
Evaluation of Two Adjuvant Formulations for an Inactivated Yellow Fever 17DD Vaccine Candidate in Mice

 , , , , ,
, , , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells and 17DD Virus Production
2.2. 17DD Virus Purification and Inactivation
2.3. Analytical Methods
2.4. Animals and Experimental Procedures
2.5. Immunogenicity and Protection
2.6. Plaque Reduction Neutralization Test (PRNT50)
2.7. Total IgG by ELISA
IgG Subtypes
2.8. Cellular Response
2.8.1. Cytokine ELISpot Assays
2.8.2. IgG ELISpot Assay
2.9. Statistical Analysis
3. Results
3.1. Protection after Lethal Challenge
3.2. Immunogenicity of YF 17DD Inactivated Vaccine Formulations
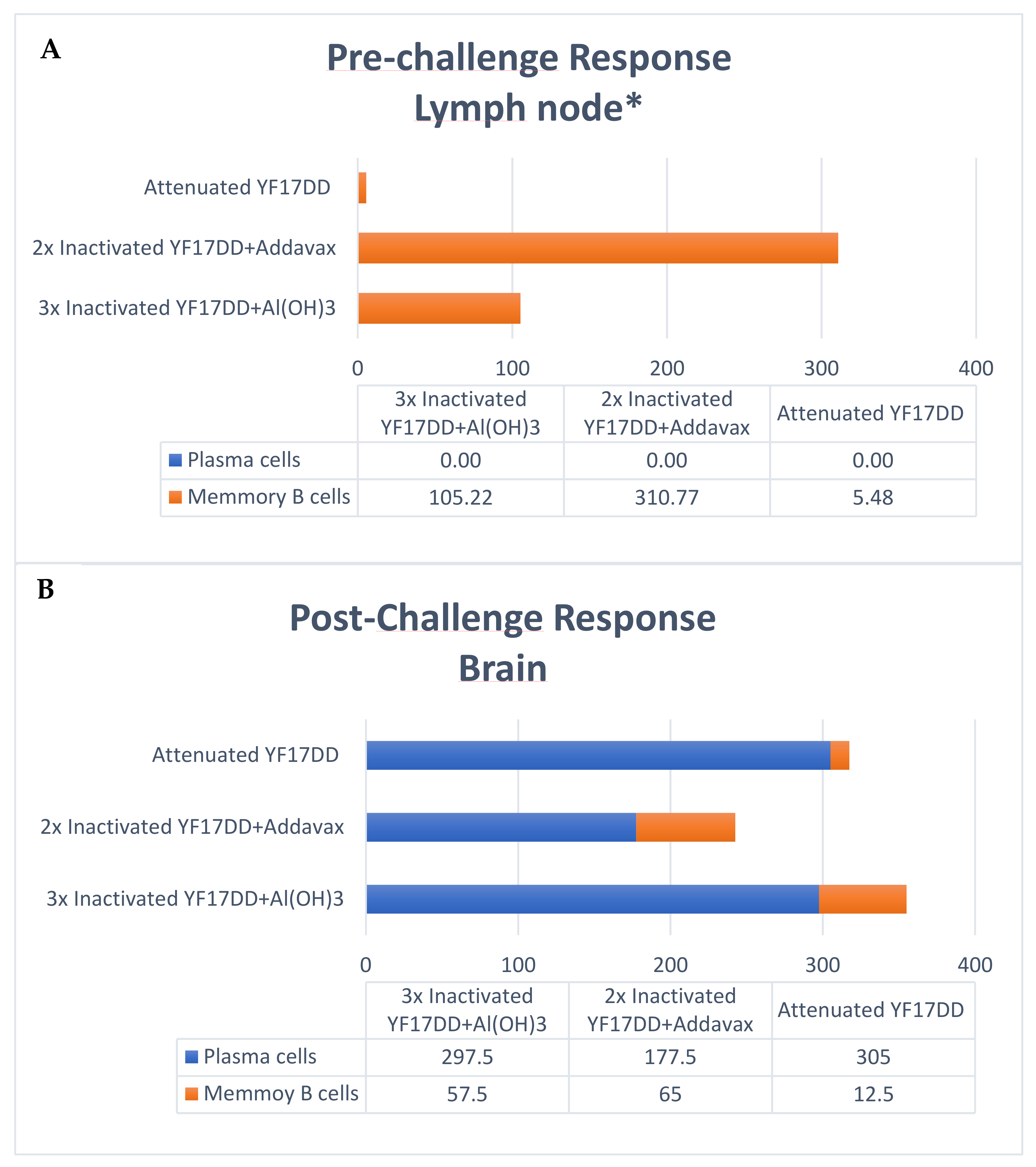
3.3. Cellular Responses
IgG ELISpot
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- De Oliveira Figueiredo, P.; Stoffella-Dutra, A.G.; Barbosa Costa, G.; Silva de Oliveira, J.; Dourado Amaral, C.; Duarte Santos, J.; Soares Rocha, K.L.; Araújo Júnior, J.P.; Lacerda Nogueira, M.; Zazá Borges, M.A.; et al. Re-Emergence of Yellow Fever in Brazil during 2016–2019: Challenges, Lessons Learned, and Perspectives. Viruses 2020, 12, 1233. [Google Scholar] [CrossRef] [PubMed]
- Beck, A.S.; Barrett, A.D.T. Current status and future prospects of yellow fever vaccines. Expert Rev. Vaccines 2015, 14, 1479–1492. [Google Scholar] [CrossRef] [PubMed]
- Vratskikh, O.; Stiasny, K.; Zlatkovic, J.; Tsouchnikas, G.; Jarmer, J.; Karrer, U.; Roggendorf, M.; Roggendorf, H.; Allwinn, R.; Heinz, F.X. Dissection of antibody specificities induced by yellow fever vaccination. PLoS Pathog. 2013, 9, e1003458. [Google Scholar] [CrossRef] [PubMed]
- Bastola, R.; Noh, G.; Keum, T.; Bashyal, S.; Seo, J.-E.; Choi, J.; Oh, Y.; Cho, Y.; Lee, S. Vaccine adjuvants: Smart components to boost the immune system. Arch. Pharm. Res. 2017, 40, 1238–1248. [Google Scholar] [CrossRef] [PubMed]
- Souza, M.C.O.; Freire, M.S.; Schulze, E.A.; Gaspar, L.P.; Castilho, L.R. Production of yellow fever virus in microcarrier-based Vero cell cultures. Vaccine 2009, 27, 6420–6423. [Google Scholar] [CrossRef]
- Mattos, D.A.; Silva, M.V.; Gaspar, L.P.; Castilho, L.R. Increasing Vero viable cell densities for yellow fever virus production in stirred-tank bioreactors using serum-free medium. Vaccine 2015, 33, 4288–4291. [Google Scholar] [CrossRef]
- Pato, T.P.; Souza, M.C.O.; Silva, A.N.M.R.; Pereira, R.C.; Silva, M.V.; Caride, E.; Gaspar, L.P.; Freire, M.S.; Castilho, L.R. Development of a membrane adsorber based capture step for the purification of yellow fever virus. Vaccine 2014, 32, 2789–2793. [Google Scholar] [CrossRef]
