
Identification of Tumor Antigens and Immune Subtypes of Malignant Mesothelioma for mRNA Vaccine Development



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Extraction
2.2. Identification of Potential Tumor Antigens
2.3. Immune Subtypes Identification
2.4. Cellular and Molecular Characteristics of Immune Subtypes
2.5. Identification of Hub Genes
2.6. Statistical Analyses
3. Results
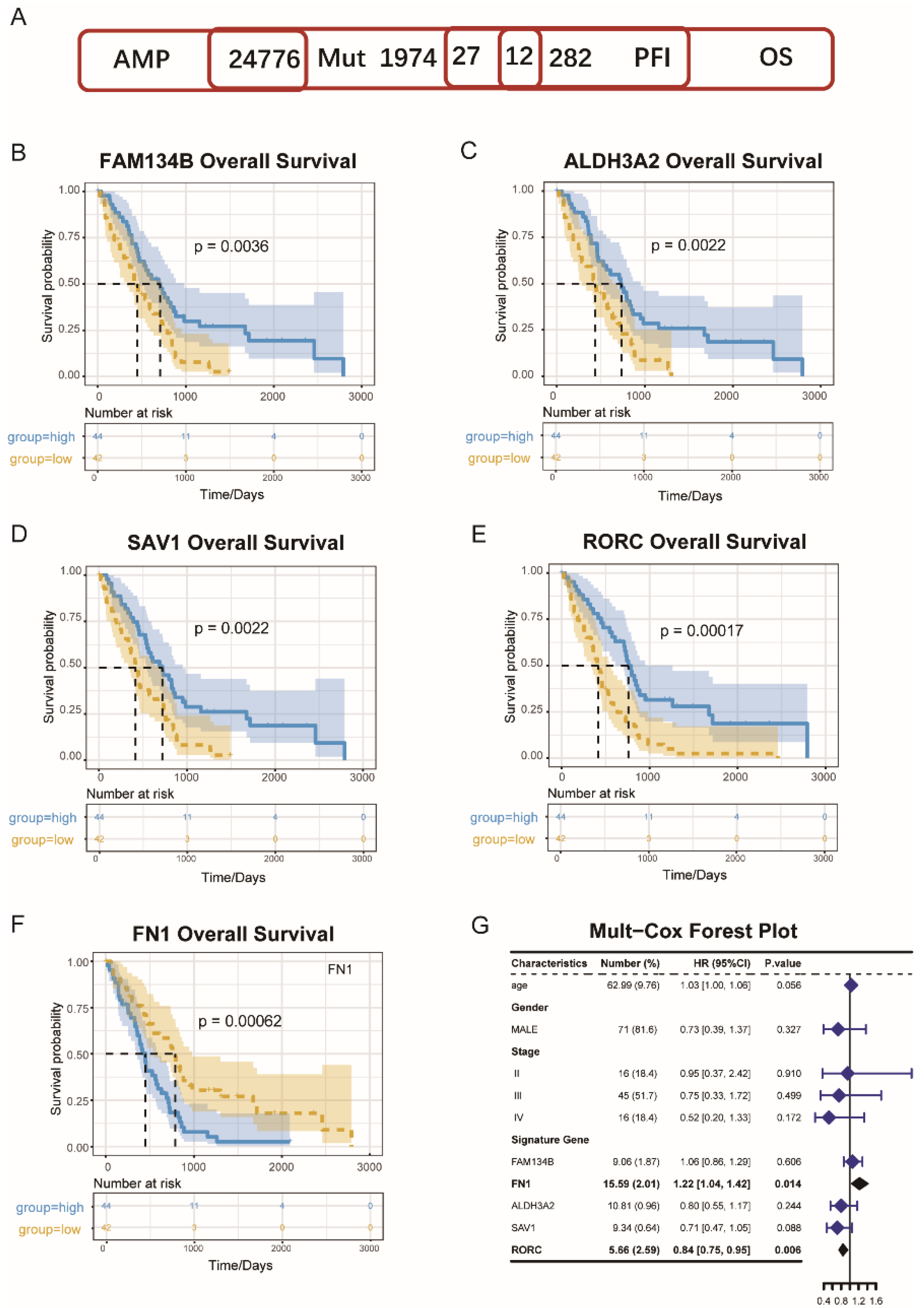
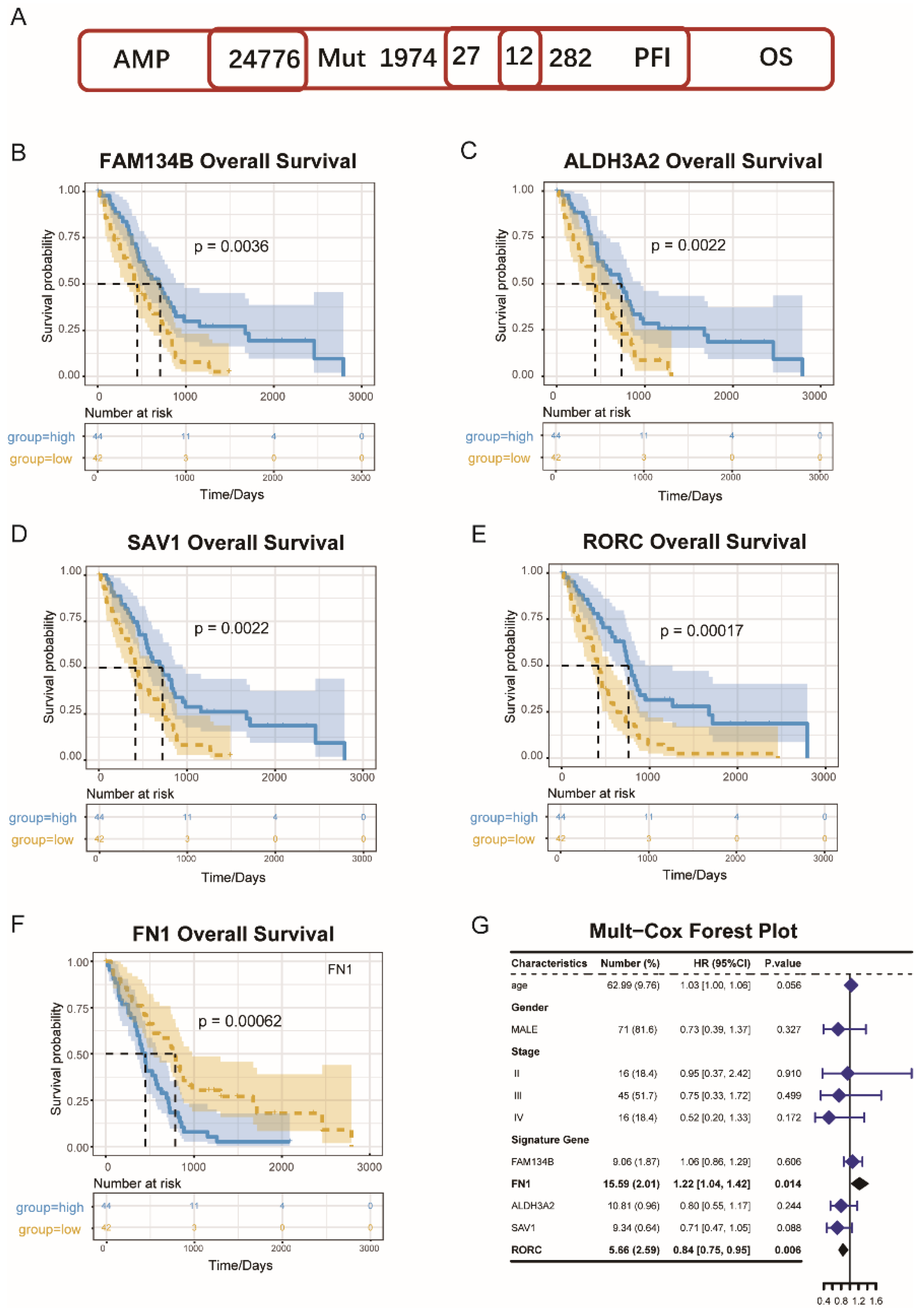
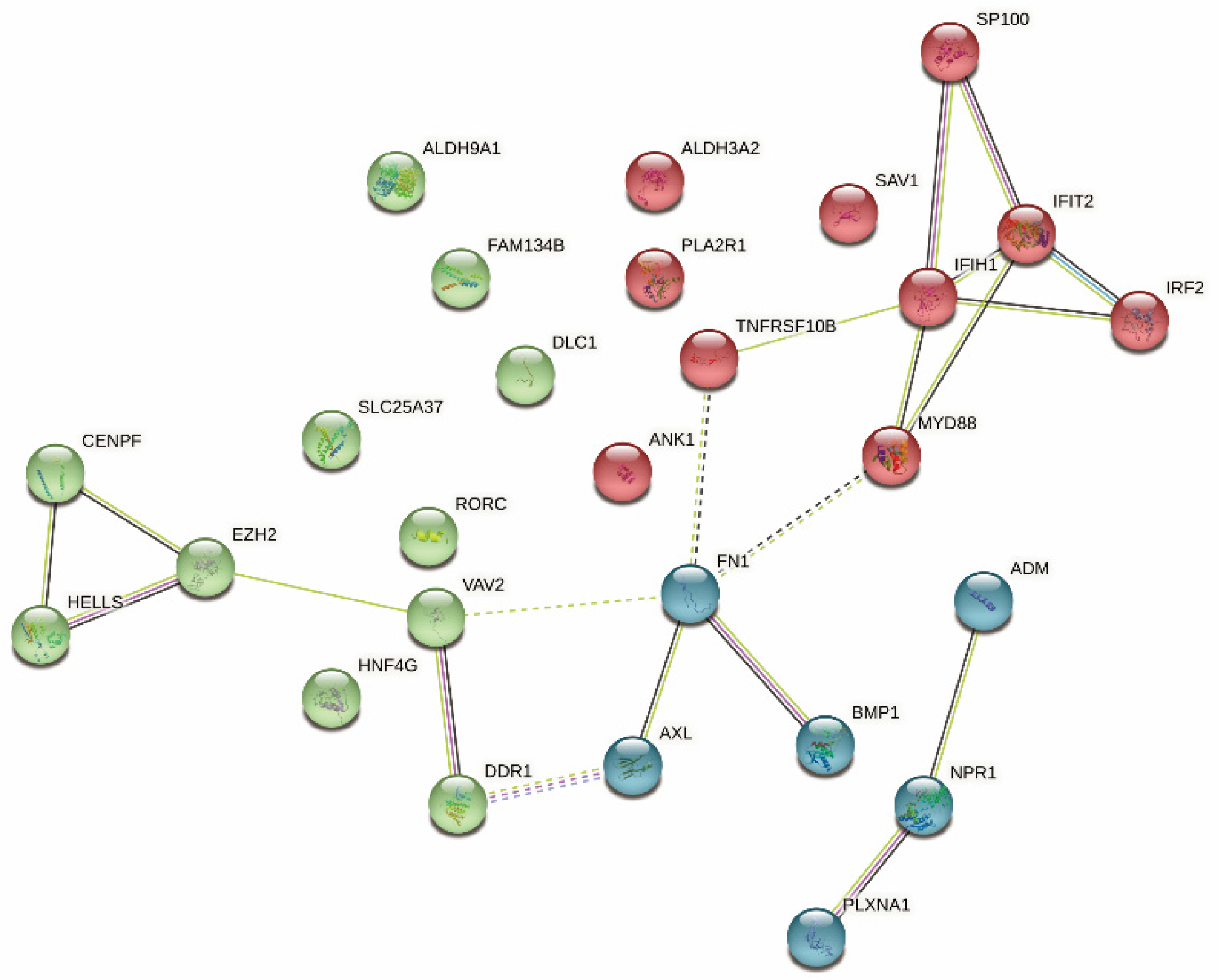
3.1. MESO Antigens Identification
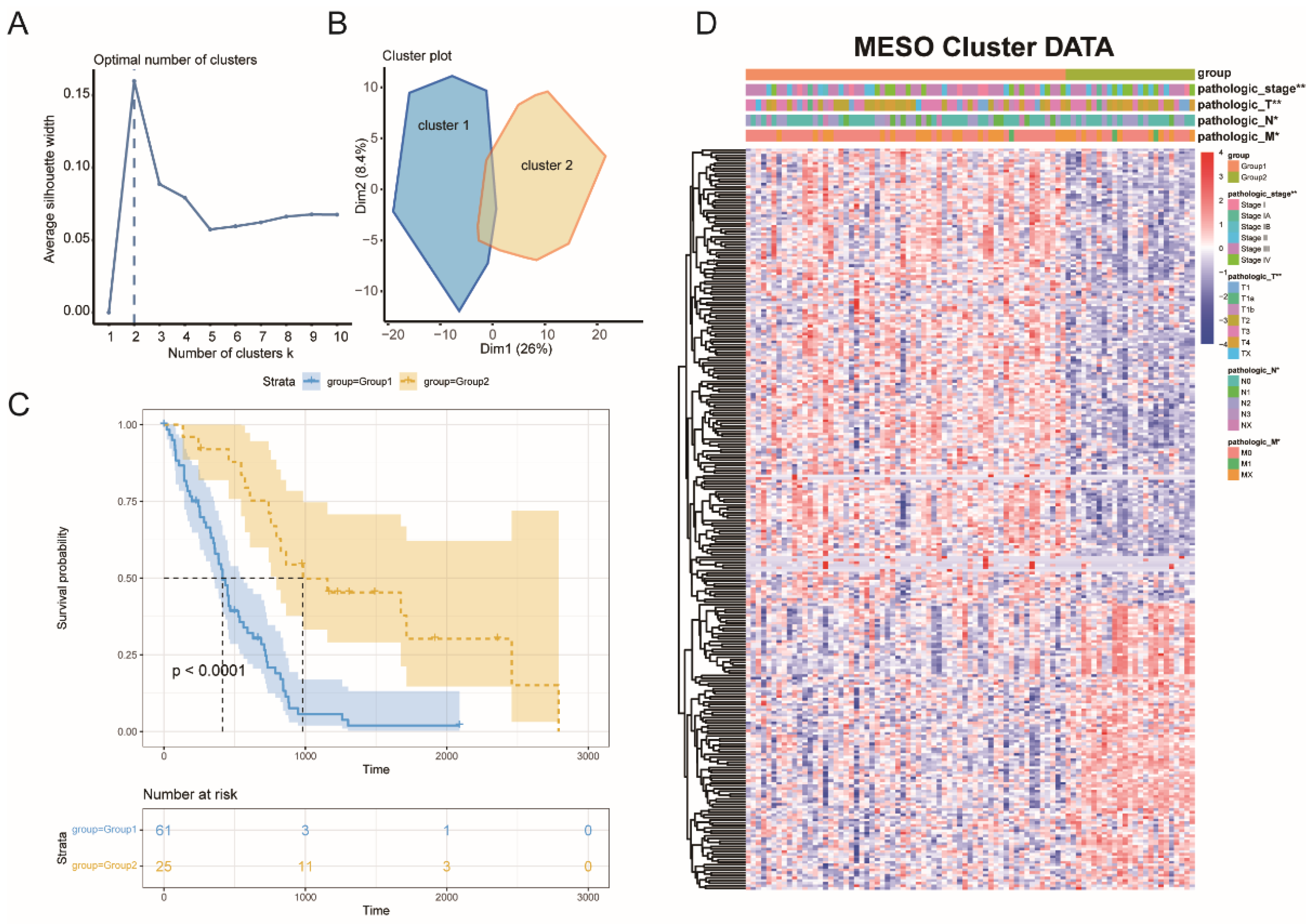
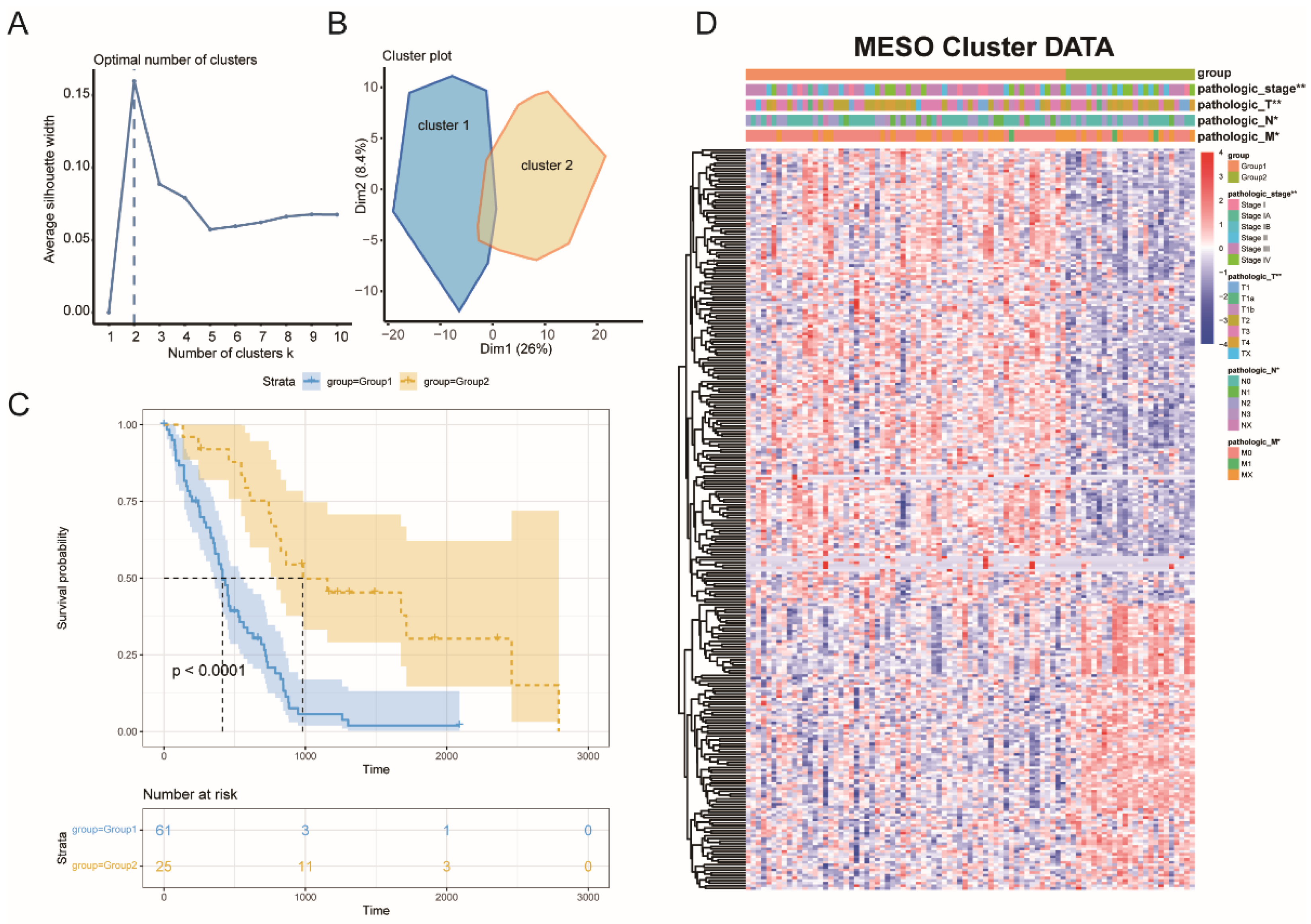
3.2. MESO Immune Subtypes Identification
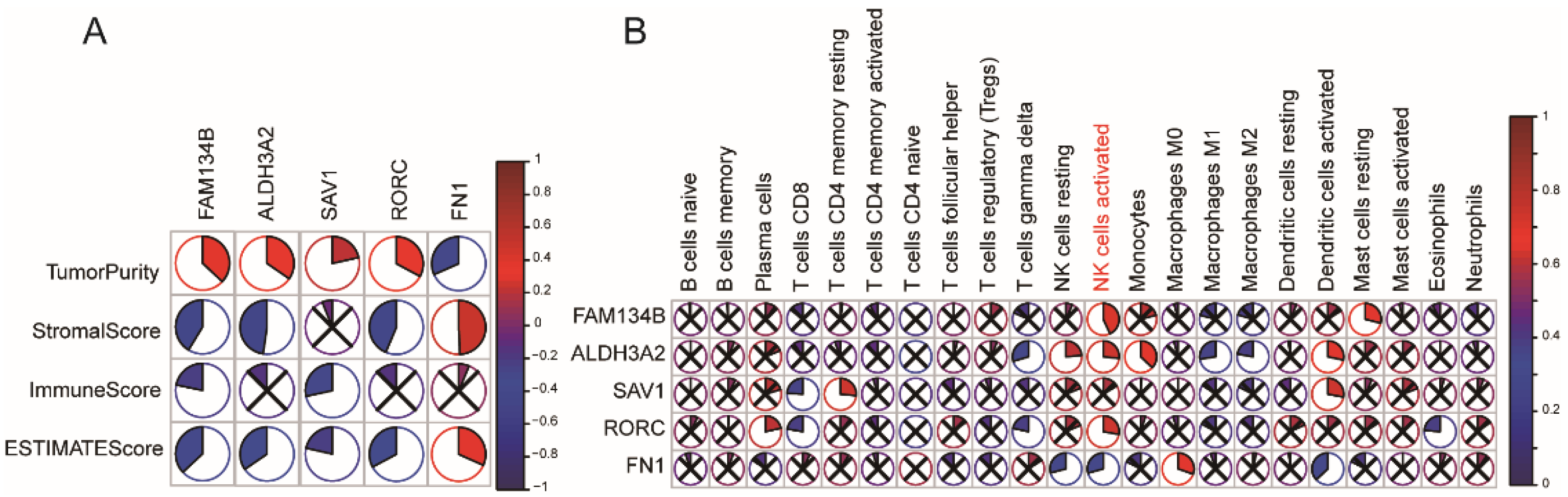
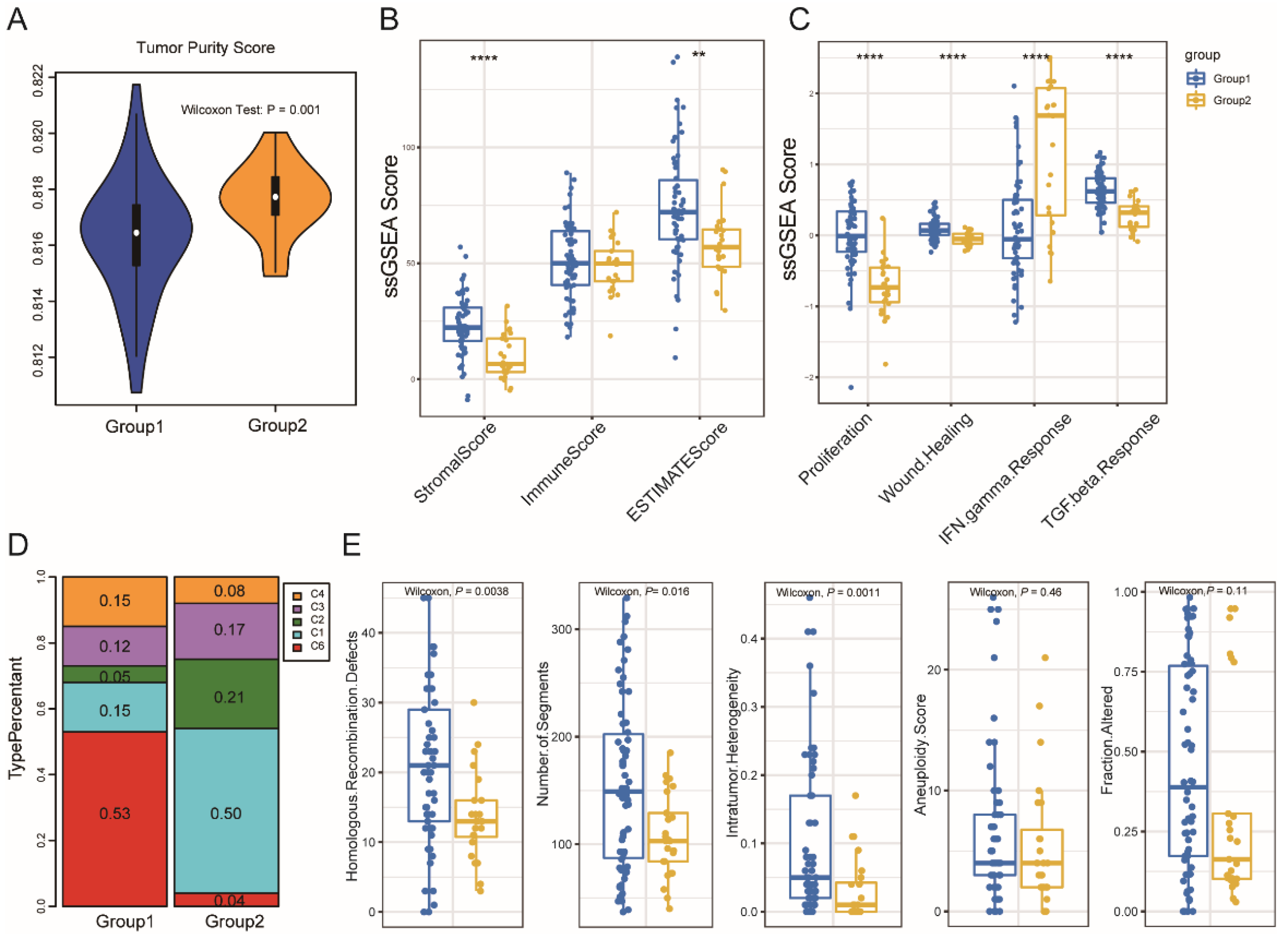
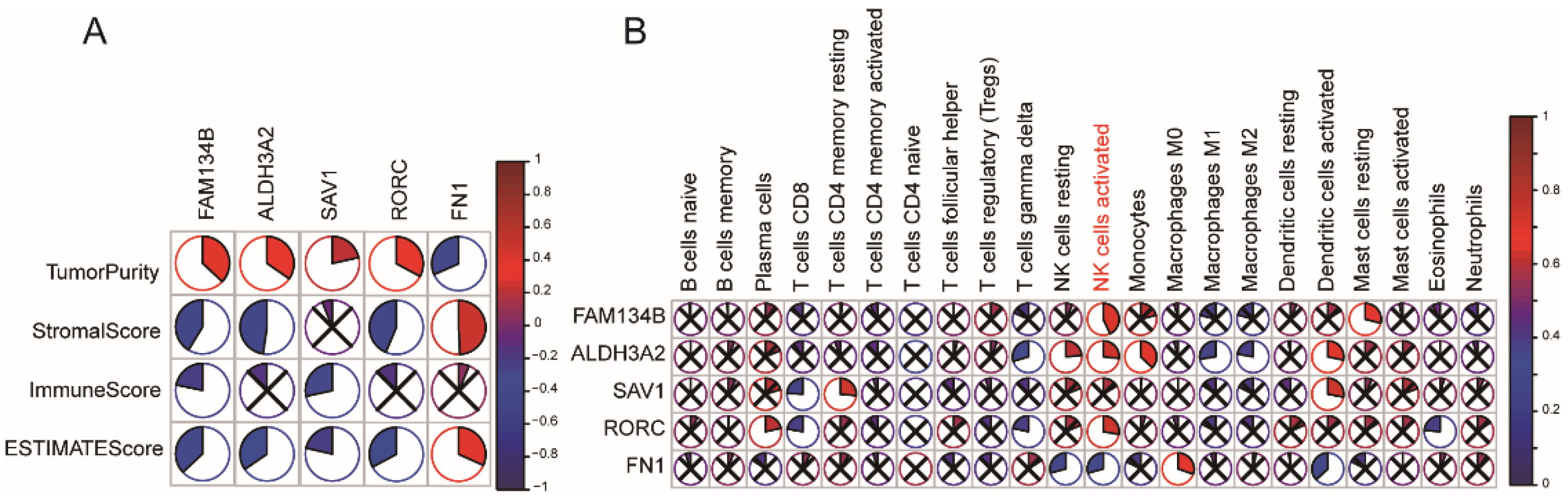
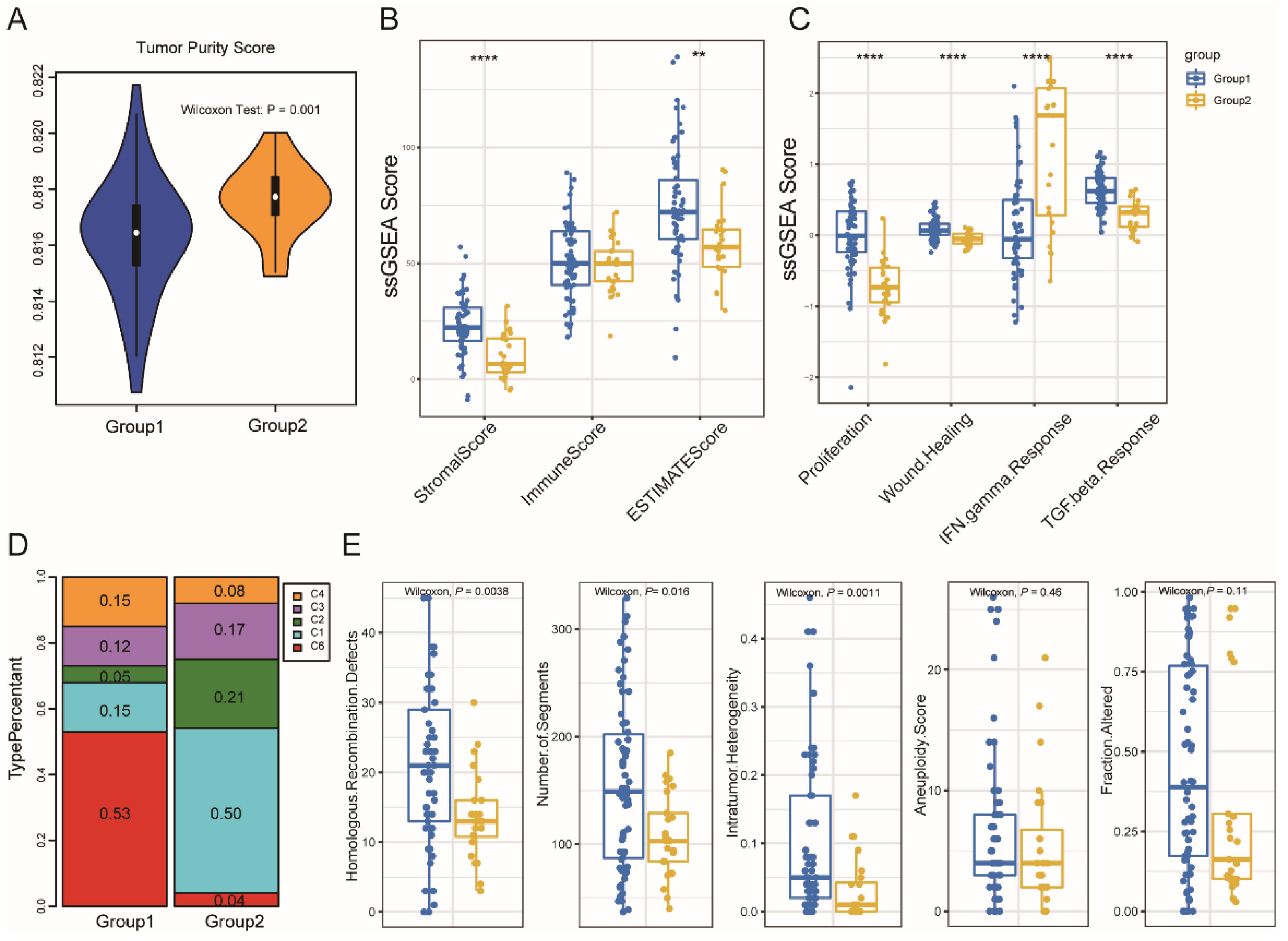
3.3. Immune Subtype Molecular and Cellular Characteristics
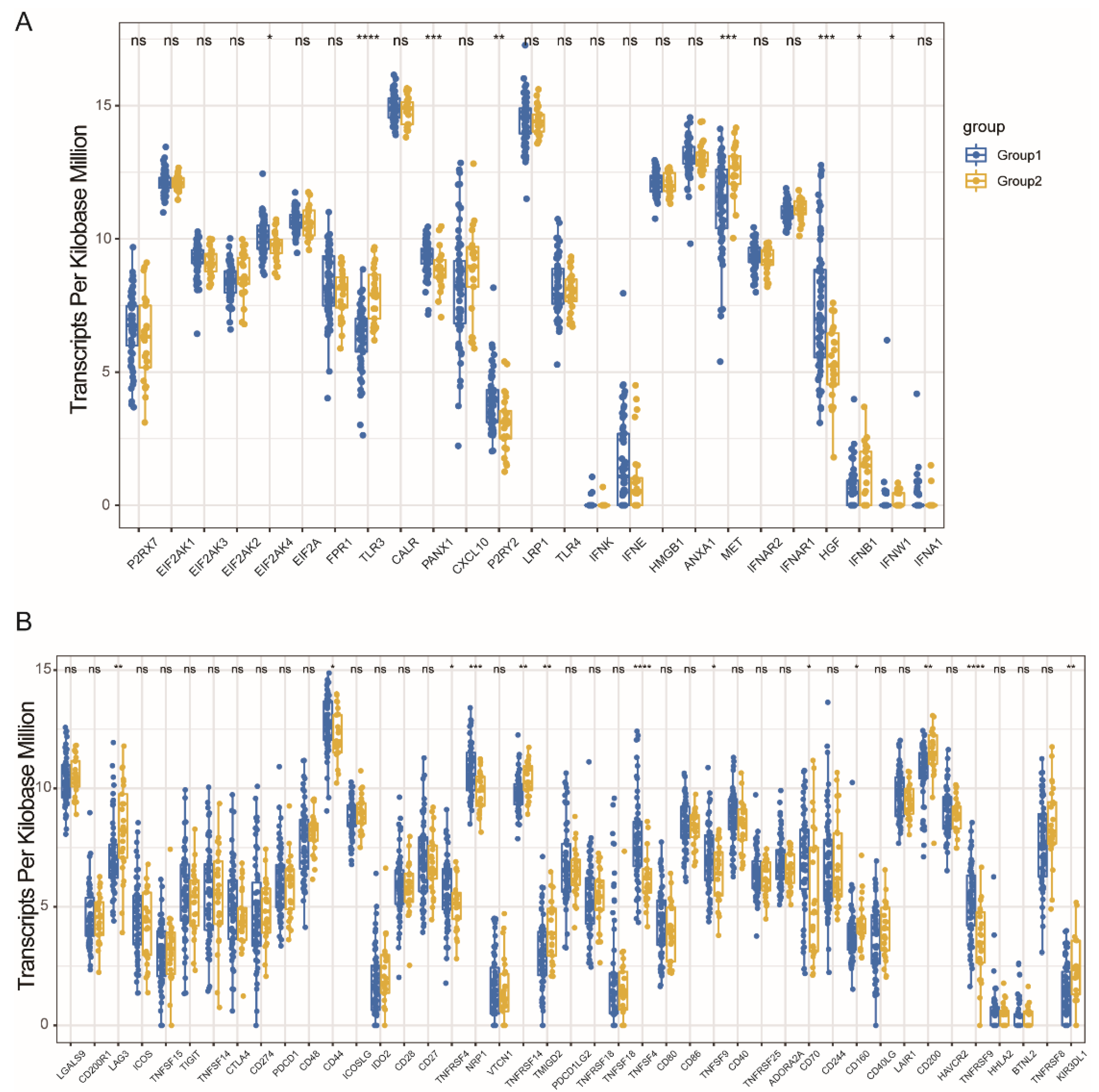
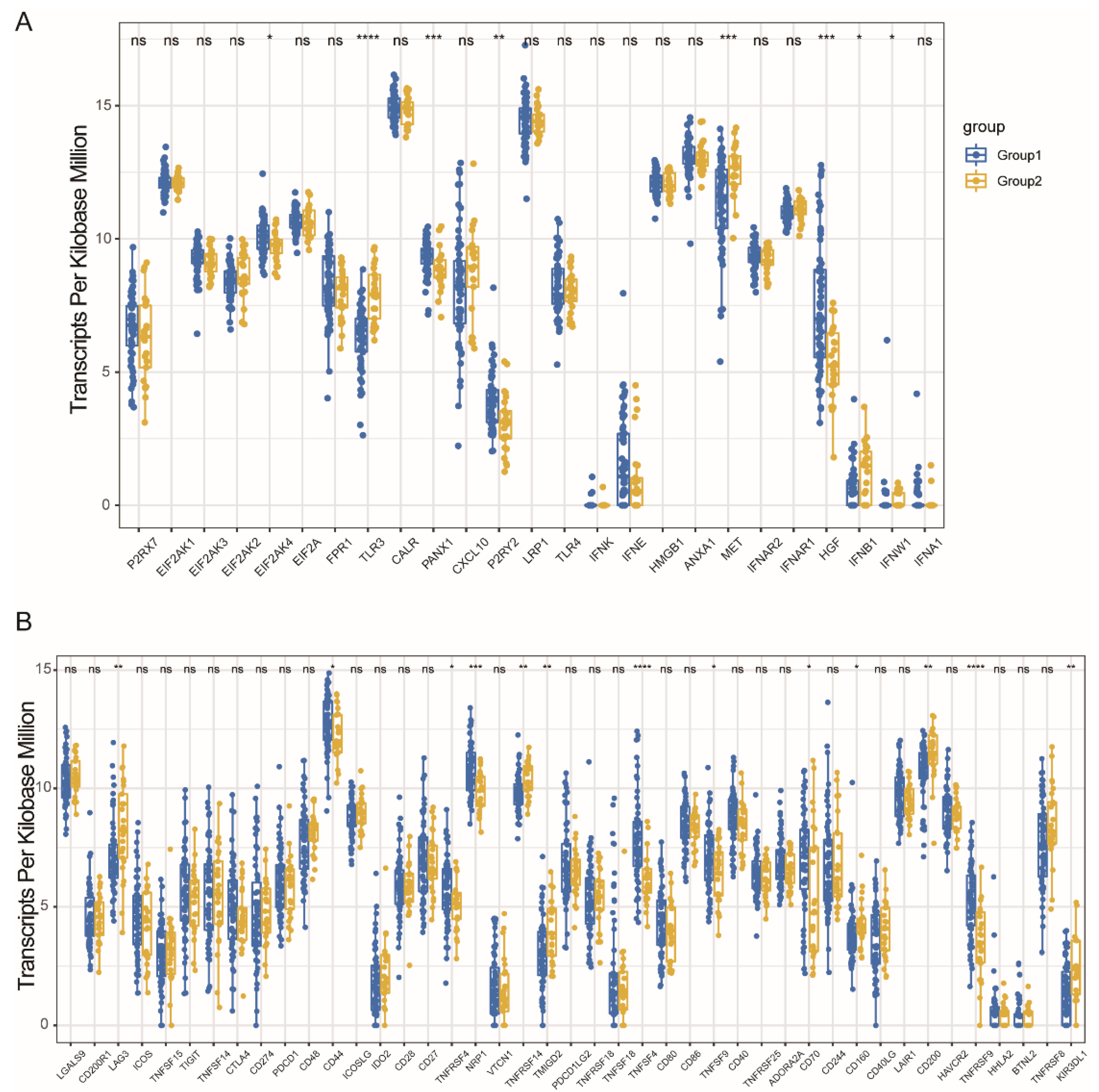
3.4. Immune Modulators and MESO Immune Subtypes Association
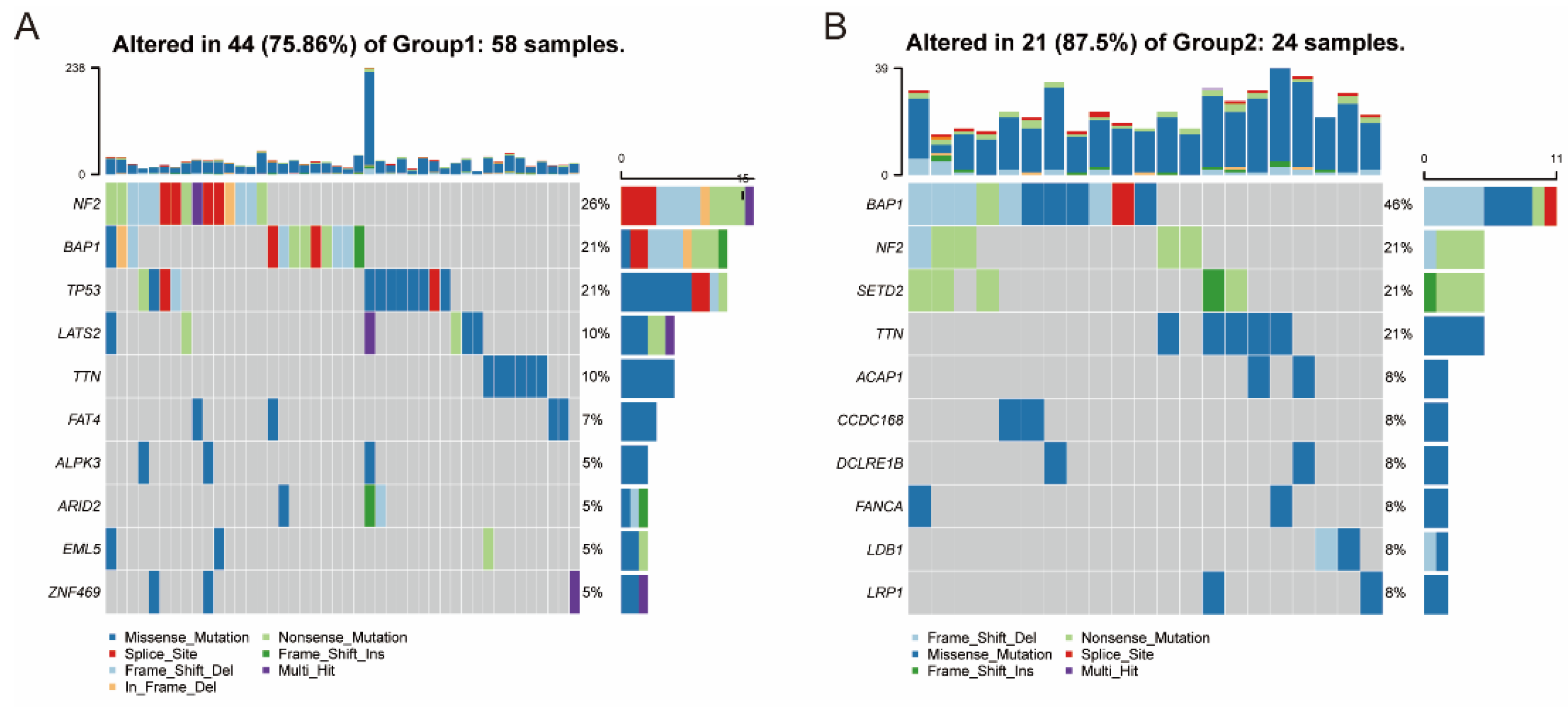
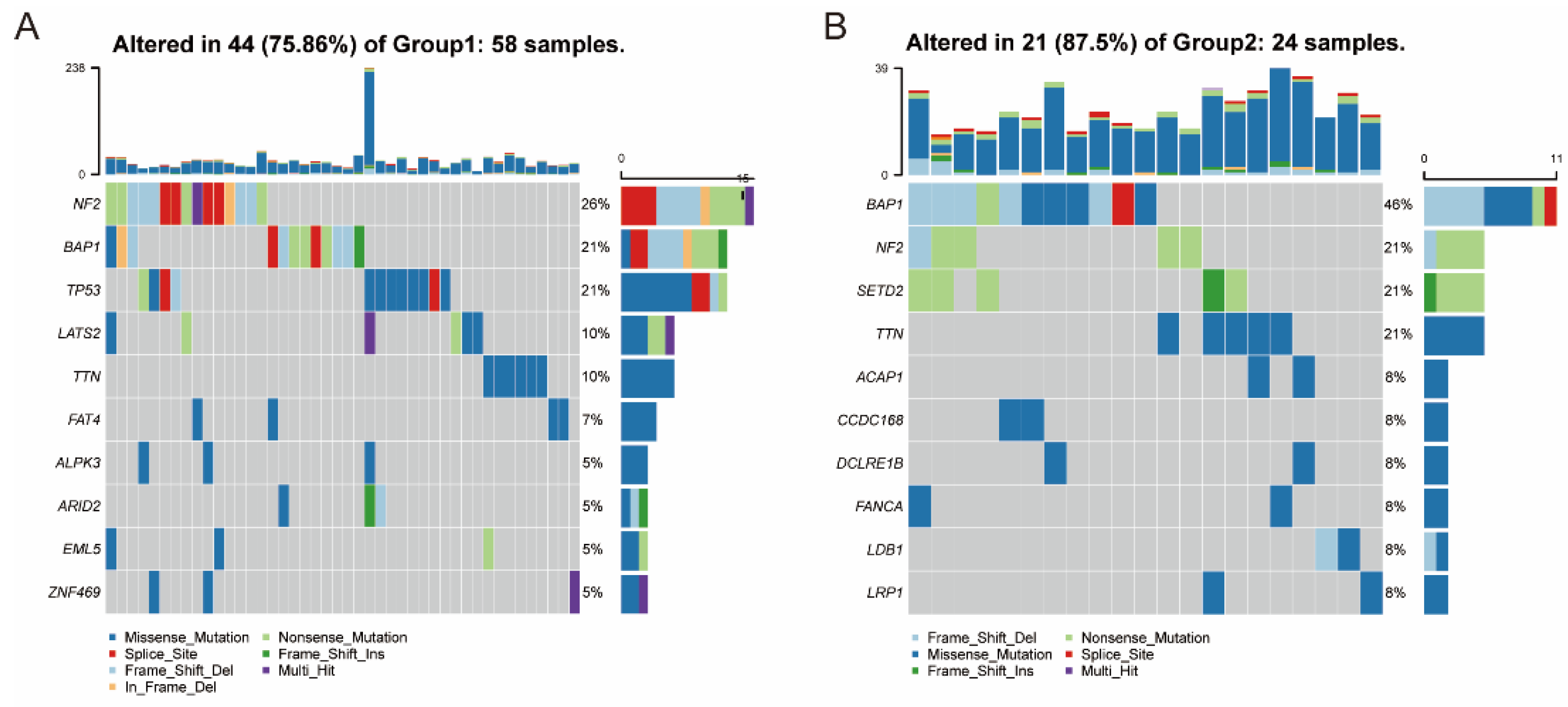
3.5. Association between Immune Subtypes and Mutational Status
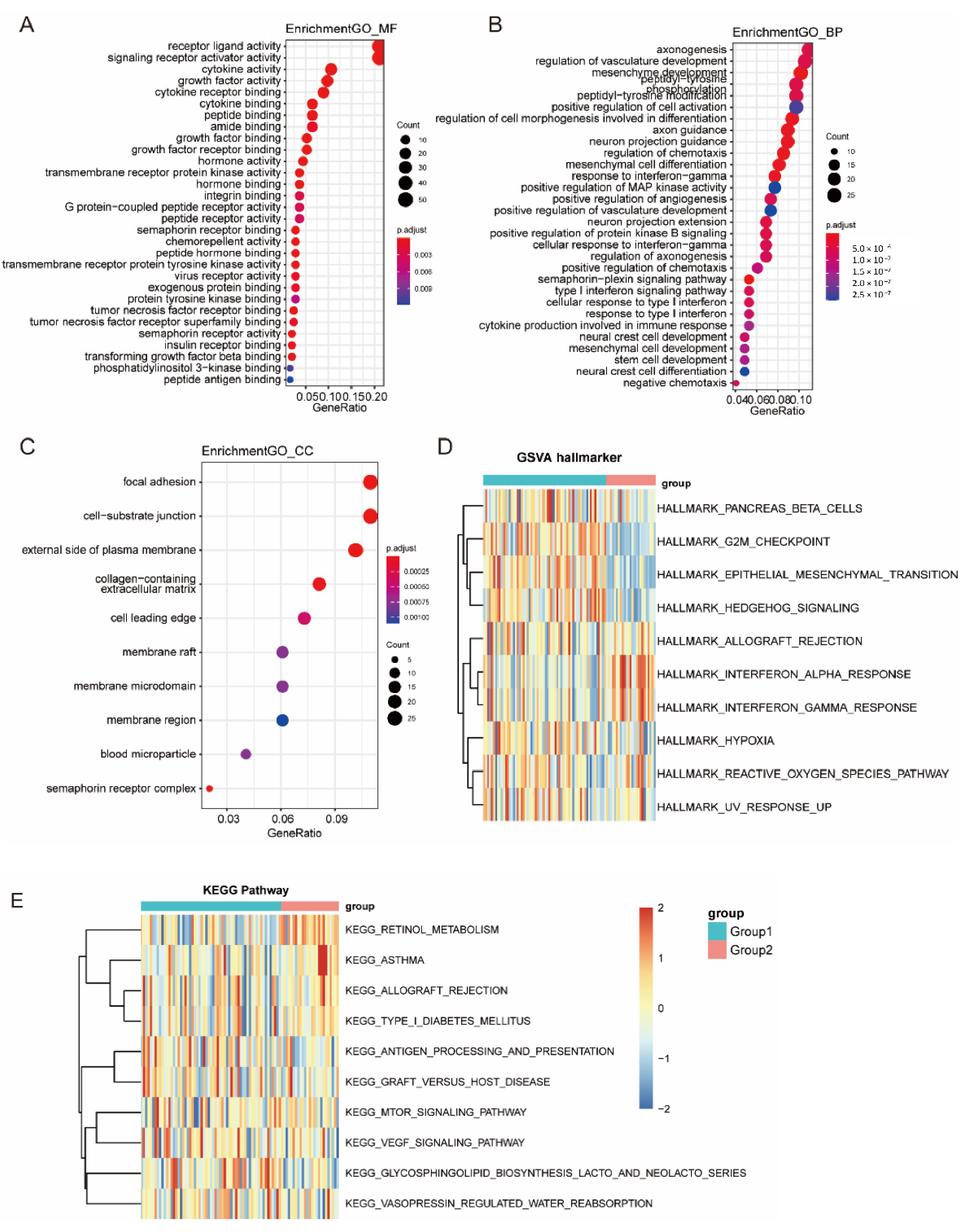
3.6. Function Enrichment Analysis of Immune Subtypes
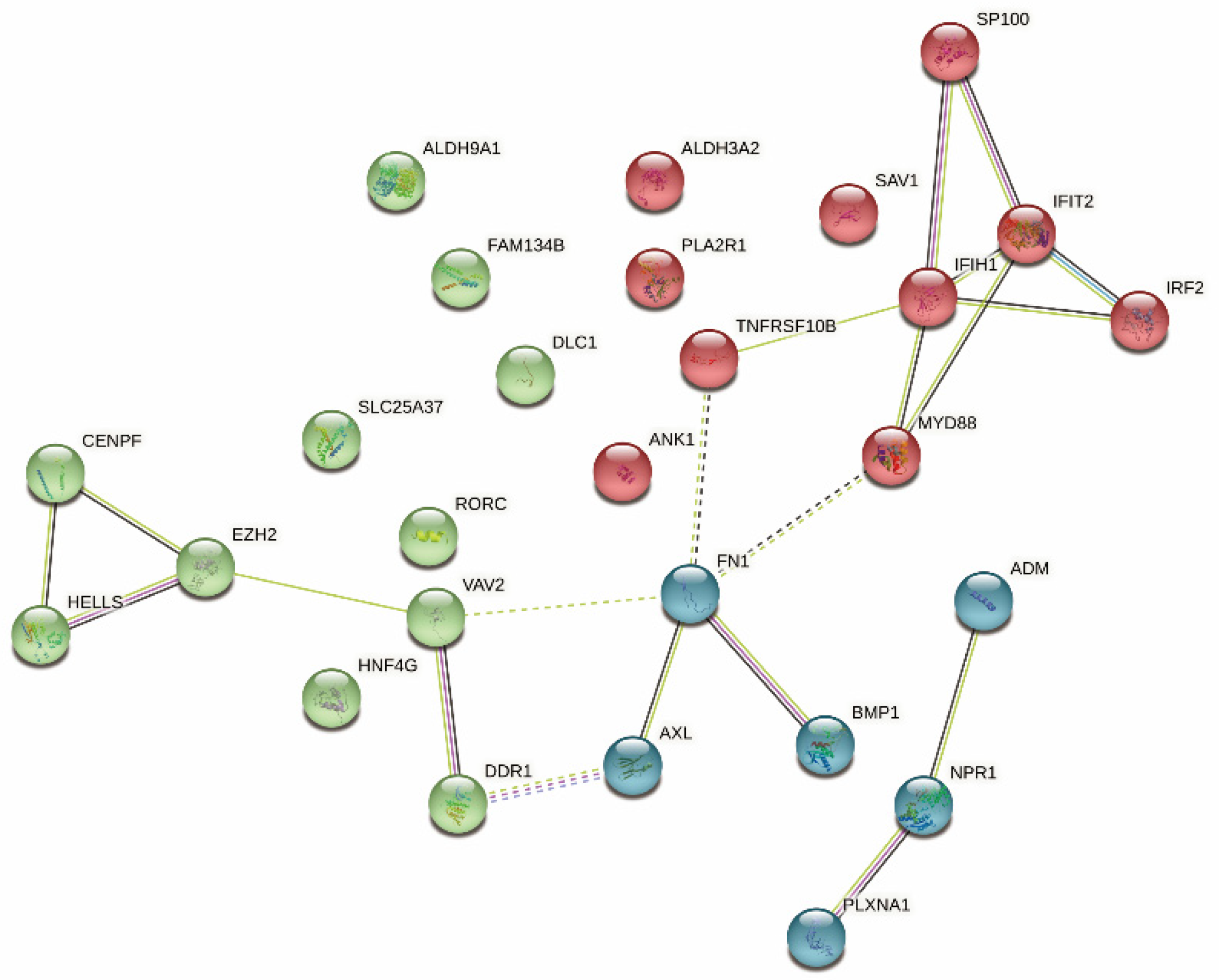
3.7. Identification of Immune Hub Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, S.; Chen, R.; Tang, Y.; Yu, Y.; Fang, Y.; Huang, H.; Wu, D.; Fang, H.; Bai, Y.; Sun, C.; et al. Comprehensive Genomic Profiling of Rare Tumors: Routes to Targeted Therapies. Front. Oncol. 2020, 10, 536. [Google Scholar] [CrossRef]
- Carbone, M.; Adusumilli, P.S.; Alexander, H.R., Jr.; Baas, P.; Bardelli, F.; Bononi, A.; Bueno, R.; Felley-Bosco, E.; Galateau-Salle, F.; Jablons, D.; et al. Mesothelioma: Scientific clues for prevention, diagnosis, and therapy. CA Cancer J. Clin. 2019, 69, 402–429. [Google Scholar] [CrossRef] [Green Version]
- Kindler, H.L.; Ismaila, N.; Armato, S.G., III; Bueno, R.; Hesdorffer, M.; Jahan, T.; Jones, C.M.; Miettinen, M.; Pass, H.; Rimner, A.; et al. Treatment of Malignant Pleural Mesothelioma: American Society of Clinical Oncology Clinical Practice Guideline. J. Clin. Oncol. 2018, 36, 1343–1373. [Google Scholar] [CrossRef]
- Mujoomdar, A.A.; Tilleman, T.R.; Richards, W.G.; Bueno, R.; Sugarbaker, D.J. Prevalence of in vitro chemotherapeutic drug resistance in primary malignant pleural mesothelioma: Result in a cohort of 203 resection specimens. J. Thorac. Cardiovasc. Surg. 2010, 140, 352–355. [Google Scholar] [CrossRef] [Green Version]
- Delgermaa, V.; Takahashi, K.; Park, E.-K.; Le, G.V.; Hara, T.; Sorahan, T. Global mesothelioma deaths reported to the World Health Organization between 1994 and 2008. Bull. World Health Organ. 2011, 89, 716–724. [Google Scholar] [CrossRef]
- Baas, P.; Scherpereel, A.; Nowak, A.K.; Fujimoto, N.; Peters, S.; Tsao, A.S.; Mansfield, A.S.; Popat, S.; Jahan, T.; Antonia, S.; et al. First-line nivolumab plus ipilimumab in unresectable malignant pleural mesothelioma (CheckMate 743): A multicentre, randomised, open-label, phase 3 trial. Lancet 2021, 397, 375–386. [Google Scholar] [CrossRef]
- Hu, Z.I.; Ghafoor, A.; Sengupta, M.; Hassan, R. Malignant mesothelioma: Advances in immune checkpoint inhibitor and mesothelin-targeted therapies. Cancer 2021, 127, 1010–1020. [Google Scholar] [CrossRef]
- Scherpereel, A.; Wallyn, F.; Albelda, S.M.; Munck, C. Novel therapies for malignant pleural mesothelioma. Lancet Oncol. 2018, 19, e161–e172. [Google Scholar] [CrossRef]
