
Harnessing Immune Evasion Strategy of Lymphatic Filariae: A Therapeutic Approach against Inflammatory and Infective Pathology



Abstract
1. Introduction
2. Immunomodulation Strategies
2.1. Infective L3 Mute Cutaneous Innate Cells
2.2. Filariae Regulate PRR Signaling
2.3. HDPs Mimic Host Glycans and Immune Molecules
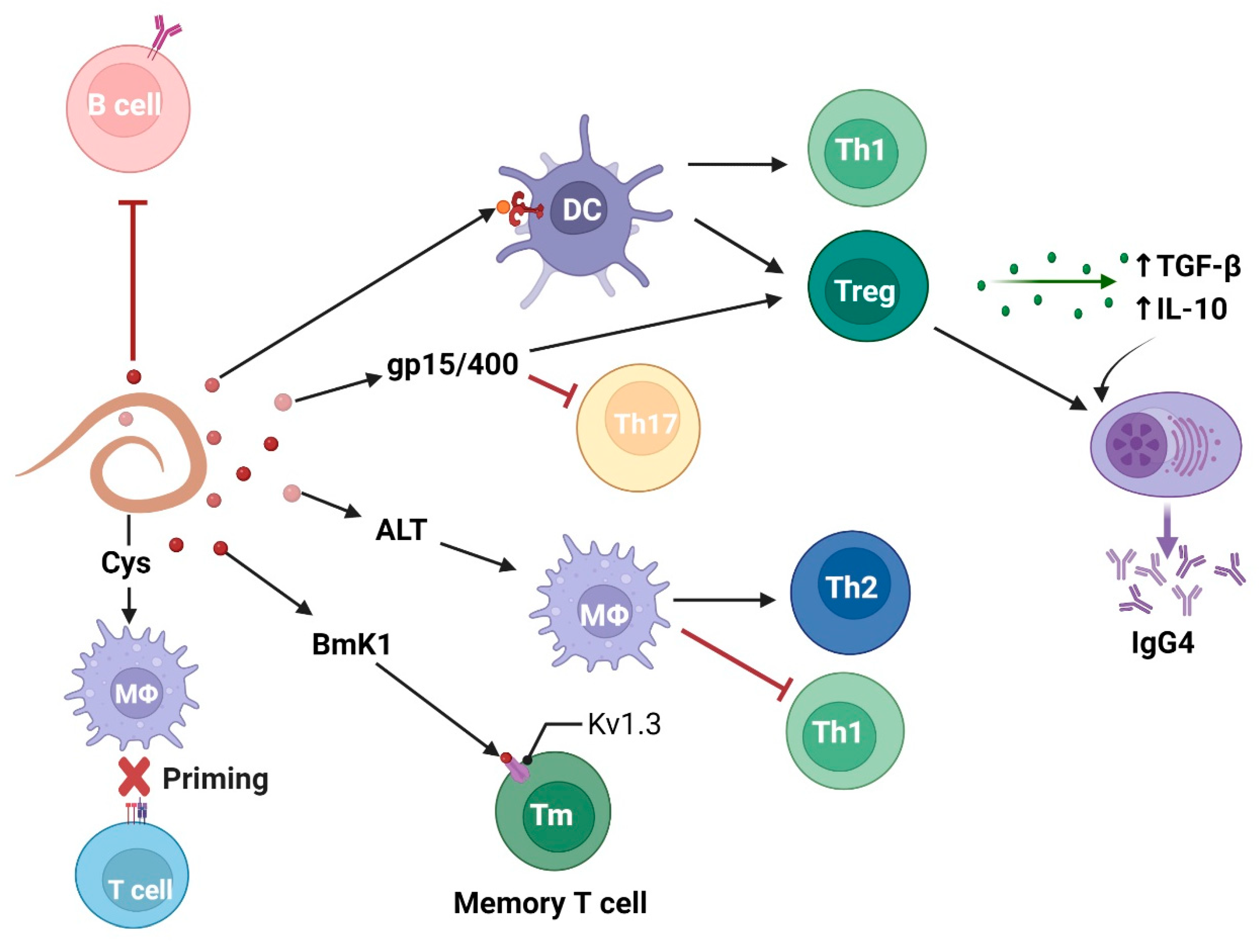
2.4. Filariae Manipulate B and T Cell Response
2.5. Filarial Parasites Induce Immune Cell Apoptosis and Autophagy
2.6. Non-Cellular Immune Evasion by Filarial Parasites
3. Filarial Immunomodulatory Strategy as a Treatment against Diseases
3.1. Lymphatic Filariasis
3.2. Malaria
3.3. Leishmaniasis
3.4. Inflammatory Diseases
4. Conclusions and Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moreno, Y.; Geary, T.G. Stage- and Gender-Specific Proteomic Analysis of Brugia malayi Excretory-Secretory Products. PLoS Negl. Trop. Dis. 2008, 2, e326. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.A.; Lizotte-Waniewski, M.R.; Foster, J.; Guiliano, D.; Daub, J.; Scott, A.L.; Slatko, B.; Blaxter, M.L. The Filarial Genome Project: Analysis of the Nuclear, Mitochondrial and Endosymbiont Genomes of Brugia malayi. Int. J. Parasitol. 2000, 30, 411–419. [Google Scholar] [CrossRef]
- Scott, A.L.; Ghedin, E. The Genome of Brugia malayi—All Worms Are Not Created Equal. Parasitol. Int. 2009, 58, 6–11. [Google Scholar] [CrossRef] [PubMed]
- International Helminth Genomes Consortium. Comparative genomics of the major parasitic worms. Nat. Genet. 2019, 51, 163–174. [Google Scholar] [CrossRef]
- Babu, S.; Nutman, T.B. Proinflammatory Cytokines Dominate the Early Immune Response to Filarial Parasites. J. Immunol. 2003, 171, 6723–6732. [Google Scholar] [CrossRef]
- Porthouse, K.H.; Chirgwin, S.R.; Coleman, S.U.; Taylor, H.W.; Klei, T.R. Inflammatory Responses to Migrating Brugia pahangi Third-Stage Larvae. Infect. Immun. 2006, 74, 2366–2372. [Google Scholar] [CrossRef]
- Allen, J.E.; Maizels, R.M. Diversity and Dialogue in Immunity to Helminths. Nat. Rev. Immunol. 2011, 11, 375–388. [Google Scholar] [CrossRef]
- Herbert, D.; Douglas, B.; Zullo, K. Group 2 Innate Lymphoid Cells (ILC2): Type 2 Immunity and Helminth Immunity. Int. J. Mol. Sci. 2019, 20, 2276. [Google Scholar] [CrossRef] [PubMed]
- Boyd, A.; Bennuru, S.; Wang, Y.; Sanprasert, V.; Law, M.; Chaussabel, D.; Nutman, T.B.; Semnani, R.T. Quiescent Innate Response to Infective Filariae by Human Langerhans Cells Suggests a Strategy of Immune Evasion. Infect. Immun. 2013, 81, 1420–1429. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Cotton, R.N.; McDonald-Fleming, R.; Boyd, A.; Spates, K.; Nutman, T.B.; Tolouei Semnani, R. Brugia malayi Infective Larvae Fail to Activate Langerhans Cells and Dermal Dendritic Cells in Human Skin. Parasite Immunol. 2015, 37, 79–91. [Google Scholar] [CrossRef]
- Bonne-Année, S.; Nutman, T.B. Human Innate Lymphoid Cells (ILCs) in Filarial Infections. Parasite Immunol. 2018, 40, e12442. [Google Scholar] [CrossRef] [PubMed]
- Boyd, A.; Ribeiro, J.M.C.; Nutman, T.B. Human CD117 (CKit)+ Innate Lymphoid Cells Have a Discrete Transcriptional Profile at Homeostasis and Are Expanded during Filarial Infection. PLoS ONE 2014, 9, e108649. [Google Scholar] [CrossRef]
- Metenou, S.; Dembele, B.; Konate, S.; Dolo, H.; Coulibaly, S.Y.; Coulibaly, Y.I.; Diallo, A.A.; Soumaoro, L.; Coulibaly, M.E.; Sanogo, D.; et al. At Homeostasis Filarial Infections Have Expanded Adaptive T Regulatory but Not Classical Th2 Cells. J. Immunol. 2010, 184, 5375–5382. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wu, M. Pattern Recognition Receptors in Health and Diseases. Signal Transduct. Target. Ther. 2021, 6, 291. [Google Scholar] [CrossRef] [PubMed]
- Tawill, S.; le Goff, L.; Ali, F.; Blaxter, M.; Allen, J.E. Both Free-Living and Parasitic Nematodes Induce a Characteristic Th2 Response That Is Dependent on the Presence of Intact Glycans. Infect. Immun. 2004, 72, 398–407. [Google Scholar] [CrossRef]
- van Die, I.; Cummings, R.D. Glycan Gimmickry by Parasitic Helminths: A Strategy for Modulating the Host Immune Response? Glycobiology 2010, 20, 2–12. [Google Scholar] [CrossRef]
- Forsyth, K.P.; Spark, R.; Kazura, J.; Brown, G.V.; Peters, P.; Heywood, P.; Dissanayake, S.; Mitchell, G.F. A Monoclonal Antibody-Based Immunoradiometric Assay for Detection of Circulating Antigen in Bancroftian Filariasis. J. Immunol. 1985, 134, 1172–1177. [Google Scholar]
- Maizels, R.M.; Burke, J.; Denham, D.A. Phosphorylcholine-Bearing Antigens in Filarial Nematode Parasites: Analysis of Somatic Extracts, in-Vitro Secretions and Infection Sera from Brugia malayi and B. pahangi. Parasite Immunol. 1987, 9, 49–66. [Google Scholar] [CrossRef]
- Harnett, W.; Deehan, M.R.; Houston, K.M.; Harnett, M.M. Immunomodulatory Properties of a Phosphorylcholine-Containing Secreted Filarial Glycoprotein. Parasite Immunol. 1999, 21, 601–608. [Google Scholar] [CrossRef]