- Pato, T.P.; Souza, M.C.O.; Mattos, D.A.; Caride, E.; Ferreira, D.F.; Gaspar, L.P.; Freire, M.S.; Castilho, L.R. Purification of yellow fever virus produced in Vero cells for inactivated vaccine manufacture. Vaccine 2019, 37, 3214–3220. [Google Scholar] [CrossRef]
- Pereira, R.C.; Silva, A.N.M.R.; Souza, M.C.O.; Silva, M.V.; Neves, P.P.C.C.; Silva, A.A.M.V.; Matos, D.D.C.S.; Herrera, M.A.O.; Yamamura, A.M.Y.; Freire, M.S.; et al. An inactivated yellow fever 17DD vaccine cultivated in Vero cell cultures. Vaccine 2015, 33, 4261–4268. [Google Scholar] [CrossRef]
- Monath, T.P.; Lee, C.K.; Julander, J.G.; Brown, A.; Beasley, D.W.; Watts, D.M.; Hayman, E.; Guertin, P.; Makowiecki, J.; Crowell, J.; et al. Inactivated yellow fever 17D vaccine: Development and nonclinical safety, immunogenicity and protective activity. Vaccine 2010, 28, 3827–3840. [Google Scholar] [CrossRef]
- Monath, T.P.; Fowler, E.; Johnson, C.T.; Balser, J.; Morin, M.J.; Sisti, M.; Trent, D.W. An inactivated cell-culture vaccine against yellow fever. N. Engl. J. Med. 2011, 364, 1326–1333. [Google Scholar] [CrossRef] [PubMed]
- Julander, J.G.; Trent, D.W.; Monath, T.P. Immune correlates of protection against yellow fever determined by passive immunization and challenge in the hamster model. Vaccine 2011, 29, 6008–6016. [Google Scholar] [CrossRef] [PubMed]
- Barrett, P.N.; Terpening, S.J.; Snow, D.; Cobb, R.R.; Kistner, O. Vero cell technology for rapid development of inactivated whole virus vaccines for emerging viral diseases. Expert Rev. Vaccines 2017, 16, 883–894. [Google Scholar] [CrossRef] [PubMed]
- Kiesslich, S.; Kamen, A.A. Vero cell upstream bioprocess development for the production of viral vectors and vaccines. Biotechnol. Adv. 2020, 44, 107608. [Google Scholar] [CrossRef] [PubMed]
- Caufour, P.S.; Motta, M.C.; Yamamura, A.M.; Vazquez, S.; Ferreira, I.I.; Jabor, A.V.; Bonaldo, M.C.; Freire, M.S.; Galler, R. Construction, characterization and immunogenicity of recombinant yellow fever 17D-dengue type 2 viruses. Virus Res. 2001, 79, 1–14. [Google Scholar] [CrossRef]
- Simões, M.; Camacho, L.A.B.; Yamamura, A.M.Y.; Miranda, E.H.; Cajaraville, A.C.R.A.; da Silva Freire, M. Evaluation of accuracy and reliability of the plaque reduction neutralization test (micro-PRNT) in detection of yellow fever virus antibodies. Biol. J. Int. Assoc. Biol. Stand. 2012, 40, 399–404. [Google Scholar] [CrossRef]
- Maciel, M.; da Silva Pereira Cruz, F.; Cordeiro, M.T.; da Motta, M.A.; de Melo Cassemiro, K.M.S.; de Cássia Carvalho Maia, R.; de Figueiredo, R.C.B.Q.; Galler, R.; da Silva Freire, M.; August, J.T.; et al. A DNA Vaccine against Yellow Fever Virus: Development and Evaluation. PLoS Negl. Trop. Dis. 2015, 9, e0003693. [Google Scholar] [CrossRef]