- Blum, Y.; Meiller, C.; Quetel, L.; Elarouci, N.; Ayadi, M.; Tashtanbaeva, D.; Armenoult, L.; Montagne, F.; Tranchant, R.; Renier, A.; et al. Dissecting heterogeneity in malignant pleural mesothelioma through histo-molecular gradients for clinical applications. Nat. Commun. 2019, 10, 1333. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.; Zhang, C.; Wu, X. Therapeutic cancer vaccines: From initial findings to prospects. Immunol. Lett. 2018, 196, 11–21. [Google Scholar] [CrossRef]
- Emens, L.A. Roadmap to a Better Therapeutic Tumor Vaccine. Int. Rev. Immunol. 2006, 25, 415–443. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, Y.; Sasada, T. Cancer Vaccines: Toward the Next Breakthrough in Cancer Immunotherapy. J. Immunol. Res. 2020, 2020, 5825401. [Google Scholar] [CrossRef] [PubMed]
- Hassan, R.; Alley, E.; Kindler, H.; Antonia, S.; Jahan, T.; Honarmand, S.; Nair, N.; Whiting, C.C.; Enstrom, A.; Lemmens, E.; et al. Clinical Response of Live-Attenuated, Listeria monocytogenes Expressing Mesothelin (CRS-207) with Chemotherapy in Patients with Malignant Pleural MesotheliomaCRS-207 with Chemotherapy for Treatment of Mesothelioma. Clin. Cancer Res. 2019, 25, 5787–5798. [Google Scholar] [CrossRef] [PubMed]
- Lester, J.F.; Casbard, A.C.; Al-Taei, S.; Harrop, R.; Katona, L.; Attanoos, R.L.; Tabi, Z.; Griffiths, G.O. A single centre phase II trial to assess the immunological activity of TroVax® plus pemetrexed/cisplatin in patients with malignant pleural mesothelioma–the SKOPOS trial. Oncoimmunology 2018, 7, e1457597. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Kashiwagi, S.; Reeves, P.; Nezivar, J.; Yang, Y.; Arrifin, N.H.; Nguyen, M.; Jean-Mary, G.; Tong, X.; Uppal, P.; et al. A novel mycobacterial Hsp70-containing fusion protein targeting mesothelin augments antitumor immunity and prolongs survival in murine models of ovarian cancer and mesothelioma. J. Hematol. Oncol. 2014, 7, 15. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.L.; Chang, M.C.; Chiang, Y.C.; Lin, H.W.; Sun, N.Y.; Chen, C.A.; Sun, W.Z.; Cheng, W.F. Immuno-modulators enhance antigen-specific immunity and anti-tumor effects of mesothelin-specific chimeric DNA vaccine through promoting DC maturation. Cancer Lett. 2018, 425, 152–163. [Google Scholar] [CrossRef]
