
Protection against Marburg Virus and Sudan Virus in NHP by an Adenovector-Based Trivalent Vaccine Regimen Is Correlated to Humoral Immune Response Levels


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viral Vaccines
2.2. Ethics Statement
2.3. Filovirus Challenge Material and Animal Challenge
2.4. Filovirus Glycoprotein Enzyme-Linked Immunosorbent Assay (ELISA) and Filovirus GP–Reactive Interferon Gamma (IFN-γ) Producing T-Cell ELISpot
2.5. Virus Neutralization Assays
2.6. Statistical Methods
3. Results
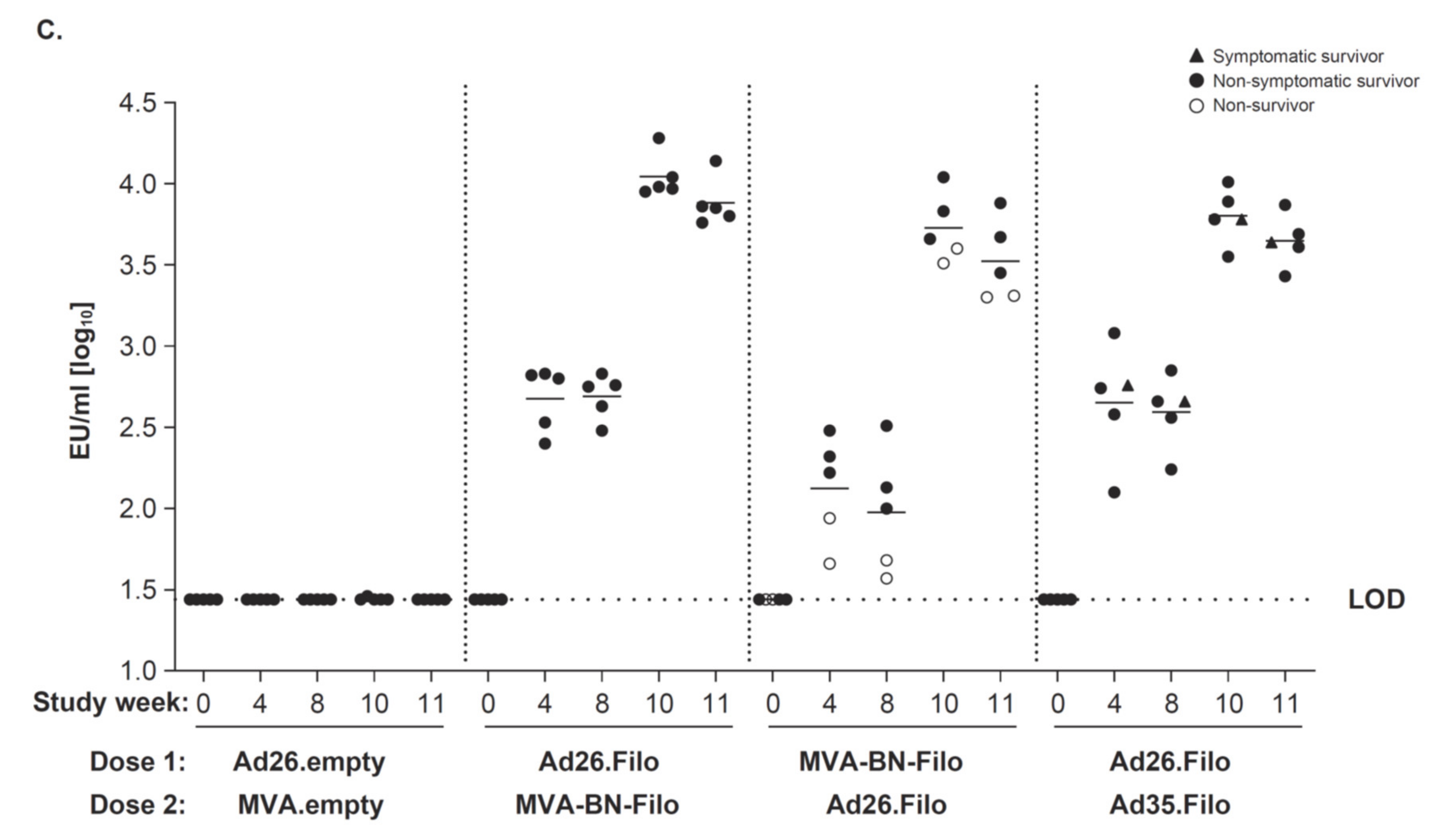
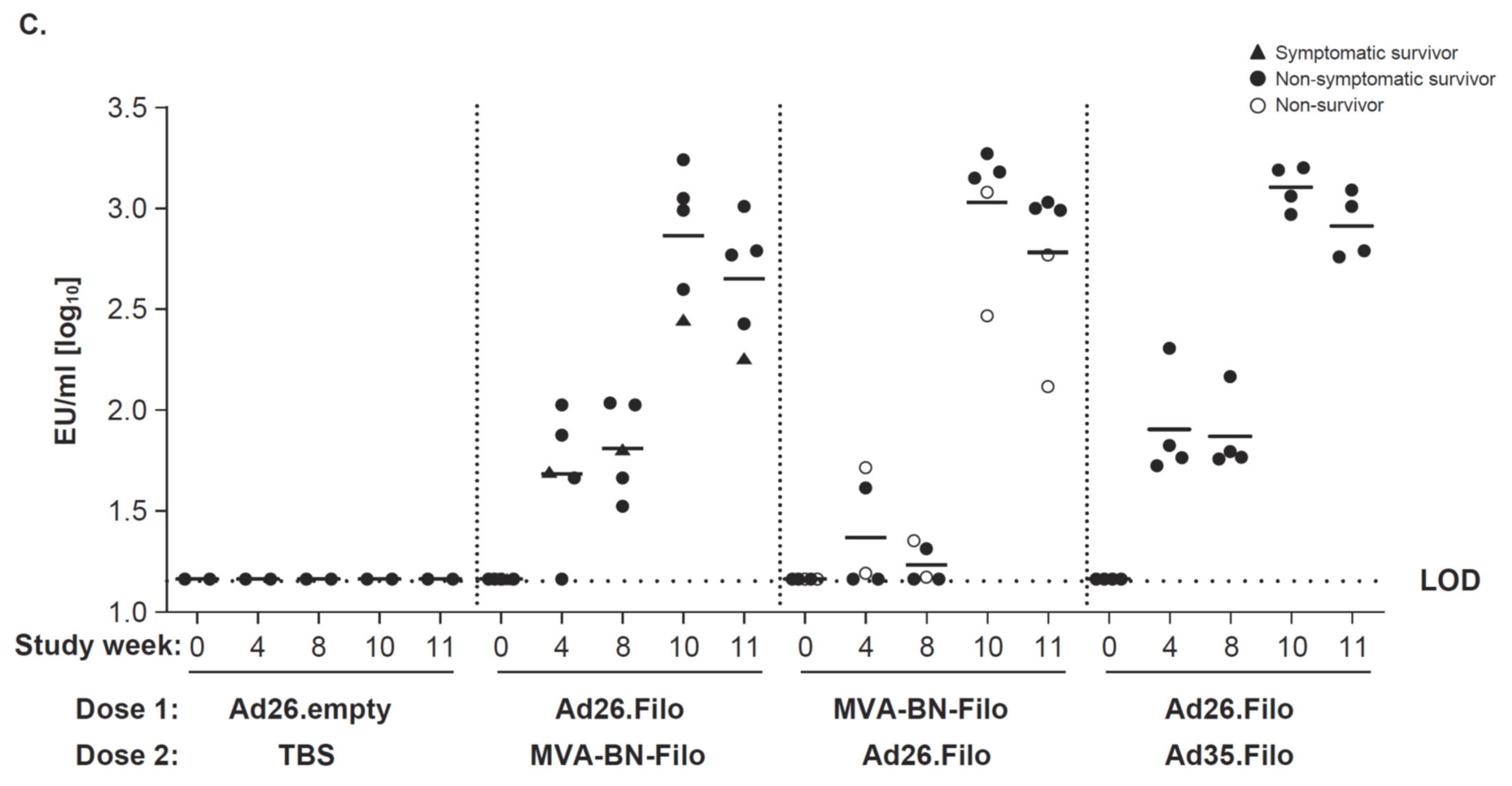
3.1. Immunogenicity and Protective Efficacy against SUDV Challenge
3.2. Immunogenicity and Protective Efficacy against MARV Challenge
3.3. Marburg and Sudan GP–Specific Antibody Responses Correlate with Protection