- Pino, P.A.; Cardona, A.E. Isolation of brain and spinal cord mononuclear cells using percoll gradients. J. Vis. Exp. 2011, 48, e2348. [Google Scholar] [CrossRef]
- Pulendran, B. Learning immunology from the yellow fever vaccine: Innate immunity to systems vaccinology. Nat. Rev. Immunol. 2009, 9, 741–747. [Google Scholar] [CrossRef]
- Da Costa-Rocha, I.A.; Campi-Azevedo, A.C.; Peruhype-Magalhães, V.; Coelho-Dos-Reis, J.G.; Fradico, J.R.B.; Souza-Lopes, T.; Reis, L.R.; Freire, L.C.; Costa-Pereira, C.; de Melo Mambrini, J.V.; et al. Duration of Humoral and Cellular Immunity 8 Years After Administration of Reduced Doses of the 17DD-Yellow Fever Vaccine. Front. Immunol. 2019, 10, 1211. [Google Scholar] [CrossRef]
- Reis, L.R.; da Costa-Rocha, I.A.; Campi-Azevedo, A.C.; Peruhype-Magalhães, V.; Coelho-dos-Reis, J.G.; Costa-Pereira, C.; Otta, D.A.; Freire, L.C.; de Lima, S.M.; de Souza Azevedo, A.; et al. Exploratory study of humoral and cellular immunity to 17DD Yellow Fever vaccination in children and adults residents of areas without circulation of Yellow Fever Virus. Vaccine 2022, 40, 798–810. [Google Scholar] [CrossRef] [PubMed]
- Martins, M.A.; Silva, M.L.; Elói-Santos, S.M.; Ribeiro, J.G.L.; Peruhype-Magalhães, V.; Marciano, A.P.V.; Homma, A.; Kroon, E.G.; Teixeira-Carvalho, A.; Martins-Filho, O.A. Innate immunity phenotypic features point toward simultaneous raise of activation and modulation events following 17DD live attenuated yellow fever first-time vaccination. Vaccine 2008, 26, 1173–1184. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.L.; Martins, M.A.; Espírito-Santo, L.R.; Campi-Azevedo, A.C.; Silveira-Lemos, D.; Ribeiro, J.G.L.; Homma, A.; Kroon, E.G.; Teixeira-Carvalho, A.; Elói-Santos, S.M.; et al. Characterization of main cytokine sources from the innate and adaptive immune responses following primary 17DD yellow fever vaccination in adults. Vaccine 2011, 29, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Germann, T.; Bongartz, M.; Dlugonska, H.; Hess, H.; Schmitt, E.; Kolbe, L.; Kölsch, E.; Podlaski, F.J.; Gately, M.K.; Rüde, E. Interleukin-12 profoundly up-regulates the synthesis of antigen-specific complement-fixing IgG2a, IgG2b and IgG3 antibody subclasses in vivo. Eur. J. Immunol. 1995, 25, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Neves, P.C.C.; Rudersdorf, R.A.; Galler, R.; Bonaldo, M.C.; de Santana, M.G.V.; Mudd, P.A.; Martins, M.A.; Rakasz, E.G.; Wilson, N.A.; Watkins, D.I. CD8+ gamma-delta TCR+ and CD4+ T cells produce IFN-γ at 5-7 days after yellow fever vaccination in Indian rhesus macaques, before the induction of classical antigen-specific T cell responses. Vaccine 2010, 28, 8183–8188. [Google Scholar] [CrossRef] [PubMed]
- Dowd, K.A.; Pierson, T.C. Antibody-mediated neutralization of flaviviruses: A reductionist view. Virology 2011, 411, 306–315. [Google Scholar] [CrossRef]
- Fox, C.B.; Barnes, V.L.; Evers, T.; Chesko, J.D.; Vedvick, T.S.; Coler, R.N.; Reed, S.G.; Baldwin, S.L. Adjuvanted pandemic influenza vaccine: Variation of emulsion components affects stability, antigen structure, and vaccine efficacy. Influenza Other Respir. Viruses 2013, 7, 815–826. [Google Scholar] [CrossRef]
- Miller, J.D.; van der Most, R.G.; Akondy, R.S.; Glidewell, J.T.; Albott, S.; Masopust, D.; Murali-Krishna, K.; Mahar, P.L.; Edupuganti, S.; Lalor, S.; et al. Human effector and memory CD8+ T cell responses to smallpox and yellow fever vaccines. Immunity 2008, 28, 710–722. [Google Scholar] [CrossRef]
- Akondy, R.S.; Monson, N.D.; Miller, J.D.; Edupuganti, S.; Teuwen, D.; Wu, H.; Quyyumi, F.; Garg, S.; Altman, J.D.; Del Rio, C.; et al. The yellow fever virus vaccine induces a broad and polyfunctional human memory CD8+ T cell response. J. Immunol. 2009, 183, 7919–7930. [Google Scholar] [CrossRef]
- Blom, K.; Braun, M.; Ivarsson, M.A.; Gonzalez, V.D.; Falconer, K.; Moll, M.; Ljunggren, H.-G.; Michaëlsson, J.; Sandberg, J.K. Temporal dynamics of the primary human T cell response to yellow fever virus 17D as it matures from an effector- to a memory-type response. J. Immunol. 2013, 190, 2150–2158. [Google Scholar] [CrossRef]
- Neves, P.C.C.; Santos, J.R.; Tubarão, L.N.; Bonaldo, M.C.; Galler, R. Early IFN-gamma production after YF 17D vaccine virus immunization in mice and its association with adaptive immune responses. PLoS ONE 2013, 8, e81953. [Google Scholar] [CrossRef] [PubMed]
- Bassi, M.R.; Kongsgaard, M.; Steffensen, M.A.; Fenger, C.; Rasmussen, M.; Skjødt, K.; Finsen, B.; Stryhn, A.; Buus, S.; Christensen, J.P.; et al. CD8+ T cells complement antibodies in protecting against yellow fever virus. J. Immunol. 2015, 194, 1141–1153. [Google Scholar] [CrossRef] [PubMed]
- Neves, P.C.d.C.; de Souza Matos, D.C.; Marcovistz, R.; Galler, R. TLR expression and NK cell activation after human yellow fever vaccination. Vaccine 2009, 27, 5543–5549. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Chambers, T.J. Yellow fever virus encephalitis: Properties of the brain-associated T-cell response during virus clearance in normal and gamma interferon-deficient mice and requirement for CD4+ lymphocytes. J. Virol. 2001, 75, 2107–2118. [Google Scholar] [CrossRef]
- Lam, L.K.M.; Watson, A.M.; Ryman, K.D.; Klimstra, W.B. Gamma-interferon exerts a critical early restriction on replication and dissemination of yellow fever virus vaccine strain 17D-204. NPJ Vaccines 2018, 3, 5. [Google Scholar] [CrossRef]
- De Silva, N.S.; Klein, U. Dynamics of B cells in germinal centres. Nat. Rev. Immunol. 2015, 15, 137–148. [Google Scholar] [CrossRef]
- Bergmann, B.; Grimsholm, O.; Thorarinsdottir, K.; Ren, W.; Jirholt, P.; Gjertsson, I.; Mårtensson, I.-L. Memory B cells in mouse models. Scand. J. Immunol. 2013, 78, 149–156. [Google Scholar] [CrossRef]
- Shapiro-Shelef, M.; Calame, K. Regulation of plasma-cell development. Nat. Rev. Immunol. 2005, 5, 230–242. [Google Scholar] [CrossRef]
- Iwasaki, A. Immune Regulation of Antibody Access to Neuronal Tissues. Trends Mol. Med. 2017, 23, 227–245. [Google Scholar] [CrossRef]
- Del Giudice, G.; Rappuoli, R.; Didierlaurent, A.M. Correlates of adjuvanticity: A review on adjuvants in licensed vaccines. Semin. Immunol. 2018, 39, 14–21. [Google Scholar] [CrossRef]
- Vesikari, T.; Knuf, M.; Wutzler, P.; Karvonen, A.; Kieninger-Baum, D.; Schmitt, H.J.; Baehner, F.; Borkowski, A.; Tsai, T.F.; Clemens, R. Oil-in-water emulsion adjuvant with influenza vaccine in young children. N. Engl. J. Med. 2011, 365, 1406–1416. [Google Scholar] [CrossRef] [PubMed]
- An Overview of Vaccine Adjuvants: Current Evidence and Future Perspectives—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/35632575/ (accessed on 8 September 2022).
- Pulendran, B.; SArunachalam, P.; O’Hagan, D.T. Emerging concepts in the science of vaccine adjuvants. Nat. Rev. Drug Discov. 2021, 20, 454–475. [Google Scholar] [CrossRef] [PubMed]
- O’Hagan, D.T.; Ott, G.S.; De Gregorio, E.; Seubert, A. The mechanism of action of MF59—An innately attractive adjuvant formulation. Vaccine 2012, 30, 4341–4348. [Google Scholar] [CrossRef] [PubMed]
- Cioncada, R.; Maddaluno, M.; Vo, H.T.M.; Woodruff, M.; Tavarini, S.; Sammicheli, C.; Tortoli, M.; Pezzicoli, A.; De Gregorio, E.; Carroll, M.C.; et al. Vaccine adjuvant MF59 promotes the intranodal differentiation of antigen-loaded and activated monocyte-derived dendritic cells. PLoS ONE 2017, 12, e0185843. [Google Scholar] [CrossRef] [PubMed]
- O’Hagan, D.T.; van der Most, R.; Lodaya, R.N.; Coccia, M.; Lofano, G. “World in motion”—Emulsion adjuvants rising to meet the pandemic challenges. NPJ Vaccines 2021, 6, 158. [Google Scholar] [CrossRef]
- Lofano, G.; Mancini, F.; Salvatore, G.; Cantisani, R.; Monaci, E.; Carrisi, C.; Tavarini, S.; Sammicheli, C.; Rossi Paccani, S.; Soldaini, E.; et al. Oil-in-Water Emulsion MF59 Increases Germinal Center B Cell Differentiation and Persistence in Response to Vaccination. J. Immunol. 2015, 195, 1617–1627. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Formulation | Antigen (µg) | Regiment | Survival Rate: Alive/Total (%) | ||
|---|---|---|---|---|---|
| Experiment I | Experiment II | Mean * | |||
| Mock | x | 3-dose | 0/9 (0%) | x | 0/9 (0%) |
| Al(OH)3 | x | 3-dose | 0/8 ** (0%) | x | 0/8 * (0%) |
| AddaVax® | x | 2-dose | 0/9 (0%) | x | 0/9 (0%) |
| Inactivated YFV+ Al(OH)3 0.3% | 10 | 3-dose | 7/9 (77.8%) | 5/8 (62.5%) | 12/17 (70.6%) |
| Inactivated YFV +AddaVax ® 50% | 10 | 2-dose | 8/8 * (100%) | 8/8 (100%) | 16/16 * (100%) |
| Attenuated YF 17DD | 105 PFU | 1-dose | 9/9 (100%) | 8/8 (100%) | 17/17 (100%) |
| Group | Formulation | Nabs (log10 1/x) | Total IgG (log10 µg/mL) | ||
|---|---|---|---|---|---|
| Pre- | Post- | Pre- | Post- | ||
| 1 | Mock | 1.12 ± 0.17 | x | 0.11 | x |
| 2 | Al(OH)3 | 0.89 ± 0.15 | x | 0.19 | x |
| 3 | AddaVax® | 0.81 ± 0.13 | x | 0.10 | x |
| 4 | 3x Inactivated YF 17DD (10 µg) + Al(OH)3 | 0.92 ± 0.11 | 2.51 ± 0.18 | 3.12 ± 0.09 | 3.14 ± 0.11 |
| 5 | 2x Inactivated YF 17DD (10 µg) + AddaVax® | 0.88 ± 0.11 | 1.64 ± 0.32 | 3.81 ± 0.07 | 3.67 ± 0.12 |
| 6 | 1x Attenuated YF17DD | 1.60 ± 0.24 | 2.69 ± 0.28 | 1.07 ± 0.08 | 2.43 ± 0.23 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cajaraville, A.C.d.R.A.; Gomes, M.P.d.B.; Azamor, T.; Pereira, R.C.; Neves, P.C.d.C.; De Luca, P.M.; Lima, S.M.B.d.; Gaspar, L.P.; Caride, E.; Freire, M.d.S.; et al. Evaluation of Two Adjuvant Formulations for an Inactivated Yellow Fever 17DD Vaccine Candidate in Mice. Vaccines 2023, 11, 73. https://doi.org/10.3390/vaccines11010073
Cajaraville ACdRA, Gomes MPdB, Azamor T, Pereira RC, Neves PCdC, De Luca PM, Lima SMBd, Gaspar LP, Caride E, Freire MdS, et al. Evaluation of Two Adjuvant Formulations for an Inactivated Yellow Fever 17DD Vaccine Candidate in Mice. Vaccines. 2023; 11(1):73. https://doi.org/10.3390/vaccines11010073
Chicago/Turabian StyleCajaraville, Ana Carolina dos Reis Albuquerque, Mariana Pierre de Barros Gomes, Tamiris Azamor, Renata Carvalho Pereira, Patrícia Cristina da Costa Neves, Paula Mello De Luca, Sheila Maria Barbosa de Lima, Luciane Pinto Gaspar, Elena Caride, Marcos da Silva Freire, and et al. 2023. "Evaluation of Two Adjuvant Formulations for an Inactivated Yellow Fever 17DD Vaccine Candidate in Mice" Vaccines 11, no. 1: 73. https://doi.org/10.3390/vaccines11010073
APA StyleCajaraville, A. C. d. R. A., Gomes, M. P. d. B., Azamor, T., Pereira, R. C., Neves, P. C. d. C., De Luca, P. M., Lima, S. M. B. d., Gaspar, L. P., Caride, E., Freire, M. d. S., & Medeiros, M. A. (2023). Evaluation of Two Adjuvant Formulations for an Inactivated Yellow Fever 17DD Vaccine Candidate in Mice. Vaccines, 11(1), 73. https://doi.org/10.3390/vaccines11010073