- Bunte, M.J.M.; Schots, A.; Kammenga, J.E.; Wilbers, R.H.P. Helminth Glycans at the Host-Parasite Interface and Their Potential for Developing Novel Therapeutics. Front. Mol. Biosci. 2022, 8, 807821. [Google Scholar] [CrossRef]
- Melendez, A.J.; Harnett, M.M.; Pushparaj, P.N.; Wong, W.S.F.; Tay, H.K.; McSharry, C.P.; Harnett, W. Inhibition of Fc Epsilon RI-Mediated Mast Cell Responses by ES-62, a Product of Parasitic Filarial Nematodes. Nat. Med. 2007, 13, 1375–1381. [Google Scholar] [CrossRef]
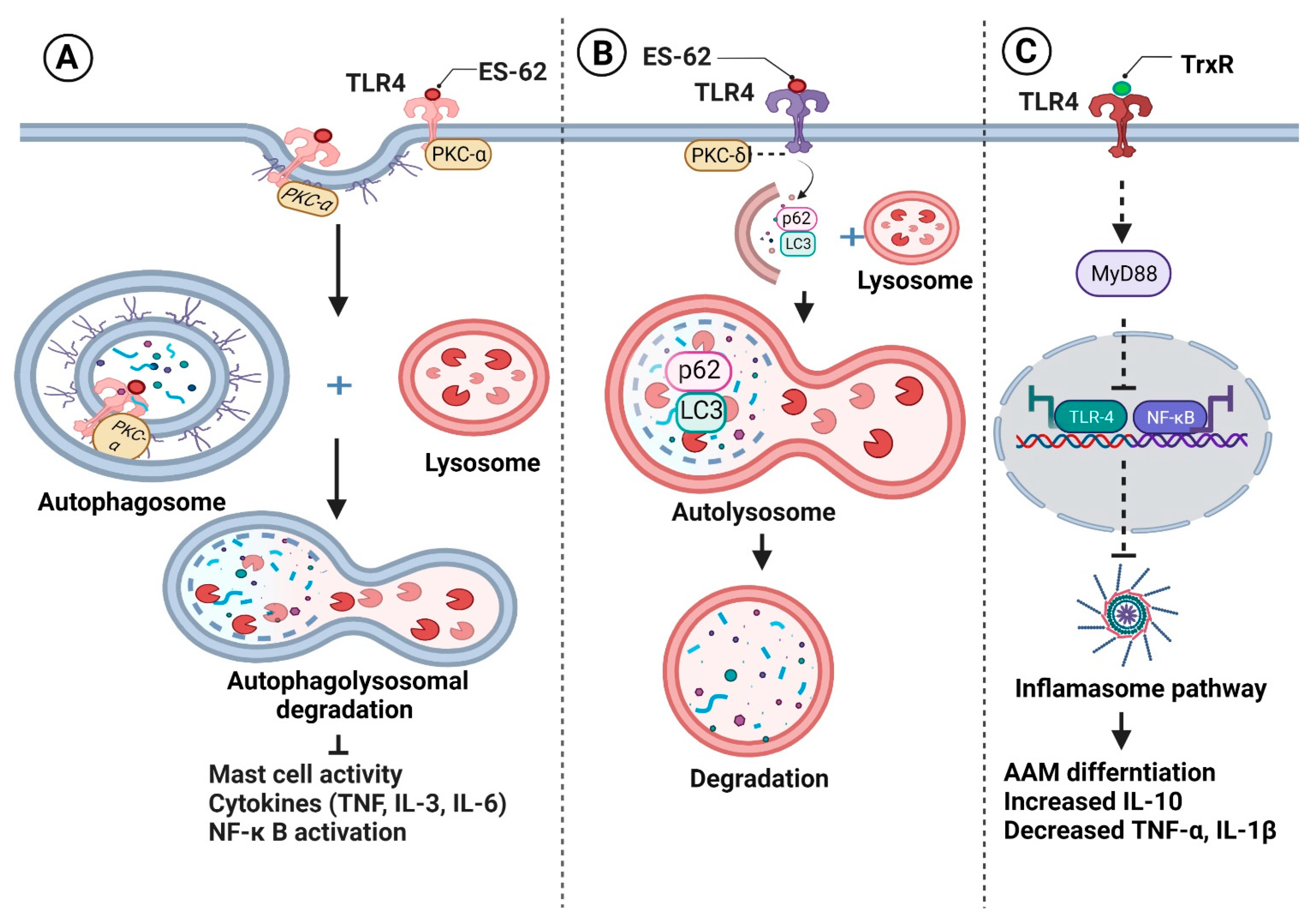
- Eason, R.J.; Bell, K.S.; Marshall, F.A.; Rodgers, D.T.; Pineda, M.A.; Steiger, C.N.; Al-Riyami, L.; Harnett, W.; Harnett, M.M. The Helminth Product, ES-62 Modulates Dendritic Cell Responses by Inducing the Selective Autophagolysosomal Degradation of TLR-Transducers, as Exemplified by PKCδ. Sci. Rep. 2016, 6, 37276. [Google Scholar] [CrossRef] [PubMed]
- Semnani, R.T.; Venugopal, P.G.; Leifer, C.A.; Mostböck, S.; Sabzevari, H.; Nutman, T.B. Inhibition of TLR3 and TLR4 Function and Expression in Human Dendritic Cells by Helminth Parasites. Blood 2008, 112, 1290–1298. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Karnam, A.; Das, M.; Babu, S.P.S.; Bayry, J. Wuchereria bancrofti Filaria Activates Human Dendritic Cells and Polarizes T Helper 1 and Regulatory T Cells via Toll-like Receptor 4. Commun. Biol. 2019, 2, 169. [Google Scholar] [CrossRef] [PubMed]
- Joardar, N.; Bhattacharya, R.; Halder, S.; Sen, A.; Biswas, S.R.; Jana, K.; Babu, S.P.S. Filarial Thioredoxin Reductase Exerts Anti-Inflammatory Effects upon Lipopolysaccharide Induced Inflammation in Macrophages. Int. J. Biol. Macromol. 2021, 193, 1379–1390. [Google Scholar] [CrossRef]
- Ludin, P.; Nilsson, D.; Mäser, P. Genome-Wide Identification of Molecular Mimicry Candidates in Parasites. PLoS ONE 2011, 6, e17546. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Escobar, N.; Gregory, W.F.; Maizels, R.M. Identification of Tgh-2, a Filarial Nematode Homolog of Caenorhabditis Elegans Daf-7 and Human Transforming Growth Factor β, Expressed in Microfilarial and Adult Stages of Brugia malayi. Infect. Immun. 2000, 68, 6402–6410. [Google Scholar] [CrossRef]
- Maizels, R.M.; Smits, H.H.; McSorley, H.J. Modulation of Host Immunity by Helminths: The Expanding Repertoire of Parasite Effector Molecules. Immunity 2018, 49, 801–818. [Google Scholar] [CrossRef]
- Dakshinamoorthy, G.; Samykutty, A.K.; Munirathinam, G.; Shinde, G.B.; Nutman, T.; Reddy, M.V.; Kalyanasundaram, R. Biochemical Characterization and Evaluation of a Brugia malayi Small Heat Shock Protein as a Vaccine against Lymphatic Filariasis. PLoS ONE 2012, 7, e34077. [Google Scholar] [CrossRef]
- Liu, L.X.; Weller, P.F. Intravascular Filarial Parasites Inhibit Platelet Aggregation. Role of Parasite-Derived Prostanoids. J. Clin. Investig. 1992, 89, 1113–1120. [Google Scholar] [CrossRef][Green Version]
- Kalinski, P. Regulation of Immune Responses by Prostaglandin E 2. J. Immunol. 2012, 188, 21–28. [Google Scholar] [CrossRef]
- Baratelli, F.; Lin, Y.; Zhu, L.; Yang, S.-C.; Heuzé-Vourc’h, N.; Zeng, G.; Reckamp, K.; Dohadwala, M.; Sharma, S.; Dubinett, S.M. Prostaglandin E 2 Induces FOXP3 Gene Expression and T Regulatory Cell Function in Human CD4+ T Cells. J. Immunol. 2005, 175, 1483–1490. [Google Scholar] [CrossRef] [PubMed]
- McSorley, H.J.; Harcus, Y.M.; Murray, J.; Taylor, M.D.; Maizels, R.M. Expansion of Foxp3+ Regulatory T Cells in Mice Infected with the Filarial Parasite Brugia malayi. J. Immunol. 2008, 181, 6456–6466. [Google Scholar] [CrossRef]
- Oyesola, O.O.; Tait Wojno, E.D. Prostaglandin Regulation of Type 2 Inflammation: From Basic Biology to Therapeutic Interventions. Eur. J. Immunol. 2021, 51, 2399–2416. [Google Scholar] [CrossRef]
- Zang, X.; Taylor, P.; Wang, J.M.; Meyer, D.J.; Scott, A.L.; Walkinshaw, M.D.; Maizels, R.M. Homologues of Human Macrophage Migration Inhibitory Factor from a Parasitic Nematode. J. Biol. Chem. 2002, 277, 44261–44267. [Google Scholar] [CrossRef]
- Prieto-Lafuente, L.; Gregory, W.F.; Allen, J.E.; Maizels, R.M. MIF Homologues from a Filarial Nematode Parasite Synergize with IL-4 to Induce Alternative Activation of Host Macrophages. J. Leukoc. Biol. 2009, 85, 844–854. [Google Scholar] [CrossRef]
- Koussa, J.; Vitrinel, B.; Whitney, P.; Kasper, B.T.; Mahal, L.K.; Vogel, C.; Lustigman, S.; Salehi-Ashtiani, K.; Ghedin, E. Sex-Specific Glycosylation of Secreted Immunomodulatory Proteins in the Filarial Nematode Brugia malayi. bioRxiv 2021. [Google Scholar] [CrossRef]
- Ritter, M.; Osei-Mensah, J.; Debrah, L.B.; Kwarteng, A.; Mubarik, Y.; Debrah, A.Y.; Pfarr, K.; Hoerauf, A.; Layland, L.E. Wuchereria Bancrofti-Infected Individuals Harbor Distinct IL-10-Producing Regulatory B and T Cell Subsets Which Are Affected by Anti-Filarial Treatment. PLoS Negl. Trop. Dis. 2019, 13, e0007436. [Google Scholar] [CrossRef]
- Gomez-Escobar, N.; Bennett, C.; Prieto-Lafuente, L.; Aebischer, T.; Blackburn, C.C.; Maizels, R.M. Heterologous Expression of the Filarial Nematode Alt Gene Products Reveals Their Potential to Inhibit Immune Function. BMC Biol. 2005, 3, 8. [Google Scholar] [CrossRef] [PubMed]
