
Influenza Virus-like Particle-Based Hybrid Vaccine Containing RBD Induces Immunity against Influenza and SARS-CoV-2 Viruses

 ,
,  , ,
, ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Antibodies and Proteins
2.2. Mice
2.3. Construction and Expression of GPI-Anchored SARS-CoV-2 RBD-GM-CSF Fusion Protein
2.4. PIPLC Treatment of CHO-S Cells
2.5. Purification of GPI-RBD-GM-CSF and GPI-IL-12
2.6. SDS-PAGE and Western Blot
2.7. Hybrid Vaccine Preparation by Protein Transfer of GPI-RBD-GM-CSF Fusion Protein and GPI-IL-12 onto VLP
2.8. Bone Marrow-Derived Dendritic Cell Stimulation Assay
2.9. Immunization with VLP Vaccine
2.10. Enzyme-Linked Immunosorbent Assay (ELISA)
2.11. Flow Cytometry Analysis of Antibody Response
2.12. Focus Reduction Neutralization Assay
2.13. Plaque Reduction Neutralization Test (PRNT)
2.14. Hemagglutination Inhibition (HAI) Assay
2.15. SARS-CoV-2 Virus Challenge
2.16. Quantification of the SARS-CoV-2 Virus Load in the Lungs
2.17. Influenza A/PR8 H1N1 Virus Challenge
2.18. Influenza Virus Titration in the Lung
2.19. Statistical Analysis
3. Results
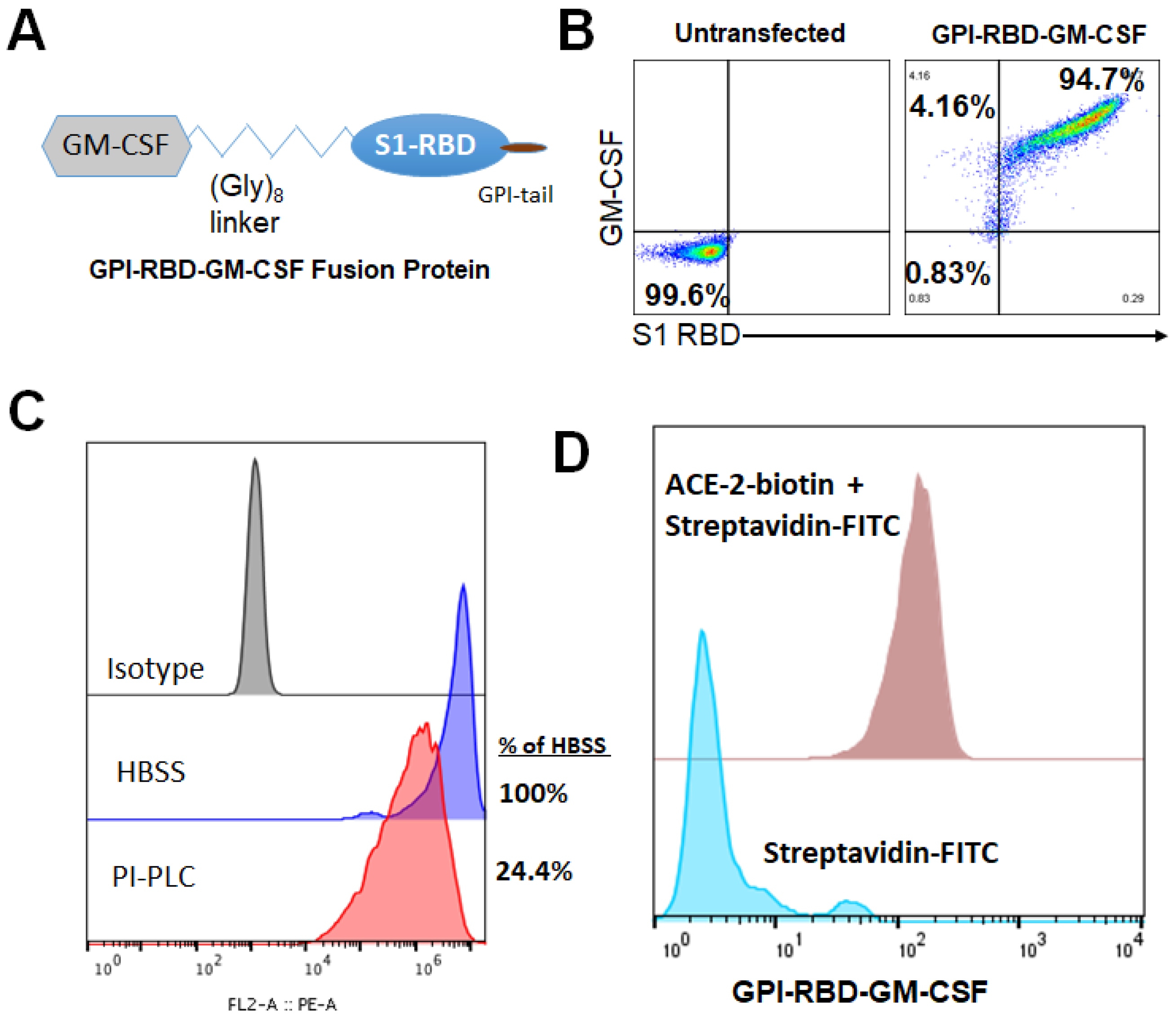
3.1. Characterization of GPI-RBD-GM-CSF Fusion Protein
3.2. GPI-RBD-GM-CSF Fusion Protein Retains Functional Activity
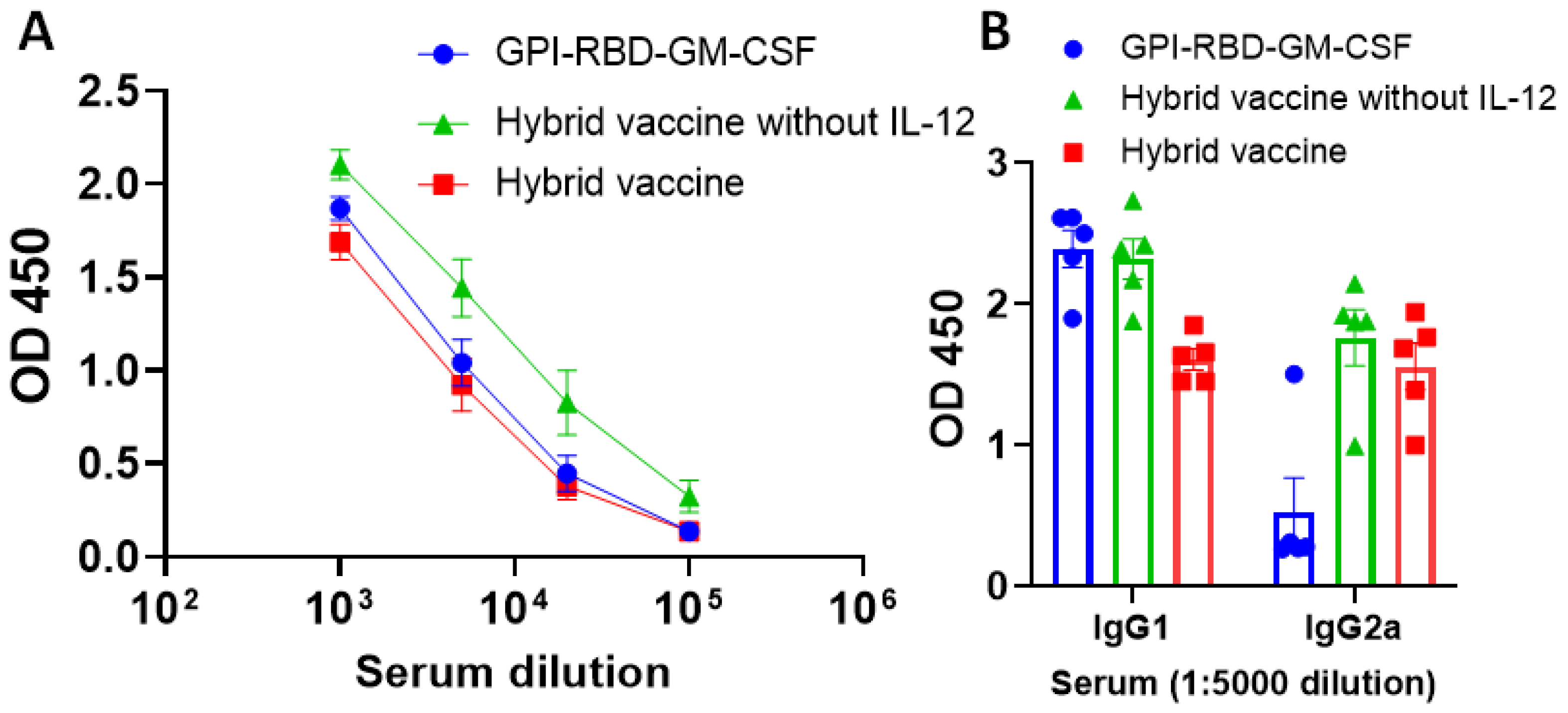
3.3. Hybrid Vaccine Induces Durable Antibody Response
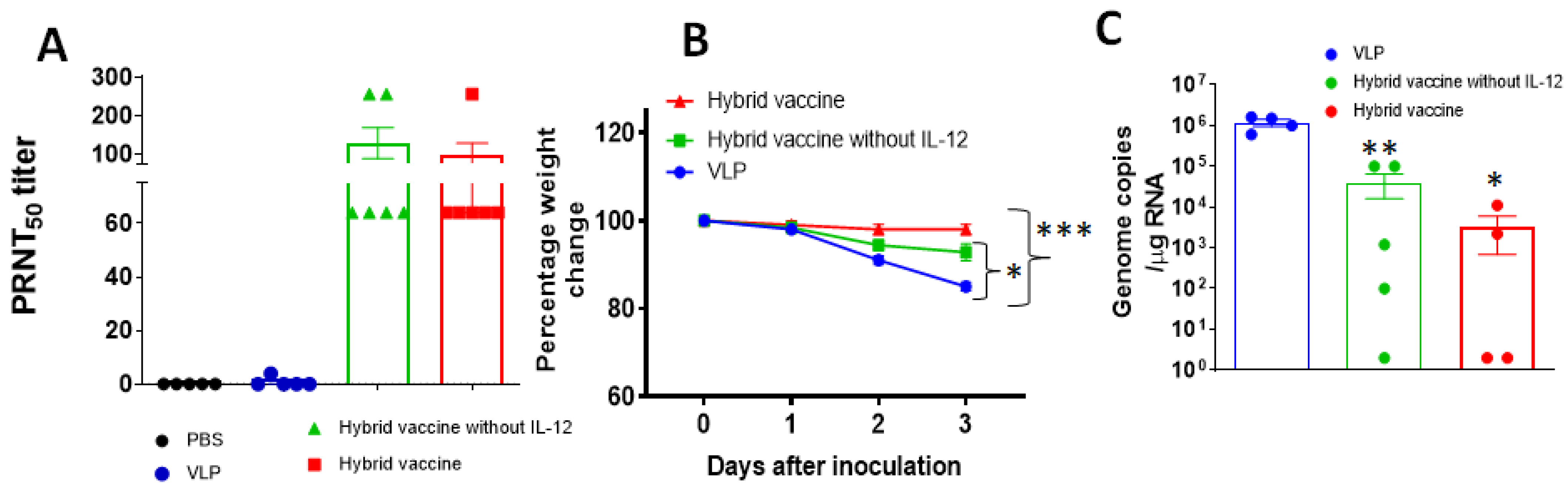
3.4. Hybrid Vaccine Induces SARS-CoV-2 Neutralizing Antibodies
3.5. Hybrid Vaccine Induces IgG2a Antibody Response against RBD
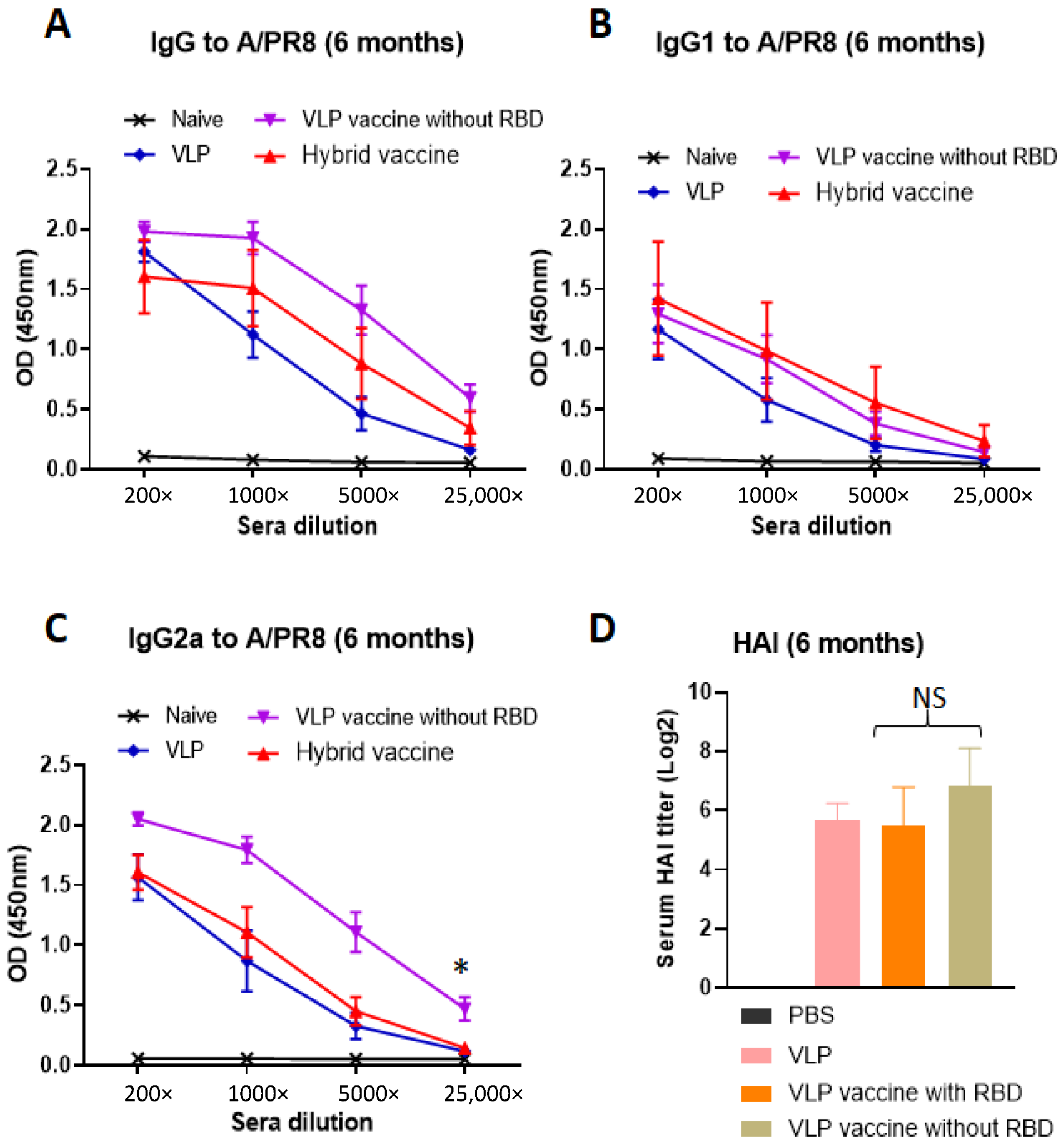
3.6. Hybrid Vaccine Induces Anti-Influenza Virus IgG1 and IgG2a Antibody Response
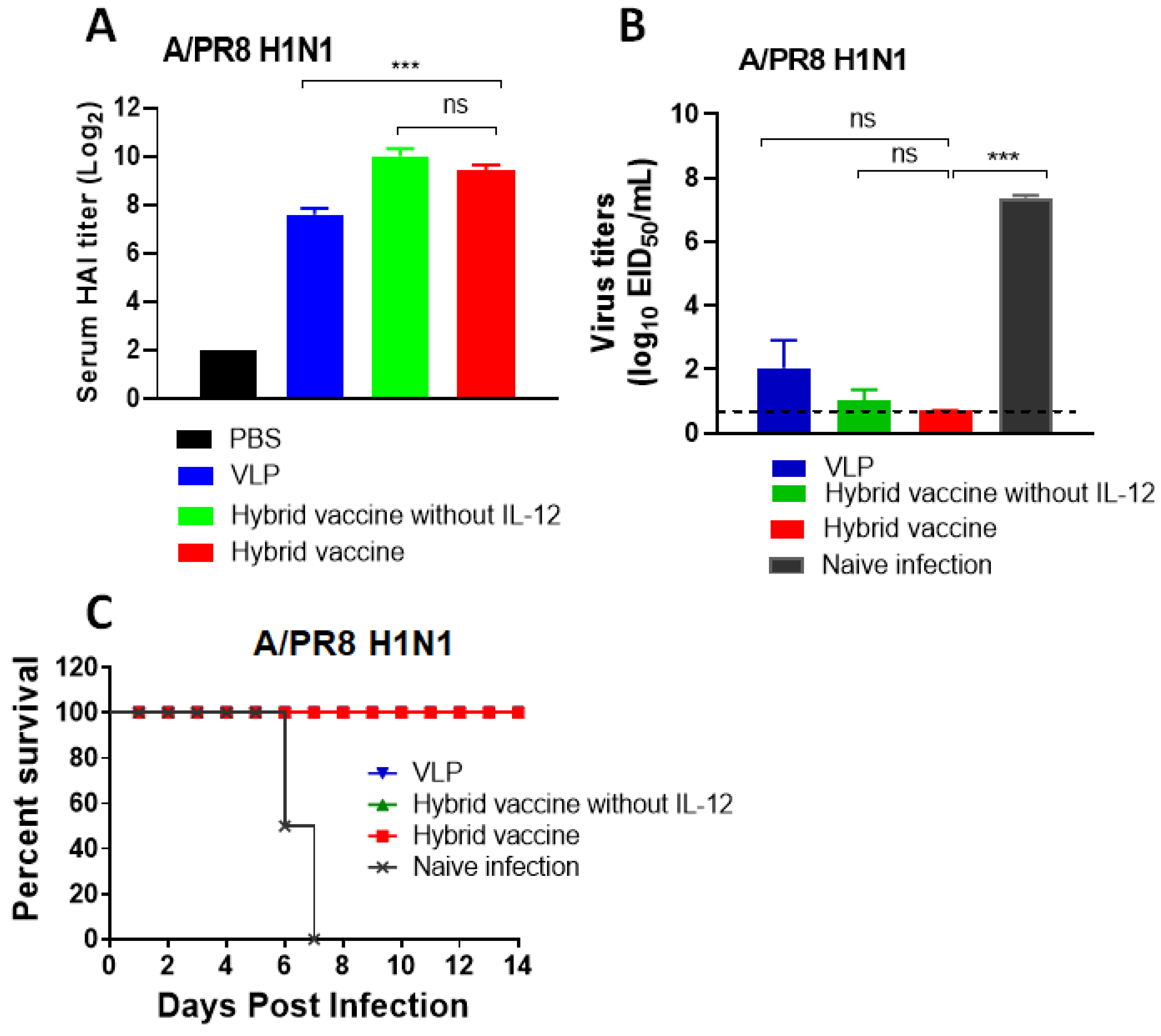
3.7. Hybrid Vaccine Protects Mice from SARS-CoV-2 and Influenza Virus Challenges
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [PubMed]
- Sandor, A.M.; Sturdivant, M.S.; Ting, J.P.Y. Influenza Virus and SARS-CoV-2 Vaccines. J. Immunol 2021, 206, 2509–2520. [Google Scholar] [CrossRef] [PubMed]
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L.; et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature 2020, 581, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Yong, C.Y.; Ong, H.K.; Yeap, S.K.; Ho, K.L.; Tan, W.S. Recent Advances in the Vaccine Development Against Middle East Respiratory Syndrome-Coronavirus. Front. Microbiol. 2019, 10, 1781. [Google Scholar] [CrossRef]
- Amanat, F.; Krammer, F. SARS-CoV-2 Vaccines: Status Report. Immunity 2020, 52, 583–589. [Google Scholar] [CrossRef]
- Prates-Syed, W.A.; Chaves, L.C.S.; Crema, K.P.; Vuitika, L.; Lira, A.; Cortes, N.; Kersten, V.; Guimaraes, F.E.G.; Sadraeian, M.; Barroso da Silva, F.L.; et al. VLP-Based COVID-19 Vaccines: An Adaptable Technology against the Threat of New Variants. Vaccines 2021, 9, 1409. [Google Scholar] [CrossRef]
- Ella, R.; Reddy, S.; Jogdand, H.; Sarangi, V.; Ganneru, B.; Prasad, S.; Das, D.; Raju, D.; Praturi, U.; Sapkal, G.; et al. Safety and immunogenicity of an inactivated SARS-CoV-2 vaccine, BBV152: Interim results from a double-blind, randomised, multicentre, phase 2 trial, and 3-month follow-up of a double-blind, randomised phase 1 trial. Lancet Infect. Dis. 2021, 21, 950–961. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, Y.; Huang, B.; Deng, W.; Quan, Y.; Wang, W.; Xu, W.; Zhao, Y.; Li, N.; Zhang, J.; et al. Development of an Inactivated Vaccine Candidate, BBIBP-CorV, with Potent Protection against SARS-CoV-2. Cell 2020, 182, 713–721.e9. [Google Scholar] [CrossRef]
- Bao, L.; Deng, W.; Qi, F.; Lv, Q.; Song, Z.; Liu, J.; Gao, H.; Wei, Q.; Yu, P.; Xu, Y.; et al. Sequential infection with H1N1 and SARS-CoV-2 aggravated COVID-19 pathogenesis in a mammalian model, and co-vaccination as an effective method of prevention of COVID-19 and influenza. Signal. Transduct Target. 2021, 6, 200. [Google Scholar] [CrossRef]
- Zhou, R.; Wang, P.; Wong, Y.C.; Xu, H.; Lau, S.Y.; Liu, L.; Mok, B.W.; Peng, Q.; Liu, N.; Woo, K.F.; et al. Nasal prevention of SARS-CoV-2 infection by intranasal influenza-based boost vaccination in mouse models. EBioMedicine 2022, 75, 103762. [Google Scholar] [CrossRef]
- Toback, S.; Galiza, E.; Cosgrove, C.; Galloway, J.; Goodman, A.L.; Swift, P.A.; Rajaram, S.; Graves-Jones, A.; Edelman, J.; Burns, F.; et al. Safety, immunogenicity, and efficacy of a COVID-19 vaccine (NVX-CoV2373) co-administered with seasonal influenza vaccines: An exploratory substudy of a randomised, observer-blinded, placebo-controlled, phase 3 trial. Lancet Respir. Med. 2022, 10, 167–179. [Google Scholar] [CrossRef]
- Patel, J.M.; Kim, M.C.; Vartabedian, V.F.; Lee, Y.N.; He, S.; Song, J.M.; Choi, H.J.; Yamanaka, S.; Amaram, N.; Lukacher, A.; et al. Protein transfer-mediated surface engineering to adjuvantate virus-like nanoparticles for enhanced anti-viral immune responses. Nanomed. Nanotechnol. Biol. Med. 2015, 11, 1097–1107. [Google Scholar] [CrossRef] [PubMed]
- Fallon, J.; Tighe, R.; Kradjian, G.; Guzman, W.; Bernhardt, A.; Neuteboom, B.; Lan, Y.; Sabzevari, H.; Schlom, J.; Greiner, J.W. The immunocytokine NHS-IL12 as a potential cancer therapeutic. Oncotarget 2014, 5, 1869–1884. [Google Scholar] [CrossRef] [PubMed]
- Metzger, D.W. IL-12 as an adjuvant for the enhancement of protective humoral immunity. Expert Rev. Vaccines 2009, 8, 515–518. [Google Scholar] [CrossRef] [PubMed]
- Dranoff, G. GM-CSF-secreting melanoma vaccines. Oncogene 2003, 22, 3188–3192. [Google Scholar] [CrossRef] [PubMed]
- Chiodoni, C.; Paglia, P.; Stoppacciaro, A.; Rodolfo, M.; Parenza, M.; Colombo, M. Dendritic cells infiltrating tumors cotransduced with granulocyte/macrophage colony-stimulating factor (GMCSF) and CD40 ligand genes take up and present endogenous tumor-associated antigens, and prime naive mice for a cytotoxic T lymphocyte response. J. Exp. Med. 1999, 190, 125–133. [Google Scholar] [CrossRef]
- Skountzou, I.; Quan, F.S.; Gangadhara, S.; Ye, L.; Vzorov, A.; Selvaraj, P.; Jacob, J.; Compans, R.W.; Kang, S.M. Incorporation of glycosylphosphatidylinositol-anchored GM-CSF or CD40 ligand enhamces immunogenicity of chimeric simian immunodeficiency virus-like particles. J. Virol. 2007, 81, 1083–1094. [Google Scholar] [CrossRef]
- Ou-Yang, P.; Hwang, L.H.; Tao, M.H.; Chiang, B.L.; Chen, D.S. Co-delivery of GM-CSF gene enhances the immune responses of hepatitis C viral core protein-expressing DNA vaccine: Role of dendritic cells. J. Med. Virol. 2002, 66, 320–328. [Google Scholar] [CrossRef]
- Mach, N.; Gillessen, S.; Wilson, S.B.; Sheehan, C.; Mihm, M.; Dranoff, G. Differences in dendritic cells stimulated in vivo by tumors engineered to secrete granulocyte-macrophage colony-stimulating factor or Flt3-ligand. Cancer Res. 2000, 60, 3239–3246. [Google Scholar]
- Poloso, N.J.; Nagarajan, S.; Mejia-Oneta, J.M.; Selvaraj, P. GPI-anchoring of GM-CSF results in active membrane-bound and partially shed cytokine. Mol. Immunol. 2002, 38, 803–816. [Google Scholar] [CrossRef]
- Kantoff, P.W.; Higano, C.S.; Shore, N.D.; Berger, E.R.; Small, E.J.; Penson, D.F.; Redfern, C.H.; Ferrari, A.C.; Dreicer, R.; Sims, R.B.; et al. Sipuleucel-T immunotherapy for castration-resistant prostate cancer. N. Engl. J. Med. 2010, 363, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Higano, C.S.; Schellhammer, P.F.; Small, E.J.; Burch, P.A.; Nemunaitis, J.; Yuh, L.; Provost, N.; Frohlich, M.W. Integrated data from 2 randomized, double-blind, placebo-controlled, phase 3 trials of active cellular immunotherapy with sipuleucel-T in advanced prostate cancer. Cancer 2009, 115, 3670–3679. [Google Scholar] [CrossRef] [PubMed]
- Blanco, P.; Palucka, A.K.; Pascual, V.; Banchereau, J. Dendritic cells and cytokines in human inflammatory and autoimmune diseases. Cytokine Growth Factor Rev. 2008, 19, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Brahmakshatriya, V.; Swain, S.L. CD4 T cell defects in the aged: Causes, consequences and strategies to circumvent. Exp. Gerontol. 2014, 54, 67–70. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Jaffee, E.M. Immunotherapy of cancer. Ann. Ny. Acad. Sci. 1999, 886, 67–72. [Google Scholar] [CrossRef]
- Trinchieri, G.; Scott, P. Interleukin-12: Basic principles and clinical applications. Redirection Th1 Th2 Responses 1999, 238, 57–78. [Google Scholar]
- Vagliani, M.; Rodolfo, M.; Cavallo, F.; Parenza, M.; Melani, C.; Parmiani, G.; Forni, G.; Colombo, M.P. Interleukin 12 potentiates the curative effect of a vaccine based on interleukin 2-transduced tumor Cells. Cancer Res. 1996, 56, 467–470. [Google Scholar]
- Trinchieri, G. Interleukin-12: A cytokine produced by antigen-presenting cells with immunoregulatory functions in the generation of T-helper cells type 1 and cytotoxic lymphocytes. Blood 1994, 84, 4008–4027. [Google Scholar] [CrossRef]
- Car, B.D.; Eng, V.M.; Lipman, J.M.; Anderson, T.D. The toxicology of interleukin-12: A review. Toxicol. Pathol. 1999, 27, 58–63. [Google Scholar] [CrossRef]
- Bajetta, E.; Del Vecchio, M.; Mortarini, R.; Nadeau, R.; Rakhit, A.; Rimassa, L.; Fowst, C.; Borri, A.; Anichini, A.; Parmiani, G. Pilot study of subcutaneous recombinant human interleukin 12 in metastatic melanoma. Clin. Cancer Res. 1998, 4, 75–85. [Google Scholar]
- Brunda, M.J.; Luistro, L.; Rumennik, L.; Wright, R.B.; Dvorozniak, M.; Aglione, A.; Wigginton, J.M.; Wiltrout, R.H.; Hendrzak, J.A.; Palleroni, A.V. Antitumor activity of interleukin 12 in preclinical models. Cancer Chemother. Pharmacol. 1996, 38, S16–S21. [Google Scholar] [CrossRef]
- Chen, L.; Chen, D.; Block, E.; O’Donnell, M.; Kufe, D.W.; Clinton, S.K. Eradication of murine bladder carcinoma by intratumor injection of a bicistronic adenoviral vector carrying cDNAs for the IL-12 heterodimer and its inhibition by the IL-12 p40 subunit homodimer. J. Immunol 1997, 159, 351–359. [Google Scholar] [PubMed]
- Tsung, K.; Meko, J.B.; Tsung, Y.L.; Peplinski, G.R.; Norton, J.A. Immune response against large tumors eradicated by treatment with cyclophosphamide and IL-12. J. Immunol 1998, 160, 1369–1377. [Google Scholar] [PubMed]
- Cohen, J. IL-12 deaths: Explanation and a puzzle. Science 1995, 270, 908. [Google Scholar] [CrossRef] [PubMed]
- Villarino, A.; Hibbert, L.; Lieberman, L.; Wilson, E.; Mak, T.; Yoshida, H.; Kastelein, R.A.; Saris, C.; Hunter, C.A. The IL-27R (WSX-1) is required to suppress T cell hyperactivity during infection. Immunity 2003, 19, 645–655. [Google Scholar] [CrossRef]
- Zhang, L.; Davies, J.S.; Serna, C.; Yu, Z.; Restifo, N.P.; Rosenberg, S.A.; Morgan, R.A.; Hinrichs, C.S. Enhanced efficacy and limited systemic cytokine exposure with membrane-anchored interleukin-12 T-cell therapy in murine tumor models. J. Immunother. Cancer 2020, 8, e000210. [Google Scholar] [CrossRef]
- Pan, W.Y.; Lo, C.H.; Chen, C.C.; Wu, P.Y.; Roffler, S.R.; Shyue, S.K.; Tao, M.H. Cancer immunotherapy using a membrane-bound interleukin-12 with B7-1 transmembrane and cytoplasmic domains. Mol. Ther. 2012, 20, 927–937. [Google Scholar] [CrossRef]
- Maegawa, K.; Sugita, S.; Arasaki, Y.; Nerome, R.; Nerome, K. Interleukin 12-containing influenza virus-like-particle vaccine elevate its protective activity against heterotypic influenza virus infection. Heliyon 2020, 6, e04543. [Google Scholar] [CrossRef]
- Selvaraj, P.; Yerra, A.; Tein, L.; Shashidharamurthy, R. Custom designing therapeutic cancer vaccines: Delivery of immunostimulatory molecule adjuvants by protein transfer. Hum. Vaccines 2008, 4, 384–388. [Google Scholar] [CrossRef]
- Patel, J.M.; Vartabedian, V.F.; Kim, M.C.; He, S.; Kang, S.M.; Selvaraj, P. Influenza virus-like particles engineered by protein transfer with tumor-associated antigens induces protective antitumor immunity. Biotechnol. Bioeng. 2015, 112, 1102–1110. [Google Scholar] [CrossRef]
- Huleatt, J.W.; Nakaar, V.; Desai, P.; Huang, Y.; Hewitt, D.; Jacobs, A.; Tang, J.; McDonald, W.; Song, L.; Evans, R.K.; et al. Potent immunogenicity and efficacy of a universal influenza vaccine candidate comprising a recombinant fusion protein linking influenza M2e to the TLR5 ligand flagellin. Vaccine 2008, 26, 201–214. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, D.M.; Fausch, S.C.; Verbeek, J.S.; Kast, W.M. Uptake of human papillomavirus virus-like particles by dendritic cells is mediated by Fcgamma receptors and contributes to acquisition of T cell immunity. J. Immunol. 2007, 178, 7587–7597. [Google Scholar] [CrossRef] [PubMed]
- Patel, J.M.; Vartabedian, V.F.; Bozeman, E.N.; Caoyonan, B.E.; Srivatsan, S.; Pack, C.D.; Dey, P.; D Souza, M.J.; Yang, L.; Selvaraj, P. Plasma membrane vesicles decorated with glycolipid-anchored antigens and adjuvants via protein transfer as an antigen delivery platform for inhibition of tumor growth. Biomaterials 2015, 74, 231–244. [Google Scholar] [CrossRef] [PubMed]
- Cohen, A.A.; Gnanapragasam, P.N.P.; Lee, Y.E.; Hoffman, P.R.; Ou, S.; Kakutani, L.M.; Keeffe, J.R.; Wu, H.-J.; Howarth, M.; West, A.P.; et al. Mosaic nanoparticles elicit cross-reactive immune responses to zoonotic coronaviruses in mice. Science 2021, 371, 735–741. [Google Scholar] [CrossRef]
- Lutz, M.; Kukutsch, N.; Ogilvie, A.; RoBner, S.; Koch, F.; Romani, N.; Schuler, G. An advanced culture method for generating large quantities of highly pure dendritic cells from mouse bone marrow. J. Immunol. Methods 1999, 223, 77–92. [Google Scholar] [CrossRef]
- Quan, F.S.; