
A Closer Look at ACE2 Signaling Pathway and Processing during COVID-19 Infection: Identifying Possible Targets
 , , and
, , and
Abstract
1. Introduction
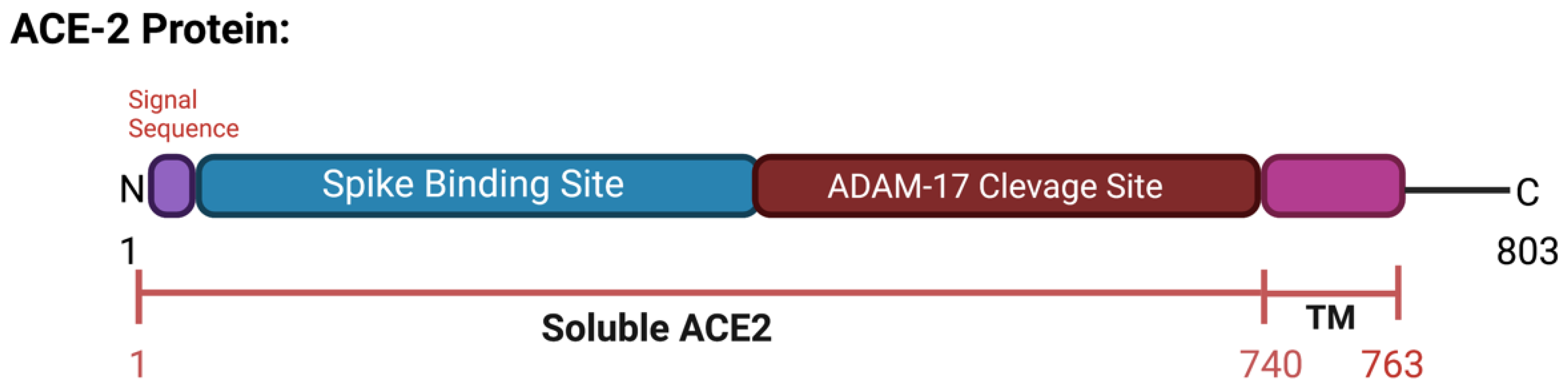
2. ACE2 Protein
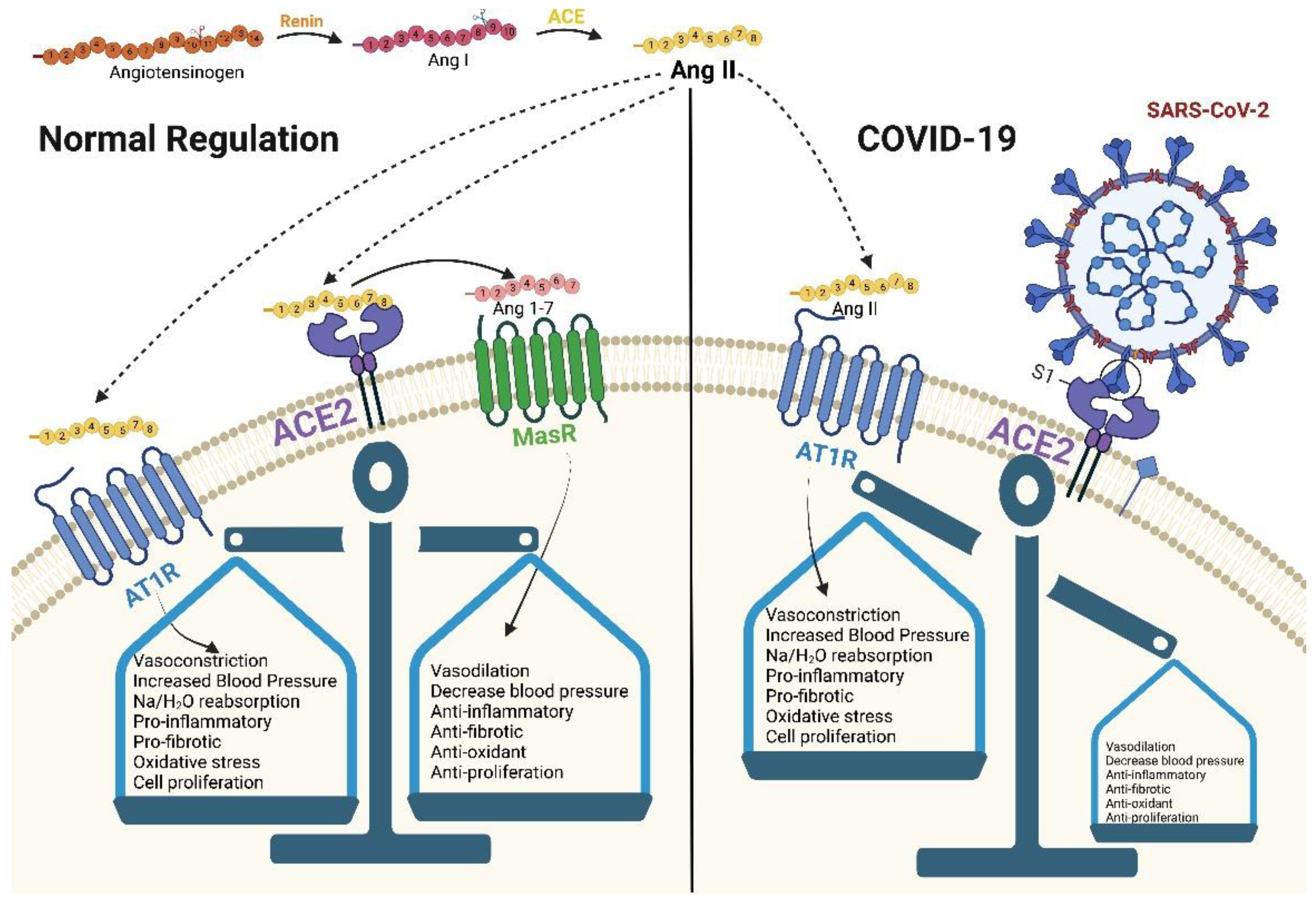
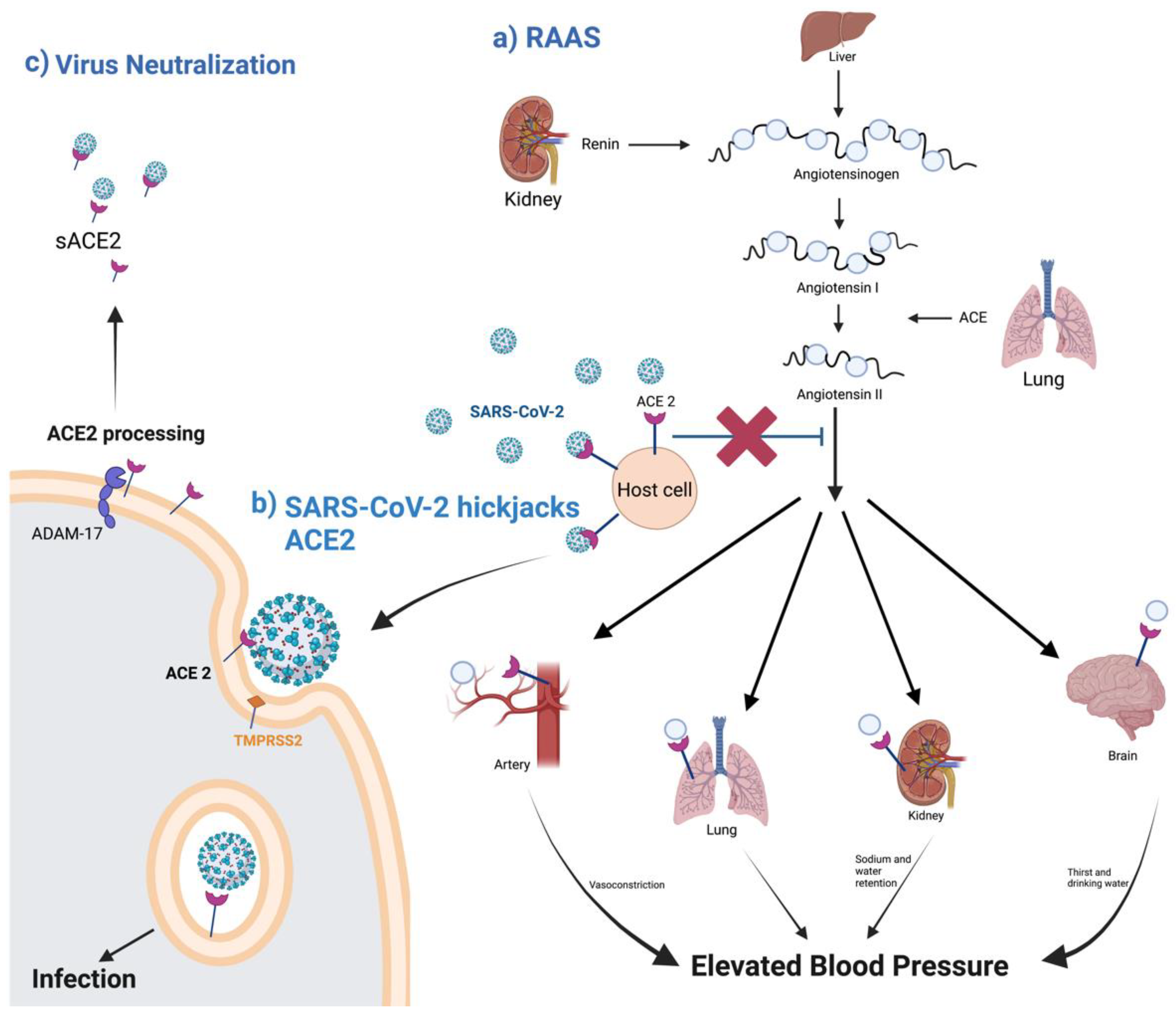
2.1. ACE2 and RAAS
2.2. ACE2 Processing during SARS-CoV-2 Infection
2.3. ACE Isoforms
3. ACE2 Variants and COVID-19 Susceptibility
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, S.; Sreenivasulu, K.; Mitra, P.; Misra, S.; Sharma, A.P. Role of Genetic Variants and Gene Expression in the Susceptibility and Severity of COVID-19. Ann. Lab. Med. 2021, 41, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Keidar, S.; Kaplan, M.; Gamliel-Lazarovich, A. ACE2 of the heart: From angiotensin I to angiotensin (1–7). Cardiovasc. Res. 2007, 73, 463–469. [Google Scholar] [CrossRef]
- Chambers, S.; Bhatia, M. ACE and ACE2 in Inflammation: A Tale of Two Enzymes. Inflamm. Allergy-Drug Targets 2014, 13, 224–234. [Google Scholar] [CrossRef]
- Wang, K.; Viveiros, A.; Nguyen, Q.; Zhong, J.; Turner, A.J.; Raizada, M.K.; Grant, M.B.; Oudit, G.Y. Angiotensin-Converting Enzyme 2: SARS-CoV-2 Receptor and Regulator of the Renin-Angiotensin System. Circ. Res. 2020, 126, 1456–1474. [Google Scholar]
- Palau, V.; Pascual, J.; Soler, M.J.; Riera, M. Role of Adam17 in Kidney Disease. Am. J. Physiol.-Ren. Physiol. 2019, 317, F333–F342. [Google Scholar] [CrossRef]
- Stephany, B.; Casaro, E.B.; Rangel, É.B. COVID-19: Angiotensin-Converting Enzyme 2 (Ace2) Expression and Tissue Susceptibility to SARS-CoV-2 Infection. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 905–919. [Google Scholar]
- Osman, I.O.; Melenotte, C.; Brouqui, P.; Million, M.; Lagier, J.-C.; Parola, P.; Stein, A.; La Scola, B.; Meddeb, L.; Mege, J.-L.; et al. Expression of ACE2, Soluble ACE2, Angiotensin I, Angiotensin II and Angiotensin-(1-7) Is Modulated in COVID-19 Patients. Front. Immunol. 2021, 12, 625732. [Google Scholar] [CrossRef]
- Bindom, S.M.; Hans, C.P.; Xia, H.; Boulares, A.H.; Lazartigues, E. Angiotensin I–Converting En-zyme Type 2 (Ace2) Gene Therapy Improves Glycemic Control in Diabetic Mice. Diabetes 2010, 59, 2540–2548. [Google Scholar] [CrossRef]
- Chhabra, K.H.; Chodavarapu, H.; Lazartigues, E. Angiotensin converting enzyme 2: A new important player in the regulation of glycemia. IUBMB Life 2013, 65, 731–738. [Google Scholar] [CrossRef]
- Bosso, M.; Thanaraj, T.A.; Abu-Farha, M.; Alanbaei, M.; Abubaker, J.; Al-Mulla, F. The Two Faces of ACE2: The Role of ACE2 Receptor and Its Polymorphisms in Hypertension and COVID-19. Mol. Ther. Methods Clin. Dev. 2020, 18, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Jaehwan, Y.; Seok, J.H.; Joo, M.; Bae, J.-Y.; Kim, J.I.; Park, M.-S.; Kim, K. Multifactorial Traits of SARS-CoV-2 Cell Entry Related to Diverse Host Proteases and Proteins. Biomol. Ther. 2021, 29, 249–262. [Google Scholar]
- Hatmal, M.M.; Alshaer, W.; Al-Hatamleh, M.A.I.; Hatmal, M.; Smadi, O.; Taha, M.O.; Oweida, A.J.; Boer, J.C.; Mohamud, R.; Plebanski, M. Comprehensive Structural and Molecular Comparison of Spike Proteins of SARS-CoV-2, Sars-Cov and Mers-Cov, and Their Interactions with Ace2. Cells 2020, 9, 2638. [Google Scholar] [CrossRef] [PubMed]
- Markus, H.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on Ace2 and Tmprss2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar]
- Patrick, B.; Refae, S.; Mograbi, B.; Hofman, P.; Milano, G. Host Polymorphisms May Impact SARS-CoV-2 Infectivity. Trends Genet. 2020, 36, 813–815. [Google Scholar]
- Heurich, A.; Hofmann-Winkler, H.; Gierer, S.; Liepold, T.; Jahn, O.; Pohlmann, S. Tmprss2 and Adam17 Cleave Ace2 Differentially and Only Proteolysis by Tmprss2 Augments Entry Driven by the Severe Acute Respiratory Syndrome Corona-virus Spike Protein. J. Virol. 2014, 88, 1293–1307. [Google Scholar] [CrossRef]
- Tipnis, S.R.; Hooper, N.M.; Hyde, R.; Karran, E.; Christie, G.; Turner, A.J. A Human Homolog of Angiotensin-converting Enzyme. J. Biol. Chem. 2000, 275, 33238–33243. [Google Scholar] [CrossRef]
- Turner, A.J.; Nalivaeva, N.N. Angiotensin-converting enzyme 2 (ACE2): Two decades of revelations and re-evaluation. Peptides 2022, 151, 170766. [Google Scholar] [CrossRef]
- Donoghue, M.; Hsieh, F.; Baronas, E.; Godbout, K.; Gosselin, M.; Stagliano, N.; Donovan, M.; Woolf, B.; Robison, K.; Jeyaseelan, R.; et al. A Novel Angiotensin-Converting Enzyme–Related Carboxypeptidase (ACE2) Converts Angiotensin I to Angiotensin 1-9. Circ. Res. 2000, 87, e1–e9. [Google Scholar] [CrossRef]
- García-Escobar, A.; Jiménez-Valero, S.; Galeote, G.; Jurado-Román, A.; García-Rodríguez, J.; Moreno, R. The soluble catalytic ectodomain of ACE2 a biomarker of cardiac remodelling: New insights for heart failure and COVID19. Hear. Fail. Rev. 2021, 26, 961–971. [Google Scholar] [CrossRef]
- Bakhshandeh, B.; Sorboni, S.G.; Javanmard, A.-R.; Mottaghi, S.S.; Mehrabi, M.-R.; Sorouri, F.; Abbasi, A.; Jahanafrooz, Z. Variants in ACE2; potential influences on virus infection and COVID-19 severity. Infect. Genet. Evol. 2021, 90, 104773. [Google Scholar] [CrossRef]
- Chen, F.; Zhang, Y.; Li, X.; Li, W.; Liu, X.; Xue, X. The Impact of Ace2 Polymorphisms on COVID-19 Disease: Susceptibility, Severity, and Therapy. Front. Cell Infect. Microbiol. 2021, 11, 753721. [Google Scholar] [CrossRef] [PubMed]
- Daniel, W.; Yarski, M.; Warner, F.J.; Thornhill, P.; Parkin, E.T.; Smith, A.I.; Hooper, N.M.; Turner, A.J. Tumor Necrosis Factor-A Convertase (Adam17) Mediates Regulated Ectodomain Shedding of the Se-vere-Acute Respiratory Syndrome-Coronavirus (Sars-Cov) Receptor, Angiotensin-Converting Enzyme-2 (Ace2). J. Biol. Chem. 2005, 280, 30113–30119. [Google Scholar]
- Jia, H.P.; Look, D.C.; Tan, P.; Shi, L.; Hickey, M.; Gakhar, L.; Chappell, M.C.; Wohlford-Lenane, C.; McCray, P.B., Jr. Ectodomain shedding of angiotensin converting enzyme 2 in human airway epithelia. Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 297, L84–L96. [Google Scholar] [CrossRef] [PubMed]
- Monika, G. Adam-17: The Enzyme That Does It All. Crit. Rev. Biochem. Mol. Biol. 2010, 45, 146–169. [Google Scholar]
- Haga, S.; Yamamoto, N.; Nakai-Murakami, C.; Osawa, Y.; Tokunaga, K.; Sata, T.; Yamamoto, N.; Sasazuki, T.; Ishizaka, Y. Modulation of Tnf- -Converting Enzyme by the Spike Protein of Sars-Cov and Ace2 Induces Tnf- Production and Facilitates Viral Entry. Proc. Natl. Acad. Sci. USA 2008, 105, 7809–7814. [Google Scholar] [CrossRef]
- Lobelia, S.; Uhal, B.D. Ace2, Much More Than Just a Receptor for SARS-CoV-2. Front. Cell. Infect. Microbiol. 2020, 10, 317. [Google Scholar]
- Zipeto, D.; Palmeira, J.D.F.; Arganaraz, G.A.; Arganaraz, E.R. Ace2/Adam17/Tmprss2 Interplay May Be the Main Risk Factor for COVID-19. Front. Immunol. 2020, 11, 576745. [Google Scholar] [CrossRef]
- Rahman, M.M.; Hasan, M.; Ahmed, A. Potential detrimental role of soluble ACE2 in severe COVID-19 comorbid patients. Rev. Med. Virol. 2021, 31, 1–12. [Google Scholar] [CrossRef]
- Chamsi-Pasha, M.A.R.; Shao, Z.; Tang, W.H.W. Angiotensin-Converting Enzyme 2 as a Therapeutic Target for Heart Failure. Curr. Hear. Fail. Rep. 2013, 11, 58–63. [Google Scholar] [CrossRef]
- Shahnawaz, M.; Nasrin, N.; Alotaibi, F.O.; Prasad, G.; Singh, S.K.; Alam, I.; Mustafa, G. Treatment Options Available for COVID-19 and an Analysis on Possible Role of Combination of Rhace2, Angioten-sin (1-7) and Angiotensin (1-9) as Effective Therapeutic Measure. SN Compr. Clin. Med. 2020, 2, 1761–1766. [Google Scholar]
- Yeung, M.L.; Teng, J.L.L.; Jia, L.; Zhang, C.; Huang, C.; Cai, J.P.; Zhou, R.; Chan, K.H.; Zhao, H.; Zhu, L.; et al. Soluble Ace2-Mediated Cell Entry of SARS-CoV-2 Via Interaction with Proteins Related to the Renin-Angiotensin System. Cell 2021, 184, 2212–2228 e12. [Google Scholar] [CrossRef] [PubMed]
- WHO. Coronavirus Disease (COVID-19) Situations Reports; WHO: Geneva, Switzerland, 2021. [Google Scholar]
- Guo, X.; Chen, Z.; Xia, Y.; Lin, W.; Li, H. Investigation of the Genetic Variation in Ace2 on the Structural Recognition by the Novel Coronavirus (SARS-CoV-2). J. Transl. Med. 2020, 18, 321. [Google Scholar] [CrossRef] [PubMed]
- Yi, L.; Liu, C.; Guan, T.; Li, Y.; Lai, Y.; Li, F.; Zhao, H.; Maimaiti, T.; Zeyaweiding, A. Association of Ace2 Genetic Polymorphisms with Hypertension-Related Target Organ Damages in South Xin-jiang. Hypertens. Res. 2019, 42, 681–689. [Google Scholar]
- Aneta, A.; Gagno, G.; Sinagra, G.; Beltrami, A.P.; Janjusevic, M.; Ippolito, G.; Zumla, A.; Fluca, A.L.; Ferro, F. Effects of SARS-CoV-2 on Cardiovascular System: The Dual Role of Angiotensin-Converting Enzyme 2 (Ace2) as the Virus Receptor and Homeostasis Regulator-Review. Int. J. Mol. Sci. 2021, 22, 4526. [Google Scholar]
- Antony, P.; Vijayan, R. Role of SARS-CoV-2 and Ace2 Variations in COVID-19. Biomed. J. 2021, 44, 235–244. [Google Scholar] [CrossRef]
- Suryamohan, K.; Diwanji, D.; Stawiski, E.W.; Gupta, R.; Miersch, S.; Liu, J.; Chen, C.; Jiang, Y.P.; Fellouse, F.A.; Sathirapongsasuti, J.F.; et al. Human Ace2 Receptor Polymorphisms and Altered Susceptibility to SARS-CoV-2. Commun. Biol. 2021, 4, 475. [Google Scholar] [CrossRef]
- Macgowan, S.A.; Barton, M.I.; Kutuzov, M.; Dushek, O.; van der Merwe, P.A.; Barton, G.J. Missense Variants in Human Ace2 Strongly Affect Binding to SARS-CoV-2 Spike Providing a Mechanism for Ace2 Mediated Genetic Risk in COVID-19: A Case Study in Affinity Predictions of Interface Variants. PLoS Comput. Biol. 2022, 18, e1009922. [Google Scholar] [CrossRef]
- Mushtaq, H.; Jabeen, N.; Raza, F.; Shabbir, S.; Baig, A.A.; Amanullah, A.; Aziz, B. Structural Variations in Human Ace2 May Influence Its Binding with SARS-CoV-2 Spike Protein. J. Med. Virol. 2020, 92, 1580–1586. [Google Scholar]
- Teng, S.; Tang, Q. ACE2 enhance viral infection or viral infection aggravate the underlying diseases. Comput. Struct. Biotechnol. J. 2020, 18, 2100–2106. [Google Scholar] [CrossRef]
- Marčetić, D.; Samaržija, M.; Dugac, A.V.; Knežević, J. Angiotensin-Converting Enzyme 2 (ACE2) as a Potential Diagnostic and Prognostic Biomarker for Chronic Inflammatory Lung Diseases. Genes 2021, 12, 1054. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Troyano, N.; Gabriel-Medina, P.; Weber, S.; Klammer, M.; Barquin-DelPino, R.; Castillo-Ribelles, L.; Esteban, A.; Hernandez-Gonzalez, M.; Ferrer-Costa, R.; Pumarola, T.; et al. Soluble Angiotensin-Converting Enzyme 2 as a Prognostic Biomarker for Disease Progression in Patients Infected with SARS-CoV-2. Diagnostics 2022, 12, 886. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| dbSNP ID | Substitution | Description | References | |
|---|---|---|---|---|
| AFR | rs73635825 | S19P |
| [39] |
| [40] | ||||
| AMR | rs781255386 | T27A |
| [37,41] |
| rs924799658 | F40L |
| [39] | |
| NFE | rs778030746 | I21V |
| [37] |
| rs756231991 | D23K |
| [37] | |
| ALL Al | rs1244687367 | I21T |
| [37] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sodhi, P.V.; Sidime, F.; Tarazona, D.D.; Valdivia, F.; Levano, K.S. A Closer Look at ACE2 Signaling Pathway and Processing during COVID-19 Infection: Identifying Possible Targets. Vaccines 2023, 11, 13. https://doi.org/10.3390/vaccines11010013
Sodhi PV, Sidime F, Tarazona DD, Valdivia F, Levano KS. A Closer Look at ACE2 Signaling Pathway and Processing during COVID-19 Infection: Identifying Possible Targets. Vaccines. 2023; 11(1):13. https://doi.org/10.3390/vaccines11010013
Chicago/Turabian StyleSodhi, Pia V., Francoise Sidime, David D. Tarazona, Faviola Valdivia, and Kelly S. Levano. 2023. "A Closer Look at ACE2 Signaling Pathway and Processing during COVID-19 Infection: Identifying Possible Targets" Vaccines 11, no. 1: 13. https://doi.org/10.3390/vaccines11010013
APA StyleSodhi, P. V., Sidime, F., Tarazona, D. D., Valdivia, F., & Levano, K. S. (2023). A Closer Look at ACE2 Signaling Pathway and Processing during COVID-19 Infection: Identifying Possible Targets. Vaccines, 11(1), 13. https://doi.org/10.3390/vaccines11010013