- Chhabra, S.; Chang, S.C.; Nguyen, H.M.; Huq, R.; Tanner, M.R.; Londono, L.M.; Estrada, R.; Dhawan, V.; Chauhan, S.; Upadhyay, S.K.; et al. Kv1.3 Channel-blocking Immunomodulatory Peptides from Parasitic Worms: Implications for Autoimmune Diseases. FASEB J. 2014, 28, 3952–3964. [Google Scholar] [CrossRef] [PubMed]
- Harnett, W.; Harnett, M.M. Inhibition of Murine B Cell Proliferation and Down-Regulation of Protein Kinase C Levels by a Phosphorylcholine-Containing Filarial Excretory-Secretory Product. J. Immunol. 1993, 151, 4829–4837. [Google Scholar]
- Hewitson, J.P.; Harcus, Y.M.; Curwen, R.S.; Dowle, A.A.; Atmadja, A.K.; Ashton, P.D.; Wilson, A.; Maizels, R.M. The Secretome of the Filarial Parasite, Brugia malayi: Proteomic Profile of Adult Excretory–Secretory Products. Mol. Biochem. Parasitol. 2008, 160, 8–21. [Google Scholar] [CrossRef]
- Adjobimey, T.; Hoerauf, A. Induction of Immunoglobulin G4 in Human Filariasis: An Indicator of Immunoregulation. Ann. Trop. Med. Parasitol. 2010, 104, 455–464. [Google Scholar] [CrossRef]
- Khatri, V.; Chauhan, N.; Kalyanasundaram, R. Parasite Cystatin: Immunomodulatory Molecule with Therapeutic Activity against Immune Mediated Disorders. Pathogens 2020, 9, 431. [Google Scholar] [CrossRef] [PubMed]
- Manoury, B.; Gregory, W.F.; Maizels, R.M.; Watts, C. Bm-CPI-2, a Cystatin Homolog Secreted by the Filarial Parasite Brugia malayi, Inhibits Class II MHC-Restricted Antigen Processing. Curr. Biol. 2001, 11, 447–451. [Google Scholar] [CrossRef]
- Hewitson, J.P.; Grainger, J.R.; Maizels, R.M. Helminth Immunoregulation: The Role of Parasite Secreted Proteins in Modulating Host Immunity. Mol. Biochem. Parasitol. 2009, 167, 1–11. [Google Scholar] [CrossRef]
- Mucida, D.; Park, Y.; Kim, G.; Turovskaya, O.; Scott, I.; Kronenberg, M.; Cheroutre, H. Reciprocal TH17 and Regulatory T Cell Differentiation Mediated by Retinoic Acid. Science 2007, 317, 256–260. [Google Scholar] [CrossRef]
- Babu, S.; Blauvelt, C.P.; Nutman, T.B. Filarial Parasites Induce NK Cell Activation, Type 1 and Type 2 Cytokine Secretion, and Subsequent Apoptotic Cell Death. J. Immunol. 2007, 179, 2445–2456. [Google Scholar] [CrossRef]
- Semnani, R.T.; Liu, A.Y.; Sabzevari, H.; Kubofcik, J.; Zhou, J.; Gilden, J.K.; Nutman, T.B. Brugia malayi Microfilariae Induce Cell Death in Human Dendritic Cells, Inhibit Their Ability to Make IL-12 and IL-10, and Reduce Their Capacity to Activate CD4 + T Cells. J. Immunol. 2003, 171, 1950–1960. [Google Scholar] [CrossRef]
- Narasimhan, P.B.; Bennuru, S.; Meng, Z.; Cotton, R.N.; Elliott, K.R.; Ganesan, S.; McDonald-Fleming, R.; Veenstra, T.D.; Nutman, T.B.; Tolouei Semnani, R. Microfilariae of Brugia malayi Inhibit the MTOR Pathway and Induce Autophagy in Human Dendritic Cells. Infect. Immun. 2016, 84, 2463–2472. [Google Scholar] [CrossRef]
- Ricciardi, A.; Bennuru, S.; Tariq, S.; Kaur, S.; Wu, W.; Elkahloun, A.G.; Arakelyan, A.; Shaik, J.; Dorward, D.W.; Nutman, T.B.; et al. Extracellular Vesicles Released from the Filarial Parasite Brugia malayi Downregulate the Host MTOR Pathway. PLoS Negl. Trop. Dis. 2021, 15, e0008884. [Google Scholar] [CrossRef]
- Mishra, R.; Panda, S.K.; Sahoo, P.K.; Bal, M.S.; Satapathy, A.K. Increased Fas Ligand Expression of Peripheral B-1 Cells Correlated with CD4 + T-Cell Apoptosis in Filarial-Infected Patients. Parasite Immunol. 2017, 39, e12421. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, I.; Eisinger, S.W.; Raghavan, N.; Scott, A.L. Thioredoxin Peroxidases from Brugia malayi. Mol. Biochem. Parasitol. 1998, 91, 207–220. [Google Scholar] [CrossRef]
- Gnanasekar, M.; Rao, K.V.N.; Chen, L.; Narayanan, R.B.; Geetha, M.; Scott, A.L.; Ramaswamy, K.; Kaliraj, P. Molecular Characterization of a Calcium Binding Translationally Controlled Tumor Protein Homologue from the Filarial Parasites Brugia malayi and Wuchereria bancrofti. Mol. Biochem. Parasitol. 2002, 121, 107–118. [Google Scholar] [CrossRef]
- Bhargavi, R.; Vishwakarma, S.; Murty, U.S. Modeling Analysis of GST (Glutathione-S-Transferases) from Wuchereria bancrofti and Brugia malayi. Bioinformation 2005, 1, 25–27. [Google Scholar] [CrossRef][Green Version]
- Yadav, S.; Gupta, S.; Selvaraj, C.; Doharey, P.K.; Verma, A.; Singh, S.K.; Saxena, J.K. In Silico and In Vitro Studies on the Protein-Protein Interactions between Brugia malayi Immunomodulatory Protein Calreticulin and Human C1q. PLoS ONE 2014, 9, e106413. [Google Scholar] [CrossRef]
- World Health Organization Lymphatic Filariasis. 2022. Lymphatic Filariasis. Available online: https://www.who.int/news-room/fact-sheets/detail/lymphatic-filariasis (accessed on 2 June 2022).
- Mallawarachchi, C.H.; Nilmini Chandrasena, T.G.A.; Premaratna, R.; Mallawarachchi, S.M.N.S.M.; de Silva, N.R. Human Infection with Sub-Periodic Brugia spp. in Gampaha District, Sri Lanka: A Threat to Filariasis Elimination Status? Parasites Vectors 2018, 11, 68. [Google Scholar] [CrossRef]
