, USA) and nLuc activity was measured using a Tristar multiwell Luminometer (Berthold Technology, Bad Wildbad, Germany) for 2.5 s by adding 20 µL of Nano-Glo® substrate in a Nanoluc assay buffer (Promega Inc., Madison, WI, USA). Normalized relative light units were determined using uninfected monolayers of Vero cells treated identically. The data were processed and plotted using GraphPad Prism 8 v8.4.3.
2.15. Analyses of Signature Inflammatory Cytokines mRNA Expression
Brain and lung samples were collected from mice at the time of necropsy. Approximately 20 mg of tissue was suspended in 500 µL of RLT lysis buffer, and RNA was extracted using an RNeasy plus Mini kit (Qiagen Cat # 74136), reverse-transcribed with an iScript advanced cDNA kit (Bio-Rad Cat #1725036). To determine mRNA copy numbers of signature inflammatory cytokines, multiplex qPCR was conducted using iQ Multiplex Powermix (BioRad Cat # 1725848, Hercules, CA, USA) and PrimePCR Probe Assay mouse primers FAM-GAPDH, HEX-IL6, TEX615-CCL2, Cy5-CXCL10, and Cy5.5-IFNgamma. All PrimePCR Probe Assay mouse primers were purchased from Bio-Rad. The reaction plate was analysed using CFX96 touch real time PCR detection system. All channels were set to scan mode. The PCR conditions were 95 °C 2 min, 40 cycles of 95 °C for 10 s and 60 °C for 45 s, followed by a melting curve analysis to ensure that each primer pair resulted in amplification of a single PCR product. mRNA copy numbers of Il6, Ccl2, Cxcl10 and Ifng in the cDNA samples of infected mice were normalized to Gapdh mRNA with the formula ΔCt(target gene) = Ct(target gene) − Ct(Gapdh). The fold increase was determined using the 2−ΔΔCt method comparing treated mice to uninfected controls.
2.16. ELISA
Ninety-six-well Maxisorp plates (Nunc, Thermo Fisher Scientific) were coated with 200 ng/well of purified recombinant RBD and blocked with 100 mL/well of PBS + 1% BSA. Mice sera were threefold serially diluted in PBS + 1% BSA starting from a 1:100 initial dilution and added in each well. Goat anti-mouse alkaline phosphatase–conjugate antibodies at a final dilution of 1:2000 (Sigma Aldrich, Burlington, MA, USA) were used. Finally, a 100 mL/well of 3 mg/mL paranitrophenyl-phosphate disodium hexahydrate (Sigma Aldrich Burlington, MA, USA) in 1 M diethanolamine buffer pH 9.8 and plates were incubated at room temperature in the dark for 45 min. Absorbance was read at 405 nm using Varioskan™ LUX multimode (Thermo Fisher Scientific, Waltham, MA, USA) microplate reader.
2.17. Preparation of Viral Pseudotypes and Neutralization Assays
Lentiviral particles pseudotyped with SARS-CoV-2 spike were produced in 10 cm plates prepared the day before transfection with 3 million HEK293T cells in 10 mL complete DMEM, supplemented with 10% FBS. Simian Immunodeficiency virus (SIV)-based vectors were produced by transfecting cells using the Calcium Phosphate method with 17.5 μg of env-defective SIV-Mac239-GFP construct with GFP expressed in place of Nef and 2.5 μg of PCDNA3.1 encoding the WT SARS-CoV-2 spike (reference sequence Wuhan-Hu-1, accession number YP_009724390) engineered to truncate the C-terminal 19 amino acids. Pseudotyped vector supernatants were harvested 48 h post-transfection and filtered through a 0.45 μm filter before. Neutralization titres were tested on Huh-7 cells overexpressing ACE2. Target cells were seeded onto a 96-well tissue culture one day before neutralization. The vector inoculum was adjusted to produce no more than 10% transduction of the monolayer to ensure a linear working range of the assay.
Sera dilutions were added to pseudotyped virus particles, incubated at room temperature for 30 min and added to cells. After 48 h, transduction was assessed by counting the GFP-expressing cells using the fluorescent plate reader Ensight (Perkin Elmer, Waltham, MA, USA). Each serum dilution was assessed in triplicate. Neutralization was measured by calculating the residual transduction activity of the pseudovirus considering the untreated sample as 100%. Fitted sigmoidal curves and IC50 were obtained using Prism (Graphpad) with the least-squares variable slope method and using the dose-normalized response protocol.
2.18. Statistical Analyses and Software
Detailed information concerning the statistical methods used is provided in the figure legends. Flow data were collected using FlowJo Version 10.5.3 (Treestar). Statistical analyses were performed with GraphPad Prism software version 8 (GraphPad). n represents individual mice analysed per experiments. Error bars indicate the standard error of the mean (SEM).
4. Discussion
We demonstrate for the first time that engineered
E. coli OMVs can provide an ideal platform for the development of an effective SARS-CoV-2 vaccine. While properly folded eukaryotic glycoproteins can be typically expressed only in eukaryotic expression systems, the crucial portion of the Spike RBM, which directly contacts ACE2, can be efficiently incorporated into OMVs. An effective immunity is elicited in animals vaccinated with RBM
ancestral-OMVs, with the production of neutralizing antibodies at levels sufficient to protect hACE2-transgenic mice from infection. This is a very exciting result, considering that OMVs are extremely easy to produce, upscale and distribute. The separation of the biomass from the culture supernatant and an ultrafiltration step to concentrate OMVs and eliminate contaminants is essentially all that is needed for vaccine production [
16]. Crucially, the same process can be applied to any antigen and different vaccines and is amenable to large-scale production which can be easily transferred to different facilities worldwide. Moreover, the production yields make the vaccine costs particularly affordable. Under laboratory conditions, we reproducibly obtain more than 5000 OMV-based vaccine doses/litre of culture. Practically speaking, this means that with a 1.000 litre fermentation unit associated with a tangential flow ultrafiltration device it is possible to provide >1 million doses of vaccine/week at costs that are expected to be well below 1 USD/dose. Given the ease of production, upscaling, and product stability, OMV vaccines should favourably compete with RNA-based technologies.
The intranasal delivery of the RBM
ancestral-OMV vaccine is likely to be effective in preventing infection in the upper respiratory tract and viral dissemination in different organs and tissues, including the brain. This can be at least partially inferred from the results of the two animal experiments described in this work. In the first set of experiments, we immunized hACE2-transgenic mice systemically and subsequently challenged them intranasally with the ancestral strain. Three out of four animals survived, and viruses were not detected, using both immunohistochemistry and PCR, in the lungs of the protected mice. However, the virus could be found in the brain tissue of both protected and mock-immunized mice. In the second set of experiments, the RBM
ancestral-OMV vaccine was given both systemically and intranasally before challenge. Following this immunization schedule, all five immunized animals under this regimen were completely protected, and the virus could not be detected in the lung, brain and nose of the animals. Moreover, protection strongly correlated with the absence of inflammatory cytokines in the analysed tissues. Therefore, although we have not yet tested the intranasal immunization alone, the data suggest that mucosal delivery contributes substantially to overall protection. Such data would be in line with the data obtained with a mucosal vaccine based on the OMVs from
Neisseria meningitidis combined with recombinant Spike protein [
42].
Our work also showed that the RBM
ancestral-OMVs vaccine elicits cross-neutralizing antibodies against different VOCs. The OMVs platform is flexible enough to be rapidly adapted to cope with new emerging variants. In addition to the RBM of the ancestral and BA.1 strain, we have successfully engineered our OMVs with the RBM from the P1 and B.1.617 isolates (not shown). All engineered OMVs elicited high levels of antibodies, which effectively neutralized the homologous pseudoviruses in vitro. Moreover, we found that the antibodies induced by the RBM
ancestral-OMVs vaccine provided robust cross-protection against the most recent Omicron variants. This is remarkable, considering that the vaccines and the neutralizing monoclonal antibodies based on the ancestral Spike protein are known to be poorly effective against Omicron variants [
40]. When expressed on OMVs, the RBM of the ancestral strain, but not the RBM of the BA.1 strain, presents otherwise poorly immunogenic epitopes, which induce antibodies against conserved structural regions of the RBD involved in the interaction with the ACE2 receptor. It would be of particular interest to isolate the B cells from the RBM
ancestral-OMVs immunized mice and characterize these putative new classes of antibodies. The existence of a new family of neutralized antibodies would be of particular interest from a vaccination standpoint. Nowadays, most of the worldwide population has been either infected by SARS-CoV-2 or vaccinated. Therefore, the immunization with the RBM
ancestral-OMVs vaccine should synergize with the natural immunity of the vaccinees, thus making the vaccination particularly effective.
While further experiments are needed to test the efficacy against a larger panel of SARS-CoV-2 variants, our data suggest that the RBMancestral-OMVs vaccine has the potential to become a broadly protective vaccine that, thanks to its ease of manufacturing and low production costs, could be produced and distributed in low-income countries.


 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}