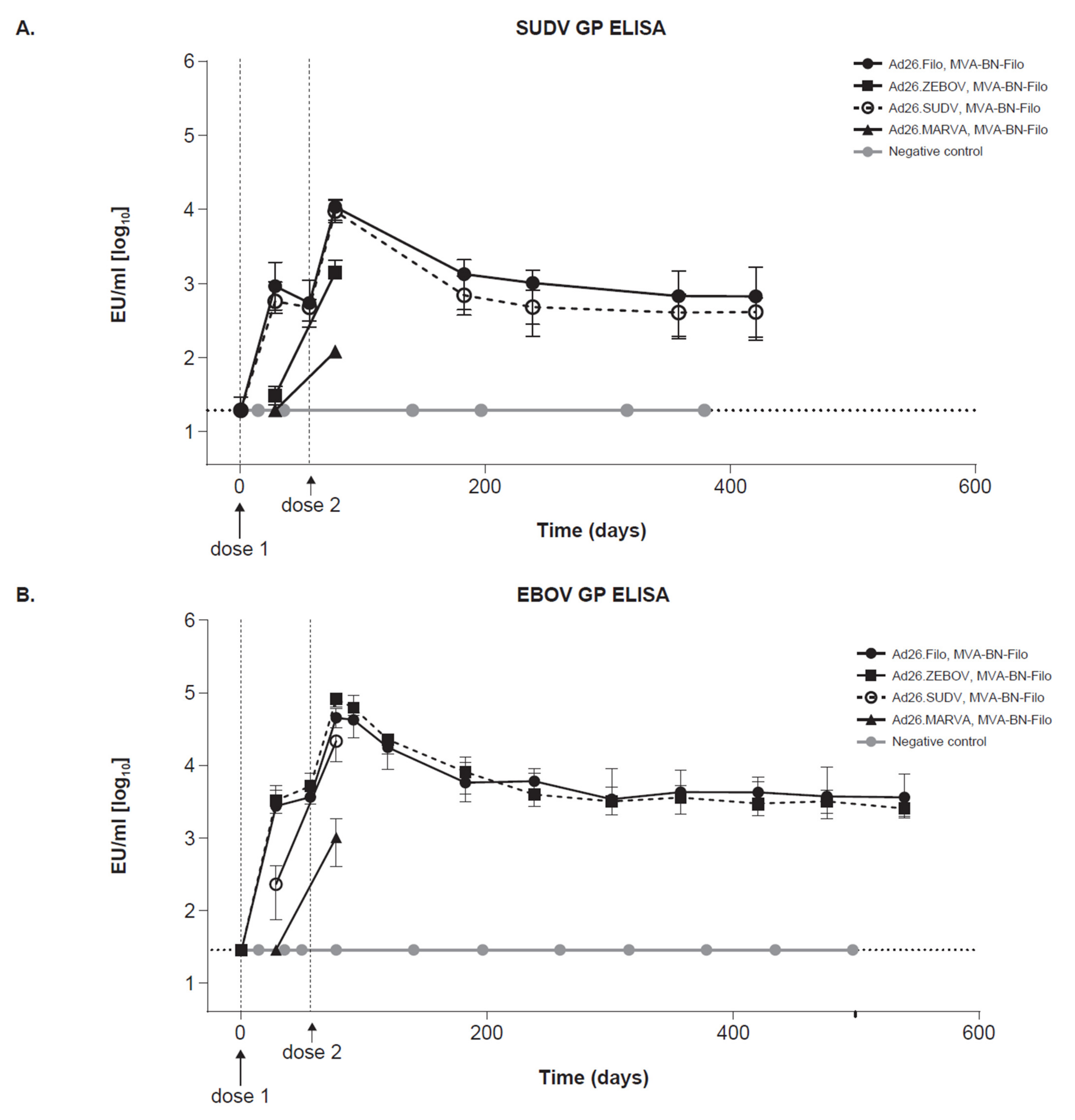
3.4. Protective Vaccine Regimens Induce Persistent Immune Responses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. West Africa’s First-Ever Case of Marburg Virus Disease Confirmed in Guinea. Available online: https://www.afro.who.int/news/west-africas-first-ever-case-marburg-virus-disease-confirmed-guinea (accessed on 5 May 2022).