- George, S.; Joy, T.M.; Kumar, A.; Panicker, K.N.; George, L.S.; Raj, M.; Leelamoni, K.; Nair, P. Prevalence of Neglected Tropical Diseases (Leishmaniasis and Lymphatic Filariasis) and Malaria among a Migrant Labour Settlement in Kerala, India. J. Immigr. Minority Health 2019, 21, 563–569. [Google Scholar] [CrossRef]
- Zuchi, A.; Prust, L.T.; Rocha, A.; Araújo, J.; da Silva, P.S.; Fiorillo, K.; Brandão, E.; Ximenes, C.; Lopes, F.; Ponzi, C.C. Screening and Evaluation of Lymphatic Filariasis in Immigrants from Endemic Countries Residing in a Focus Where It Is Considered Eliminated in the Southern Region of Brazil: A Risk of Reemergence? Acta Trop. 2017, 176, 192–196. [Google Scholar] [CrossRef]
- Da Silva, E.F.; de Lacerda, M.V.G.; Fontes, G.; Mourão, M.P.G.; Martins, M. Wuchereria bancrofti Infection in Haitian Immigrants and the Risk of Re-Emergence of Lymphatic Filariasis in the Brazilian Amazon. Rev. Soc. Bras. Med. Trop. 2017, 50, 256–259. [Google Scholar] [CrossRef][Green Version]
- Xu, Z.; Lau, C.L.; Zhou, X.; Fuimaono, S.; Soares Magalhães, R.J.; Graves, P.M. The Extensive Networks of Frequent Population Mobility in the Samoan Islands and Their Implications for Infectious Disease Transmission. Sci. Rep. 2018, 8, 10136. [Google Scholar] [CrossRef]
- Prada, J.M.; Stolk, W.A.; Davis, E.L.; Touloupou, P.; Sharma, S.; Muñoz, J.; Caja Rivera, R.M.; Reimer, L.J.; Michael, E.; de Vlas, S.J.; et al. Delays in Lymphatic Filariasis Elimination Programmes Due to COVID-19, and Possible Mitigation Strategies. Trans. R. Soc. Trop. Med. Hyg. 2021, 115, 261–268. [Google Scholar] [CrossRef]
- Bizhani, N.; Hashemi Hafshejani, S.; Mohammadi, N.; Rezaei, M.; Rokni, M.B. Lymphatic Filariasis in Asia: A Systematic Review and Meta-Analysis. Parasitol. Res. 2021, 120, 411–422. [Google Scholar] [CrossRef]
- Hussain, M.A.; Sitha, A.K.; Swain, S.; Kadam, S.; Pati, S. Mass Drug Administration for Lymphatic Filariasis Elimination in a Coastal State of India: A Study on Barriers to Coverage and Compliance. Infect. Dis. Poverty 2014, 3, 31. [Google Scholar] [CrossRef] [PubMed]
- Ahorlu, C.S.K.; Koka, E.; Adu-Amankwah, S.; Otchere, J.; de Souza, D.K. Community Perspectives on Persistent Transmission of Lymphatic Filariasis in Three Hotspot Districts in Ghana after 15 Rounds of Mass Drug Administration: A Qualitative Assessment. BMC Public Health 2018, 18, 238. [Google Scholar] [CrossRef]
- Willis, G.A.; Mayfield, H.J.; Kearns, T.; Naseri, T.; Thomsen, R.; Gass, K.; Sheridan, S.; Graves, P.M.; Lau, C.L. A Community Survey of Coverage and Adverse Events Following Country-Wide Triple-Drug Mass Drug Administration for Lymphatic Filariasis Elimination, Samoa 2018. PLOS Negl. Trop. Dis. 2020, 14, e0008854. [Google Scholar] [CrossRef]
- de Souza, D.K.; Gass, K.; Otchere, J.; Htet, Y.M.; Asiedu, O.; Marfo, B.; Biritwum, N.-K.; Boakye, D.A.; Ahorlu, C.S. Review of MDA Registers for Lymphatic Filariasis: Findings, and Potential Uses in Addressing the Endgame Elimination Challenges. PLoS Negl. Trop. Dis. 2020, 14, e0008306. [Google Scholar] [CrossRef]
- Martin, R.J. Modes of Action of Anthelmintic Drugs. Vet. J. 1997, 154, 11–34. [Google Scholar] [CrossRef]
- Moreno, Y.; Nabhan, J.F.; Solomon, J.; Mackenzie, C.D.; Geary, T.G. Ivermectin Disrupts the Function of the Excretory-Secretory Apparatus in Microfilariae of Brugia malayi. Proc. Natl. Acad. Sci. USA 2010, 107, 20120–20125. [Google Scholar] [CrossRef]
- Maclean, M.J.; Lorenz, W.W.; Dzimianski, M.T.; Anna, C.; Moorhead, A.R.; Reaves, B.J.; Wolstenholme, A.J. Effects of Diethylcarbamazine and Ivermectin Treatment on Brugia malayi Gene Expression in Infected Gerbils (Meriones unguiculatus). Parasitol. Open 2019, 5, e2. [Google Scholar] [CrossRef][Green Version]
- Norling, L.V.; Sampaio, A.L.F.; Cooper, D.; Perretti, M. Inhibitory Control of Endothelial Galectin-1 on in Vitro and in Vivo Lymphocyte Trafficking. FASEB J. 2008, 22, 682–690. [Google Scholar] [CrossRef] [PubMed]
- MacKinnon, A.C.; Farnworth, S.L.; Hodkinson, P.S.; Henderson, N.C.; Atkinson, K.M.; Leffler, H.; Nilsson, U.J.; Haslett, C.; Forbes, S.J.; Sethi, T. Regulation of Alternative Macrophage Activation by Galectin-3. J. Immunol. 2008, 180, 2650–2658. [Google Scholar] [CrossRef]
- Wang, W.; Wang, S.; Zhang, H.; Yuan, C.; Yan, R.; Song, X.; Xu, L.; Li, X. Galectin Hco-Gal-m from Haemonchus Contortus Modulates Goat Monocytes and T Cell Function in Different Patterns. Parasites Vectors 2014, 7, 342. [Google Scholar] [CrossRef]
- Bahekar, S.P.; Hande, S.V.; Agrawal, N.R.; Chandak, H.S.; Bhoj, P.S.; Goswami, K.; Reddy, M.V.R. Sulfonamide Chalcones: Synthesis and in Vitro Exploration for Therapeutic Potential against Brugia malayi. Eur. J. Med. Chem. 2016, 124, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Bhoj, P.S.; Bahekar, S.; Khatri, V.; Singh, N.; Togre, N.S.; Goswami, K.; Chandak, H.S.; Dash, D. Role of Glutathione in Chalcone Derivative Induced Apoptosis of Brugia malayi and Its Possible Therapeutic Implication. Acta Parasitol. 2021, 66, 406–415. [Google Scholar] [CrossRef] [PubMed]
- Kalyanasundaram, R.; Khatri, V.; Chauhan, N. Advances in Vaccine Development for Human Lymphatic Filariasis. Trends Parasitol. 2020, 36, 195–205. [Google Scholar] [CrossRef]
- Thirugnanam, S.; Pandiaraja, P.; Ramaswamy, K.; Murugan, V.; Gnanasekar, M.; Nandakumar, K.; Reddy, M.V.R.; Kaliraj, P. Brugia malayi: Comparison of Protective Immune Responses Induced by Bm-Alt-2 DNA, Recombinant Bm-ALT-2 Protein and Prime-Boost Vaccine Regimens in a Jird Model. Exp. Parasitol. 2007, 116, 483–491. [Google Scholar] [CrossRef]
- Dakshinamoorthy, G.; Munirathinam, G.; Stoicescu, K.; Reddy, M.V.; Kalyanasundaram, R. Large Extracellular Loop of Tetraspanin as a Potential Vaccine Candidate for Filariasis. PLoS ONE 2013, 8, e77394. [Google Scholar] [CrossRef]
- Dakshinamoorthy, G.; Kalyanasundaram, R. Evaluating the Efficacy of RBmHATαc as a Multivalent Vaccine against Lymphatic Filariasis in Experimental Animals and Optimizing the Adjuvant Formulation. Vaccine 2013, 32, 19–25. [Google Scholar] [CrossRef][Green Version]
- Dakshinamoorthy, G.; von Gegerfelt, A.; Andersen, H.; Lewis, M.; Kalyanasundaram, R. Evaluation of a Multivalent Vaccine against Lymphatic Filariasis in Rhesus Macaque Model. PLoS ONE 2014, 9, e112982. [Google Scholar] [CrossRef]
- Chauhan, N.; Khatri, V.; Banerjee, P.; Kalyanasundaram, R. Evaluating the Vaccine Potential of a Tetravalent Fusion Protein (RBmHAXT) Vaccine Antigen Against Lymphatic Filariasis in a Mouse Model. Front. Immunol. 2018, 9, 01520. [Google Scholar] [CrossRef] [PubMed]
- Khatri, V.; Chauhan, N.; Vishnoi, K.; von Gegerfelt, A.; Gittens, C.; Kalyanasundaram, R. Prospects of Developing a Prophylactic Vaccine against Human Lymphatic Filariasis—Evaluation of Protection in Non-Human Primates. Int. J. Parasitol. 2018, 48, 773–783. [Google Scholar] [CrossRef] [PubMed]
- Zang, X.; Atmadja, A.K.; Gray, P.; Allen, J.E.; Gray, C.A.; Lawrence, R.A.; Yazdanbakhsh, M.; Maizels, R.M. The Serpin Secreted by Brugia malayi Microfilariae, Bm-SPN-2, Elicits Strong, but Short-Lived, Immune Responses in Mice and Humans. J. Immunol. 2000, 165, 5161–5169. [Google Scholar] [CrossRef] [PubMed]
- Veerapathran, A.; Dakshinamoorthy, G.; Gnanasekar, M.; Reddy, M.V.R.; Kalyanasundaram, R. Evaluation of Wuchereria bancrofti GST as a Vaccine Candidate for Lymphatic Filariasis. PLoS Negl. Trop. Dis. 2009, 3, e457. [Google Scholar] [CrossRef] [PubMed]
- Andure, D.; Pote, K.; Khatri, V.; Amdare, N.; Padalkar, R.; Reddy, M.V.R. Immunization with Wuchereria bancrofti Glutathione-S-Transferase Elicits a Mixed Th1/Th2 Type of Protective Immune Response Against Filarial Infection in Mastomys. Indian J. Clin. Biochem. 2016, 31, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Dakshinamoorthy, G.; Samykutty, A.K.; Munirathinam, G.; Reddy, M.V.; Kalyanasundaram, R. Multivalent Fusion Protein Vaccine for Lymphatic Filariasis. Vaccine 2013, 31, 1616–1622. [Google Scholar] [CrossRef]
- Kushwaha, S.; Singh, P.K.; Rana, A.K.; Misra-Bhattacharya, S. Immunization of Mastomys Coucha with Brugia malayi Recombinant Trehalose-6-Phosphate Phosphatase Results in Significant Protection against Homologous Challenge Infection. PLoS ONE 2013, 8, e72585. [Google Scholar] [CrossRef]
- Prince, P.R.; Madhumathi, J.; Anugraha, G.; Jeyaprita, P.J.; Reddy, M.V.R.; Kaliraj, P. Tandem Antioxidant Enzymes Confer Synergistic Protective Responses in Experimental Filariasis. J. Helminthol. 2014, 88, 402–410. [Google Scholar] [CrossRef]
- Arumugam, S.; Wei, J.; Ward, D.; Abraham, D.; Lustigman, S.; Zhan, B.; Klei, T.R. Vaccination with a Genetically Modified Brugia malayi Cysteine Protease Inhibitor-2 Reduces Adult Parasite Numbers and Affects the Fertility of Female Worms Following a Subcutaneous Challenge of Mongolian Gerbils (Meriones unguiculatus) with B. malayi Infective Larvae. Int. J. Parasitol. 2014, 44, 675–679. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Paul, R.; Ilamaran, M.; Khatri, V.; Amdare, N.; Reddy, M.V.R.; Kaliraj, P. Immunological Evaluation of Fusion Protein of Brugia malayi Abundant Larval Protein Transcript-2 (BmALT-2) and Tuftsin in Experimental Mice Model. Parasite Epidemiol. Control 2019, 4, e00092. [Google Scholar] [CrossRef]
- Yadav, S.; Sharma, P.; Sharma, A.; Ganga, L.; Saxena, J.K.; Srivastava, M. Immunization with Brugia malayi Calreticulin Protein Generates Robust Antiparasitic Immunity and Offers Protection during Experimental Lymphatic Filariasis. ACS Infect. Dis. 2021, 7, 790–799. [Google Scholar] [CrossRef] [PubMed]
- Khatri, V.; Chauhan, N.; Kalyanasundaram, R. Fecundity of Adult Female Worms Were Affected When Brugia malayi Infected Mongolian Gerbils Were Immunized with a Multivalent Vaccine (RBmHAXT) against Human Lymphatic Filarial Parasite. Acta Trop. 2020, 208, 105487. [Google Scholar] [CrossRef]
- Metenou, S.; Babu, S.; Nutman, T.B. Impact of Filarial Infections on Coincident Intracellular Pathogens. Curr. Opin. HIV AIDS 2012, 7, 231–238. [Google Scholar] [CrossRef]