- Hoffmann, P.R.; Hoffmann, F.W.; Premeaux, T.A.; Fujita, T.; Soprana, E.; Panigada, M.; Chew, G.M.; Richard, G.; Hindocha, P.; Menor, M.; et al. Multi-antigen Vaccination with Simultaneous Engagement of the OX40 Receptor Delays Malignant Mesothelioma Growth and Increases Survival in Animal Models. Front. Oncol. 2019, 9, 720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Chen, J.; Xu, Q. Current Developments and Challenges of mRNA Vaccines. Annu. Rev. Biomed. Eng. 2022, 24, 85–109. [Google Scholar] [CrossRef]
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. mRNA vaccines-a new era in vaccinology. Nat. Rev. Drug Discov. 2018, 17, 261–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayour, E.J.; Mendez-Gomez, H.R.; Mitchell, D.A. Cancer Vaccine Immunotherapy with RNA-Loaded Liposomes. Int. J. Mol. Sci. 2018, 19, 2890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khodadous, M.S.; Alizadeh, A.A. Tumor Antigen Discovery Through Translation of the Cancer Genome. Immunol. Res. 2014, 58, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Hieronymus, H.; Murali, R.; Tin, A.; Yadav, K.; Abida, W.; Moller, H.; Berney, D.; Scher, H.; Carver, B.; Scardino, P.; et al. Tumor copy number alteration burden is a pan-cancer prognostic factor associated with recurrence and death. eLife 2018, 7, e37294. [Google Scholar] [CrossRef] [PubMed]
- Yoshihara, K.; Shahmoradgoli, M.; Martínez, E.; Vegesna, R.; Kim, H.; Torres-Garcia, W.; Verhaak, R.G. Inferring tumour purity and stromal and immune cell admixture from expression data. Nat. Commun. 2013, 4, 2612. [Google Scholar] [CrossRef] [PubMed]
- Charoentong, P.; Finotello, F.; Angelova, M.; Mayer, C.; Efremova, M.; Rieder, D.; Hackl, H.; Trajanoski, Z. Pan-cancer Immunogenomic Analyses Reveal Genotype-Immunophenotype Relationships and Predictors of Response to Checkpoint Blockade. Cell Rep. 2017, 18, 248–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, Y.R.; Xia, M.; Luo, M.; Luo, T.; Yang, M.; Guo, A.Y. ImmuCellAI-mouse: A tool for comprehensive prediction of mouse immune cell abundance and immune microenvironment depiction. Bioinformatics 2021, 38, 785–791. [Google Scholar] [CrossRef] [PubMed]
- Thorsson, V.; Gibbs, D.L.; Brown, S.D.; Wolf, D.; Bortone, D.S.; Ou Yang, T.-H.; Porta-Pardo, E.; Gao, G.F.; Plaisier, C.L.; Eddy, J.A.; et al. The Immune Landscape of Cancer. Immunity 2018, 48, 812–830.e14. [Google Scholar] [CrossRef] [Green Version]
- Mi, H.; Muruganujan, A.; Ebert, D.; Huang, X.; Thomas, P.D. PANTHER version 14: More genomes, a new PANTHER GO-slim and improvements in enrichment analysis tools. Nucleic Acids Res. 2019, 47, D419–D426. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Kawashima, M. KEGG mapping tools for uncovering hidden features in biological data. Protein Sci. 2021, 31, 47–53. [Google Scholar] [CrossRef]
- Chin, C.-H.; Chen, S.-H.; Wu, H.-H.; Ho, C.-W.; Ko, M.-T.; Lin, C.-Y. cytoHubba: Identifying hub objects and sub-networks from complex interactome. BMC Syst. Biol. 2014, 8, S11. [Google Scholar] [CrossRef] [Green Version]
- Sha, D.; Jin, Z.; Budczies, J.; Kluck, K.; Stenzinger, A.; Sinicrope, F.A. Tumor Mutational Burden as a Predictive Biomarker in Solid Tumors. Cancer Discov. 2020, 10, 1808–1825. [Google Scholar] [CrossRef]
- Huang, X.; Zhang, G.; Tang, T.; Liang, T. Identification of tumor antigens and immune subtypes of pancreatic adenocarcinoma for mRNA vaccine development. Mol. Cancer 2021, 20, 44. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Tang, T.; Zhang, G.; Liang, T. Identification of tumor antigens and immune subtypes of cholangiocarcinoma for mRNA vaccine development. Mol. Cancer 2021, 20, 50. [Google Scholar] [CrossRef]
- Zhu, L.; Wang, X.; Wang, Y. Roles of FAM134B in diseases from the perspectives of organelle membrane morphogenesis and cellular homeostasis. J. Cell. Physiol. 2021, 236, 7242–7255. [Google Scholar] [CrossRef] [PubMed]
- Mo, J.; Chen, J.; Zhang, B. Critical roles of FAM134B in ER-phagy and diseases. Cell Death Dis. 2020, 11, 983. [Google Scholar] [CrossRef] [PubMed]
- Kasem, K.; Gopalan, V.; Salajegheh, A.; Lu, C.-T.; Smith, R.A.; Lam, A.K.-Y. The roles of JK-1 (FAM134B) expressions in colorectal cancer. Exp. Cell Res. 2014, 326, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Islam, F.; Gopalan, V.; Pillai, S.; Lu, C.-T.; Kasem, K.; Lam, A.K.-Y. Promoter hypermethylation inactivate tumor suppressor FAM134B and is associated with poor prognosis in colorectal cancer. Genes Chromosom. Cancer 2018, 57, 240–251. [Google Scholar] [CrossRef] [Green Version]
- Islam, F.; Chaousis, S.; Wahab, R.; Gopalan, V.; Lam, A.K. Protein interactions of FAM134B with EB1 and APC/beta-catenin in vitro in colon carcinoma. Mol. Carcinog. 2018, 57, 1480–1491. [Google Scholar] [CrossRef]
- Amr, K.; El-Bassyouni, H.T.; Ismail, S.; Youness, E.; El-Daly, S.M.; Ebrahim, A.Y.; El-Kamah, G. Genetic assessment of ten Egyptian patients with Sjögren–Larsson syndrome: Expanding the clinical spectrum and reporting a novel ALDH3A2 mutation. Arch. Dermatol. Res. 2019, 311, 721–730. [Google Scholar] [CrossRef]
- Yin, Z.; Wu, D.; Shi, J.; Wei, X.; Jin, N.; Lu, X.; Ren, X. Identification of ALDH3A2 as a novel prognostic biomarker in gastric adenocarcinoma using integrated bioinformatics analysis. BMC Cancer 2020, 20, 1062. [Google Scholar]
- Ehmer, U.; Sage, J. Control of Proliferation and Cancer Growth by the Hippo Signaling Pathway. Mol. Cancer Res. 2016, 14, 127–140. [Google Scholar] [CrossRef] [Green Version]
- de Amorim, Í.S.S.; de Sousa Rodrigues, M.M.; Mencalha, A.L. The tumor suppressor role of salvador family WW domain-containing protein 1 (SAV1): One of the key pieces of the tumor puzzle. J. Cancer Res. Clin. Oncol. 2021, 147, 1287–1297. [Google Scholar] [CrossRef] [PubMed]
- Oh, T.G.; Wang, S.-C.; Acharya, B.R.; Goode, J.M.; Graham, J.D.; Clarke, C.L.; Yap, A.; Muscat, G.E. The Nuclear Receptor, RORγ, Regulates Pathways Necessary for Breast Cancer Metastasis. eBioMedicine 2016, 6, 59–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.X.; Liu, H.; Xu, L.; Zhang, H.; Zhou, R.X. Melatonin downregulates nuclear receptor RZR/RORgamma expression causing growth-inhibitory and anti-angiogenesis activity in human gastric cancer cells in vitro and in vivo. Oncol. Lett. 2016, 12, 897–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brozyna, A.A.; Jozwicki, W.; Skobowiat, C.; Jetten, A.; Slominski, A.T. RORalpha and RORgamma expression inversely correlates with human melanoma progression. Oncotarget 2016, 7, 63261–63282. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.; Qi, Z.; Pang, Y.; Li, H.; Xie, H.; Wu, J.; Wang, Z. Retinoic Acid–Related Orphan Receptor C Regulates Proliferation, Glycolysis, and Chemoresistance via the PD-L1/ITGB6/STAT3 Signaling Axis in Bladder CancerThe RORC/PD-L1/ITGB6/STAT3 Signaling Axis in Bladder Cancer. Cancer Res. 2019, 79, 2604–2618. [Google Scholar] [CrossRef] [Green Version]
- Cai, X.; Liu, C.; Zhang, T.N.; Zhu, Y.W.; Dong, X.; Xue, P. Down-regulation of FN1 inhibits colorectal carcinogenesis by suppressing proliferation, migration, and invasion. J. Cell Biochem. 2018, 119, 4717–4728. [Google Scholar] [CrossRef] [PubMed]
- Qin, Z.; Zhou, C. HOXA13 promotes gastric cancer progression partially via the FN1-mediated FAK/Src axis. Exp. Hematol. Oncol. 2022, 11, 7. [Google Scholar] [CrossRef]
- Xu, X.; Cheng, L.; Fan, Y.; Mao, W. Tumor Microenvironment-Associated Immune-Related Genes for the Prognosis of Malignant Pleural Mesothelioma. Front. Oncol. 2020, 10, 544789. [Google Scholar] [CrossRef]
- Fuso Nerini, I.; Roca, E.; Mannarino, L.; Grosso, F.; Frapolli, R.; D’Incalci, M. Is DNA repair a potential target for effective therapies against malignant mesothelioma? Cancer Treat. Rev. 2020, 90, 102101. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Yang, Y.; Li, L.; Ma, P.; Jiang, Y.; Ge, M.; Yu, Y.; Huang, H.; Fang, Y.; Jiang, N.; et al. Identification of Tumor Antigens and Immune Subtypes of Malignant Mesothelioma for mRNA Vaccine Development. Vaccines 2022, 10, 1168. https://doi.org/10.3390/vaccines10081168
Wang S, Yang Y, Li L, Ma P, Jiang Y, Ge M, Yu Y, Huang H, Fang Y, Jiang N, et al. Identification of Tumor Antigens and Immune Subtypes of Malignant Mesothelioma for mRNA Vaccine Development. Vaccines. 2022; 10(8):1168. https://doi.org/10.3390/vaccines10081168
Chicago/Turabian StyleWang, Shuhang, Yuqi Yang, Lu Li, Peiwen Ma, Yale Jiang, Minghui Ge, Yue Yu, Huiyao Huang, Yuan Fang, Ning Jiang, and et al. 2022. "Identification of Tumor Antigens and Immune Subtypes of Malignant Mesothelioma for mRNA Vaccine Development" Vaccines 10, no. 8: 1168. https://doi.org/10.3390/vaccines10081168
APA StyleWang, S., Yang, Y., Li, L., Ma, P., Jiang, Y., Ge, M., Yu, Y., Huang, H., Fang, Y., Jiang, N., Miao, H., Guo, H., Yan, L., Ren, Y., Sun, L., Zha, Y., & Li, N. (2022). Identification of Tumor Antigens and Immune Subtypes of Malignant Mesothelioma for mRNA Vaccine Development. Vaccines, 10(8), 1168. https://doi.org/10.3390/vaccines10081168