- Jacob, S.T.; Crozier, I.; Fischer, W.A., 2nd; Hewlett, A.; Kraft, C.S.; Vega, M.A.; Soka, M.J.; Wahl, V.; Griffiths, A.; Bollinger, L.; et al. Ebola virus disease. Nat. Rev. Dis. Primers 2020, 6, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maganga, G.D.; Kapetshi, J.; Berthet, N.; Kebela Ilunga, B.; Kabange, F.; Mbala Kingebeni, P.; Mondonge, V.; Muyembe, J.J.; Bertherat, E.; Briand, S.; et al. Ebola virus disease in the Democratic Republic of Congo. N. Engl. J. Med. 2014, 371, 2083–2091. [Google Scholar] [CrossRef] [PubMed]
- Keita, A.K.; Koundouno, F.R.; Faye, M.; Dux, A.; Hinzmann, J.; Diallo, H.; Ayouba, A.; Le Marcis, F.; Soropogui, B.; Ifono, K.; et al. Resurgence of Ebola virus in 2021 in Guinea suggests a new paradigm for outbreaks. Nature 2021, 597, 539–543. [Google Scholar] [CrossRef]
- Chughtai, A.A.; Barnes, M.; Macintyre, C.R. Persistence of Ebola virus in various body fluids during convalescence: Evidence and implications for disease transmission and control. Epidemiol. Infect. 2016, 144, 1652–1660. [Google Scholar] [CrossRef] [Green Version]
- Den Boon, S.; Marsto, N.B.J.; Nyenswah, T.G.; Jambai, A.; Barry, M.; Keita, S.; Durski, K.; Senesie, S.S.; Perkins, D.; Shah, A.; et al. Ebola virus infection associated with transmission from survivors. Emerg. Infect. Dis. 2019, 25, 249–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martini, G.A.; Knauff, H.G.; Schmidt, H.A.; Mayer, G.; Baltzer, G. A hitherto unknown infectious disease contracted from monkeys. “Marburg-virus” disease. Ger. Med. Mon. 1968, 13, 457–470. [Google Scholar]
- Shoemaker, T.; MacNeil, A.; Balinandi, S.; Campbell, S.; Wamala, J.F.; McMullan, L.K.; Downing, R.; Lutwama, J.; Mbidde, E.; Stroher, U.; et al. Reemerging Sudan Ebola virus disease in Uganda, 2011. Emerg. Infect. Dis. 2012, 18, 1480–1483. [Google Scholar] [CrossRef]
- Baize, S.; Pannetier, D.; Oestereich, L.; Rieger, T.; Koivogui, L.; Magassouba, N.; Soropogui, B.; Sow, M.S.; Keita, S.; De Clerck, H.; et al. Emergence of Zaire Ebola virus disease in Guinea. N. Engl. J. Med. 2014, 371, 1418–1425. [Google Scholar] [CrossRef] [Green Version]
- Holmes, E.C.; Dudas, G.; Rambaut, A.; Andersen, K.G. The evolution of Ebola virus: Insights from the 2013–2016 epidemic. Nature 2016, 538, 193–200. [Google Scholar] [CrossRef] [Green Version]
- Henao-Restrepo, A.M.; Longini, I.M.; Egger, M.; Dean, N.E.; Edmunds, W.J.; Camacho, A.; Carroll, M.W.; Doumbia, M.; Draguez, B.; Duraffour, S.; et al. Efficacy and effectiveness of an rVSV-vectored vaccine expressing Ebola surface glycoprotein: Interim results from the Guinea ring vaccination cluster-randomised trial. Lancet 2015, 386, 857–866. [Google Scholar] [CrossRef]
- Henao-Restrepo, A.M.; Camacho, A.; Longini, I.M.; Watson, C.H.; Edmunds, W.J.; Egger, M.; Carroll, M.W.; Dean, N.E.; Diatta, I.; Doumbia, M.; et al. Efficacy and effectiveness of an rVSV-vectored vaccine in preventing Ebola virus disease: Final results from the Guinea ring vaccination, open-label, cluster-randomised trial (Ebola Ca Suffit!). Lancet 2017, 389, 505–518. [Google Scholar] [CrossRef] [Green Version]
- Callendret, B.; Vellinga, J.; Wunderlich, K.; Rodriguez, A.; Steigerwald, R.; Dirmeier, U.; Cheminay, C.; Volkmann, A.; Brasel, T.; Carrion, R.; et al. A prophylactic multivalent vaccine against different filovirus species is immunogenic and provides protection from lethal infections with Ebolavirus and Marburgvirus species in non-human primates. PLoS ONE 2018, 13, e0192312. [Google Scholar] [CrossRef] [Green Version]
- Roozendaal, R.; Hendriks, J.; van Effelterre, T.; Spiessens, B.; Dekking, L.; Solforosi, L.; Czapska-Casey, D.; Bockstal, V.; Stoop, J.; Splinter, D.; et al. Nonhuman primate to human immunobridging to infer the protective effect of an Ebola virus vaccine candidate. NPJ Vaccines 2020, 5, 112. [Google Scholar] [CrossRef]
- Pollard, A.J.; Launay, O.; Lelievre, J.D.; Lacabaratz, C.; Grande, S.; Goldstein, N.; Robinson, C.; Gaddah, A.; Bockstal, V.; Wiedemann, A.; et al. Safety and immunogenicity of a two-dose heterologous Ad26.ZEBOV and MVA-BN-Filo Ebola vaccine regimen in adults in Europe (EBOVAC2): A randomised, observer-blind, participant-blind, placebo-controlled, phase 2 trial. Lancet Infect. Dis. 2021, 21, 493–506. [Google Scholar] [CrossRef]
- Ishola, D.; Manno, D.; Afolabi, M.O.; Keshinro, B.; Bockstal, V.; Rogers, B.; Owusu-Kyei, K.; Serry-Bangura, A.; Swaray, I.; Lowe, B.; et al. Safety and long-term immunogenicity of the two-dose heterologous Ad26.ZEBOV and MVA-BN-Filo Ebola vaccine regimen in adults in Sierra Leone: A combined open-label, non-randomised stage 1, and a randomised, double-blind, controlled stage 2 trial. Lancet Infect. Dis. 2022, 22, 97–109. [Google Scholar] [CrossRef]
- Bockstal, V.; Leyssen, M.; Heerwegh, D.; Spiessens, B.; Robinson, C.; Stoop, J.N.; Roozendaal, R.; Van Effelterre, T.; Gaddah, A.; Van Roey, G.A.; et al. Nonhuman primate to human immunobridging demonstrates a protective effect of Ad26.ZEBOV, MVA-BN-Filo vaccine against Ebola virus disease. NPJ Vaccines 2022. submitted. [Google Scholar]
- Zahn, R.; Gillisen, G.; Roos, A.; Koning, M.; van der Helm, E.; Spek, D.; Weijtens, M.; Grazia Pau, M.; Radosevic, K.; Weverling, G.J.; et al. Ad35 and Ad26 vaccine vectors induce potent and cross-reactive antibody and T-cell responses to multiple filovirus species. PLoS ONE 2012, 7, e44115. [Google Scholar] [CrossRef]
- Towner, J.S.; Khristova, M.L.; Sealy, T.K.; Vincent, M.J.; Erickson, B.R.; Bawiec, D.A.; Hartman, A.L.; Comer, J.A.; Zaki, S.R.; Stroher, U.; et al. Marburgvirus genomics and association with a large hemorrhagic fever outbreak in Angola. J. Virol. 2006, 80, 6497–6516. [Google Scholar] [CrossRef] [Green Version]
- Alfson, K.J.; Avena, L.E.; Beadles, M.W.; Menzie, H.; Patterson, J.L.; Carrion, R., Jr.; Griffiths, A. Genetic changes at the glycoprotein editing site associated with serial passage of Sudan virus. J. Infect. Dis. 2015, 212 (Suppl. S2), S295–S304. [Google Scholar] [CrossRef] [Green Version]
- Rudge, T.L., Jr.; Sankovich, K.A.; Niemuth, N.A.; Anderson, M.S.; Badorrek, C.S.; Skomrock, N.D.; Cirimotich, C.M.; Sabourin, C.L. Development, qualification, and validation of the Filovirus Animal Nonclinical Group anti-Ebola virus glycoprotein immunoglobulin G enzyme-linked immunosorbent assay for human serum samples. PLoS ONE 2019, 14, e0215457. [Google Scholar] [CrossRef]
- Geisbert, T.W.; Bailey, M.; Geisbert, J.B.; Asiedu, C.; Roederer, M.; Grazia-Pau, M.; Custers, J.; Jahrling, P.; Goudsmit, J.; Koup, R.; et al. Vector choice determines immunogenicity and potency of genetic vaccines against Angola Marburg virus in nonhuman primates. J. Virol. 2010, 84, 10386–10394. [Google Scholar] [CrossRef] [Green Version]
- R Foundation for Statistical Computing. R: A Language and Environment for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 22 April 2022).
- Marzi, A.; Menicucci, A.R.; Engelmann, F.; Callison, J.; Horne, E.J.; Feldmann, F.; Jankeel, A.; Feldmann, H.; Messaoudi, I. Protection against Marburg virus using a recombinant VSV-vaccine depends on T and B cell activation. Front. Immunol. 2018, 9, 3071. [Google Scholar] [CrossRef] [Green Version]
- Marzi, A.; Jankeel, A.; Menicucci, A.R.; Callison, J.; O’Donnell, K.L.; Feldmann, F.; Pinski, A.N.; Hanley, P.W.; Messaoudi, I. Single dose of a VSV-based vaccine rapidly protects macaques from Marburg virus disease. Front. Immunol. 2021, 12, 774026. [Google Scholar] [CrossRef]
- Hunegnaw, R.; Honko, A.; Wang, L.; Carr, D.; Murray, T.; Shi, W.; Dulan, C.N.M.; Foulds, K.E.; Agans, K.N.; Cross, R.W.; et al. Rapid and durable protection against Marburg virus with a single-shot ChAd3-MARV GP vaccine. bioRxiv 2021. preprint. [Google Scholar] [CrossRef]
- Geisbert, T.W.; Bailey, M.; Hensley, L.; Asiedu, C.; Geisbert, J.; Stanley, D.; Honko, A.; Johnson, J.; Mulangu, S.; Pau, M.G.; et al. Recombinant adenovirus serotype 26 (Ad26) and Ad35 vaccine vectors bypass immunity to Ad5 and protect nonhuman primates against ebolavirus challenge. J. Virol. 2011, 85, 4222–4233. [Google Scholar] [CrossRef] [Green Version]
- Jones, S.M.; Feldmann, H.; Stroher, U.; Geisbert, J.B.; Fernando, L.; Grolla, A.; Klenk, H.D.; Sullivan, N.J.; Volchkov, V.E.; Fritz, E.A.; et al. Live attenuated recombinant vaccine protects nonhuman primates against Ebola and Marburg viruses. Nat. Med. 2005, 11, 786–790. [Google Scholar] [CrossRef]
- Sullivan, N.J.; Sanchez, A.; Rollin, P.E.; Yang, Z.Y.; Nabel, G.J. Development of a preventive vaccine for Ebola virus infection in primates. Nature 2000, 408, 605–609. [Google Scholar] [CrossRef]
- Wang, D.; Raja, N.U.; Trubey, C.M.; Juompan, L.Y.; Luo, M.; Woraratanadharm, J.; Deitz, S.B.; Yu, H.; Swain, B.M.; Moore, K.M.; et al. Development of a cAdVax-based bivalent Ebola virus vaccine that induces immune responses against both the Sudan and Zaire species of Ebola virus. J. Virol. 2006, 80, 2738–2746. [Google Scholar] [CrossRef] [Green Version]
- Swenson, D.L.; Wang, D.; Luo, M.; Warfield, K.L.; Woraratanadharm, J.; Holman, D.H.; Dong, J.Y.; Pratt, W.D. Vaccine to confer to nonhuman primates complete protection against multistrain Ebola and Marburg virus infections. Clin. Vaccine Immunol. 2008, 15, 460–467. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | N | Day 0 Immunization | Day 56 Immunization | Challenge |
|---|---|---|---|---|
| A | 5 | Ad26.empty | MVA.empty | SUDV Gulu * |
| B | 5 | Ad26.Filo | MVA-BN-Filo | SUDV Gulu * |
| C | 5 | MVA-BN-Filo | Ad26.Filo | SUDV Gulu * |
| D | 5 | Ad26.Filo | Ad35.Filo | SUDV Gulu * |
| Group | N | Day 0 Immunization | Day 56 Immunization | Challenge |
|---|---|---|---|---|
| A | 2 | Ad26.empty | TBS | MARV Angola * |
| B | 5 | Ad26.Filo | MVA-BN-Filo | MARV Angola * |
| C | 5 | MVA-BN-Filo | Ad26.Filo | MARV Angola * |
| D | 4 | Ad26.Filo | Ad35.Filo | MARV Angola * |
| Group | N | Day 0 Immunization | Day 56 Immunization |
|---|---|---|---|
| 1 | 10 | Ad26.Filo | MVA-BN-Filo |
| 2 | 5 | Ad26.ZEBOV | MVA-BN-Filo |
| 3 | 5 | Ad26.SUDV | MVA-BN-Filo |
| 4 | 5 | Ad26.MARVA | MVA-BN-Filo |
| 5 | 5 | Saline | Saline |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tiemessen, M.M.; Solforosi, L.; Dekking, L.; Czapska-Casey, D.; Serroyen, J.; Sullivan, N.J.; Volkmann, A.; Pau, M.G.; Callendret, B.; Schuitemaker, H.; et al. Protection against Marburg Virus and Sudan Virus in NHP by an Adenovector-Based Trivalent Vaccine Regimen Is Correlated to Humoral Immune Response Levels. Vaccines 2022, 10, 1263. https://doi.org/10.3390/vaccines10081263
Tiemessen MM, Solforosi L, Dekking L, Czapska-Casey D, Serroyen J, Sullivan NJ, Volkmann A, Pau MG, Callendret B, Schuitemaker H, et al. Protection against Marburg Virus and Sudan Virus in NHP by an Adenovector-Based Trivalent Vaccine Regimen Is Correlated to Humoral Immune Response Levels. Vaccines. 2022; 10(8):1263. https://doi.org/10.3390/vaccines10081263
Chicago/Turabian StyleTiemessen, Machteld M., Laura Solforosi, Liesbeth Dekking, Dominika Czapska-Casey, Jan Serroyen, Nancy J. Sullivan, Ariane Volkmann, Maria Grazia Pau, Benoit Callendret, Hanneke Schuitemaker, and et al. 2022. "Protection against Marburg Virus and Sudan Virus in NHP by an Adenovector-Based Trivalent Vaccine Regimen Is Correlated to Humoral Immune Response Levels" Vaccines 10, no. 8: 1263. https://doi.org/10.3390/vaccines10081263
APA StyleTiemessen, M. M., Solforosi, L., Dekking, L., Czapska-Casey, D., Serroyen, J., Sullivan, N. J., Volkmann, A., Pau, M. G., Callendret, B., Schuitemaker, H., Luhn, K., Zahn, R., & Roozendaal, R. (2022). Protection against Marburg Virus and Sudan Virus in NHP by an Adenovector-Based Trivalent Vaccine Regimen Is Correlated to Humoral Immune Response Levels. Vaccines, 10(8), 1263. https://doi.org/10.3390/vaccines10081263