- Yan, Y.; Inuo, G.; Akao, N.; Tsukidate, S.; Fujita, K. Down-Regulation of Murine Susceptibility to Cerebral Malaria by Inoculation with Third-Stage Larvae of the Filarial Nematode Brugia pahangi. Parasitology 1997, 114, 333–338. [Google Scholar] [CrossRef]
- Graham, A.L.; Lamb, T.J.; Read, A.F.; Allen, J.E. Malaria-Filaria Coinfection in Mice Makes Malarial Disease More Severe Unless Filarial Infection Achieves Patency. J. Infect. Dis. 2005, 191, 410–421. [Google Scholar] [CrossRef]
- Fernández Ruiz, D.; Dubben, B.; Saeftel, M.; Endl, E.; Deininger, S.; Hoerauf, A.; Specht, S. Filarial Infection Induces Protection against P. berghei Liver Stages in Mice. Microbes Infect. 2009, 11, 172–180. [Google Scholar] [CrossRef]
- Metenou, S.; Dembélé, B.; Konate, S.; Dolo, H.; Coulibaly, S.Y.; Coulibaly, Y.I.; Diallo, A.A.; Soumaoro, L.; Coulibaly, M.E.; Sanogo, D.; et al. Patent Filarial Infection Modulates Malaria-Specific Type 1 Cytokine Responses in an IL-10-Dependent Manner in a Filaria/Malaria-Coinfected Population. J. Immunol. 2009, 183, 916–924. [Google Scholar] [CrossRef]
- Specht, S.; Ruiz, D.F.; Dubben, B.; Deininger, S.; Hoerauf, A. Filaria-Induced IL-10 Suppresses Murine Cerebral Malaria. Microbes Infect. 2010, 12, 635–642. [Google Scholar] [CrossRef]
- Panda, M.; Sahoo, P.K.; das Mohapatra, A.; kanti Dutta, S.; Thatoi, P.K.; Tripathy, R.; Das, B.K.; Satpathy, A.K.; Ravindran, B. Decreased Prevalence of Sepsis but Not Mild or Severe P. falciparum Malaria Is Associated with Pre-Existing Filarial Infection. Parasit Vectors 2013, 6, 203. [Google Scholar] [CrossRef]
- Kolbaum, J.; Tartz, S.; Hartmann, W.; Helm, S.; Nagel, A.; Heussler, V.; Sebo, P.; Fleischer, B.; Jacobs, T.; Breloer, M. Nematode-Induced Interference with the Anti-Plasmodium CD8+ T-Cell Response Can Be Overcome by Optimizing Antigen Administration. Eur. J. Immunol. 2012, 42, 890–900. [Google Scholar] [CrossRef]
- Noland, G.S.; Chowdhury, D.R.; Urban, J.F.; Zavala, F.; Kumar, N. Helminth Infection Impairs the Immunogenicity of a Plasmodium Falciparum DNA Vaccine, but Not Irradiated Sporozoites, in Mice. Vaccine 2010, 28, 2917–2923. [Google Scholar] [CrossRef] [PubMed]
- Verma, R.; Joseph, S.K.; Kushwaha, V.; Kumar, V.; Siddiqi, M.I.; Vishwakarma, P.; Shivahare, R.; Gupta, S.; Murthy, P.K. Cross Reactive Molecules of Human Lymphatic Filaria Brugia malayi Inhibit Leishmania Donovani Infection in Hamsters. Acta Trop. 2015, 152, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Murthy, P.K.; Dixit, S.; Gaur, R.L.; Kumar, R.; Sahoo, M.K.; Shakya, N.; Joseph, S.K.; Palne, S.; Gupta, S. Influence of Brugia malayi Life Stages and BmAFII Fraction on Experimental Leishmania Donovani Infection in Hamsters. Acta Trop. 2008, 106, 81–89. [Google Scholar] [CrossRef]
- Sahoo, M.K.; Sisodia, B.S.; Dixit, S.; Joseph, S.K.; Gaur, R.L.; Verma, S.K.; Verma, A.K.; Shasany, A.K.; Dowle, A.A.; Murthy, P.K. Immunization with Inflammatory Proteome of Brugia malayi Adult Worm Induces a Th1/Th2-Immune Response and Confers Protection against the Filarial Infection. Vaccine 2009, 27, 4263–4271. [Google Scholar] [CrossRef]
- Kushwaha, V.; Kaur, S. Cross-Protective Efficacy of Immuno-Stimulatory Recombinant Brugia malayi Protein HSP60 against the Leishmania Donovani in BALB/c Mice. Biologicals 2021, 72, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Verma, R.; Kushwaha, V.; Pandey, S.; Thota, J.R.; Vishwakarma, P.; Parmar, N.; Yadav, P.K.; Tewari, P.; Kar, S.; Shukla, P.K.; et al. Leishmania Donovani Molecules Recognized by Sera of Filaria Infected Host Facilitate Filarial Infection. Parasitol. Res. 2018, 117, 2901–2912. [Google Scholar] [CrossRef]
- Sangare, M.B.; Coulibaly, Y.I.; Coulibaly, S.Y.; Coulibaly, M.E.; Traore, B.; Dicko, I.; Sissoko, I.M.; Samake, S.; Traore, S.F.; Nutman, T.B.; et al. A Cross-Sectional Study of the Filarial and Leishmania Co-Endemicity in Two Ecologically Distinct Settings in Mali. Parasit Vectors 2018, 11, 18. [Google Scholar] [CrossRef]
- Lamb, T.J.; Graham, A.L.; le Goff, L.; Allen, J.E. Co-Infected C57BL/6 Mice Mount Appropriately Polarized and Compartmentalized Cytokine Responses to Litomosoides Sigmodontis and Leishmania Major but Disease Progression Is Altered. Parasite Immunol. 2005, 27, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Rook, G.A.W. The Hygiene Hypothesis and the Increasing Prevalence of Chronic Inflammatory Disorders. Trans. R. Soc. Trop. Med. Hyg. 2007, 101, 1072–1074. [Google Scholar] [CrossRef]
- Caraballo, L. The Tropics, Helminth Infections and Hygiene Hypotheses. Expert Rev. Clin. Immunol. 2018, 14, 99–102. [Google Scholar] [CrossRef]
- Bach, J.-F. Revisiting the Hygiene Hypothesis in the Context of Autoimmunity. Front. Immunol. 2021, 11, 615192. [Google Scholar] [CrossRef]
- Smallwood, T.B.; Giacomin, P.R.; Loukas, A.; Mulvenna, J.P.; Clark, R.J.; Miles, J.J. Helminth Immunomodulation in Autoimmune Disease. Front. Immunol. 2017, 8, 00453. [Google Scholar] [CrossRef]
- Shi, W.; Xu, N.; Wang, X.; Vallée, I.; Liu, M.; Liu, X. Helminth Therapy for Immune-Mediated Inflammatory Diseases: Current and Future Perspectives. J. Inflamm. Res. 2022, 15, 475–491. [Google Scholar] [CrossRef]
- Khatri, V.; Amdare, N.; Yadav, R.S.; Tarnekar, A.; Goswami, K.; Reddy, M.V.R. Brugia malayi Abundant Larval Transcript 2 Protein Treatment Attenuates Experimentally-Induced Colitis in Mice. Indian J. Exp. Biol. 2015, 53, 732–739. [Google Scholar]
- Amdare, N.P.; Khatri, V.K.; Yadav, R.S.P.; Tarnekar, A.; Goswami, K.; Reddy, M.V.R. Therapeutic Potential of the Immunomodulatory Proteins Wuchereria bancrofti L2 and Brugia malayi Abundant Larval Transcript 2 against Streptozotocin-Induced Type 1 Diabetes in Mice. J. Helminthol. 2017, 91, 539–548. [Google Scholar] [CrossRef]
- Ou, B.; Yang, Y.; Tham, W.L.; Chen, L.; Guo, J.; Zhu, G. Genetic Engineering of Probiotic Escherichia Coli Nissle 1917 for Clinical Application. Appl. Microbiol. Biotechnol. 2016, 100, 8693–8699. [Google Scholar] [CrossRef] [PubMed]
- Khatri, V.; Amdare, N.; Tarnekar, A.; Goswami, K.; Reddy, M.V.R. Brugia malayi Cystatin Therapeutically Ameliorates Dextran Sulfate Sodium-Induced Colitis in Mice. J. Dig. Dis. 2015, 16, 585–594. [Google Scholar] [CrossRef]
- Bisht, N.; Khatri, V.; Chauhan, N.; Kalyanasundaram, R. Cystatin from Filarial Parasites Suppress the Clinical Symptoms and Pathology of Experimentally Induced Colitis in Mice by Inducing T-Regulatory Cells, B1-Cells, and Alternatively Activated Macrophages. Biomedicines 2019, 7, 85. [Google Scholar] [CrossRef]
- Yadav, R.S.P.; Khatri, V.; Amdare, N.; Goswami, K.; Shivkumar, V.B.; Gangane, N.; Reddy, M.V.R. Immuno-Modulatory Effect and Therapeutic Potential of Brugia malayi Cystatin in Experimentally Induced Arthritis. Indian J. Clin. Biochem. 2016, 31, 203–208. [Google Scholar] [CrossRef][Green Version]
- Yadav RS, P.; Khatri, V.; Amdare, N.; Goswami, K.; Shivkumar, V.B.; Gangane, N.; Reddy, M.V.R. Evaluation of Preventive Effect of Brugia malayi Recombinant Cystatin on MBSA-Induced Experimental Arthritis. Indian J. Exp. Biol. 2017, 55, 655–660. [Google Scholar]
- Khatri, V.; Chauhan, N.; Prasanna Kumar, S.B.; Kalyanasundaram, R. Peptide Fragments of Cystatin Protein from Filarial Parasite Has Potent Anti-Inflammatory Effect on DSS-Induced Colitis in Mouse. J. Immunol. 2020, 204, 237.28. [Google Scholar]
- Togre, N.; Bhoj, P.; Amdare, N.; Goswami, K.; Tarnekar, A.; Shende, M. Immunomodulatory Potential of Recombinant Filarial Protein, RWbL2, and Its Therapeutic Implication in Experimental Ulcerative Colitis in Mouse. Immunopharmacol. Immunotoxicol. 2018, 40, 483–490. [Google Scholar] [CrossRef]
- Togre, N.; Bhoj, P.; Goswami, K.; Tarnekar, A.; Patil, M.; Shende, M. Human Filarial Proteins Attenuate Chronic Colitis in an Experimental Mouse Model. Parasite Immunol. 2018, 40, e12511. [Google Scholar] [CrossRef] [PubMed]
- Amdare, N.; Khatri, V.; Yadav, R.S.P.; Tarnekar, A.; Goswami, K.; Reddy, M.V.R. Brugia malayi Soluble and Excretory-Secretory Proteins Attenuate Development of Streptozotocin-Induced Type 1 Diabetes in Mice. Parasite Immunol. 2015, 37, 624–634. [Google Scholar] [CrossRef] [PubMed]
- Kron, M.A.; Metwali, A.; Vodanovic-Jankovic, S.; Elliott, D. Nematode Asparaginyl-TRNA Synthetase Resolves Intestinal Inflammation in Mice with T-Cell Transfer Colitis. Clin. Vaccine Immunol. 2013, 20, 276–281. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Candidate Vaccine | Outcome | Reference |
|---|---|---|
| Serpin (BmSPN-2) | Immune response is strong but short-lived, suggesting that serpins alone are not effective vaccine candidates for long-term immunity | [84] |
| Aabundant larval transcript-2 (BmALT-2) | BmALT-2 protein and Bm-alt-2 DNA conferred approximately 75% and 57% protection, respectively | [78] |
| Glutathione-S-transferases (WbGST) | 61% protection in Jirds challenge experiments and 65.5% protection in in situ challenge studies | [29,85,86] |
| Small heat shock protein HSP12.6 (BmHsp12.6αc subunit) | 83% protection in mouse in situ challenge studies | |
| Large extracellular loop of tetraspanin (BmTSP-LEL) | 64% protection in mouse in situ challenge studies | [79] |
| Bivalent vaccines: HSP12.6 + ALT-2, HSP12.6 + TSP-LEL, TSP-LEL + ALT-2 | 90%, 80%, and 82% protection in mouse in situ challenge studies, respectively | [87] |
| Trehalose-6-phosphate phosphatase (BmTPP) | 78.4% decrease in microfilariae counts and 71% reduction in adult parasite load in Mastomys | [88] |
| Thioredoxin (WbTRX), Thioredoxin peroxidase (WbTPX) | 57% and 62% protection in Mastomys challenge experiments, respectively | [89] |
| BmHAT Trivalent vaccine | Protein and DNA protein prime boost vaccination yielded approximately 95% protection in mice 3 out of 5 vaccinated macaques were protected from challenge infection | [81,87] |
| Cystatin-2 in which the amino acid Asn66 was mutated to Lys66 (Bm-CPI-2M) | 48.6% and 48.0% at 42 and 90 days post-infection, respectively, with B. malayi L3 filariae | [90] |
| BmALT-2 with Tuftsin as fusion protein | 65% larvicidal activity in ADCC experiments | [91] |
| Calreticulin (BmCRT) | Offers protection during experimental lymphatic filariasis | [92] |
| BmHAXT Tetravalent vaccine | 88% protection in mouse in situ challenge studies 57% protection in rhesus macaques challenge infections Reduced fecundity and adult worm burden in Jirds | [82,83,93] |
| Lymphatic Filarial Protein | Experimental Disease Model | Study Outcome | Mechanism of Action | Reference |
|---|---|---|---|---|
| Recombinant B. malayi Cystatin (rBmCys) | DSS-induced acute colitis | Down-regulated inflammatory responses and alleviated symptoms and pathology of colitis. | Elevated IL-10 + FoxP3 + Tregs, IgM + B1a cells and AAMs in the colon and peritoneal cavity. Reduced expression of Th1 and Th17 cytokines in serum and spleen. | [118,119] |
| rBmCys | mBSA-induced rheumatoid arthritis (RA) | Both preventive and therapeutic effects on RA. Decreased synovitis, bone erosion, fibrosis, and influx of inflammatory cells in hind paw joints. | Shift from Th1 to IL-4 and IL-10 secreting Th2 immune response. | [120,121] |
| Peptide fragments of rBmCys | DSS-induced acute colitis | Anti-inflammatory effect on DSS-induced colitis in mice. Reversed the gross and histopathological changes in the colitic colon. | Decreased F4/80 + TLR-4 + CD11c+ macrophages in peritoneum. Reduced LY6G+ cells and MPO+ cells and increased FoxP3 + Tregs in colon. | [122] |
| Recombinant B. malayi abundant larval transcript-2 (rBmALT-2) | DSS-induced acute colitis | More effective in preventive mode compared to therapeutic treatment against colitis. | Associated with downregulation of IFN-γ, IL-6, IL-17, and upregulation of IL-10 cytokines in spleen. | [115] |
| Recombinant W. bancrofti L-2 (rWbL2) | DSS-induced acute colitis | Reduced lymphocyte infiltration and decreased epithelial damage in colons of treated mice. | Shift towards Th2 response as reflected by increased IL-10, and decreased IFN-γ and TNF-α by splenocytes. IgG1/IgG2 ratio in the sera. | [123] |
| rBmALT-2, rBmCys, and rWbL-2 individually and in combinations | DSS-induced chronic colitis | All treatment strategies improved the clinicopathologic status of chronic colitis. rBmALT-2 + rBmCys showed the most prominent therapeutic effect. | Downregulated IFN-γ and TNF-α expression, upregulated IL-10, and TGF-β expression in the splenocytes. Reduction in activated NF-κB level in the colon. Increased IgG1/IgG2 ratio in the sera. | [124] |
| rWbL-2, rBmALT-2, and rWbL-2 + rBmALT-2 | STZ-induced T1D | Led to reduced lymphocytic infiltration, islet damage, and blood glucose levels. | Decreased TNF-α and IFN-γ, and increased Il-4, IL-5, and IL-10 production in splenocytes. Elevated insulin-specific IgG1 and antigen-specific IgE antibodies in the sera. | [116] |
| B. malayi adult soluble (Bm A S) and microfilarial excretory-secretory proteins (Bm Mf ES) | STZ-induced T1D | More effective when used as curative rather than a preventive treatment. Reduced inflammatory changes in pancreatic islet cell architecture and fasting blood glucose levels. | Decreased TNF-α and IFN-γ, and increased IL-10 production in the splenocytes. Elevated anti-insulin IgG1 antibodies indicating a skewed response towards Th2 type in the sera. | [125] |
| B. malayi asparaginyl-tRNA synthetase (BmAsnRS) | T-cell transfer colitis | Resolves intestinal inflammation. | Increase in CD8+ T cells in the lamina propria compartment, with a corresponding increase in CD4+ cells in spleens of treated mice. Decrease in IFN-γ and IL-17, and increase in IL-4 and IL-10 in spleens, mesenteric lymph nodes, and lamina propria of treated mice. Induced upregulation of IL-10 and IL-22 receptors. | [126] |
| Brugia malayi K1 (BmK1) | - | Inhibits the delayed-type hypersensitivity response. | Blocked Kv1.3 receptors in human T cells. Suppressed the proliferation of rat CCR7-effector memory T cells and production of IFN-γ. | [40] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhoj, P.; Togre, N.; Khatri, V.; Goswami, K. Harnessing Immune Evasion Strategy of Lymphatic Filariae: A Therapeutic Approach against Inflammatory and Infective Pathology. Vaccines 2022, 10, 1235. https://doi.org/10.3390/vaccines10081235
Bhoj P, Togre N, Khatri V, Goswami K. Harnessing Immune Evasion Strategy of Lymphatic Filariae: A Therapeutic Approach against Inflammatory and Infective Pathology. Vaccines. 2022; 10(8):1235. https://doi.org/10.3390/vaccines10081235
Chicago/Turabian StyleBhoj, Priyanka, Namdev Togre, Vishal Khatri, and Kalyan Goswami. 2022. "Harnessing Immune Evasion Strategy of Lymphatic Filariae: A Therapeutic Approach against Inflammatory and Infective Pathology" Vaccines 10, no. 8: 1235. https://doi.org/10.3390/vaccines10081235
APA StyleBhoj, P., Togre, N., Khatri, V., & Goswami, K. (2022). Harnessing Immune Evasion Strategy of Lymphatic Filariae: A Therapeutic Approach against Inflammatory and Infective Pathology. Vaccines, 10(8), 1235. https://doi.org/10.3390/vaccines10081235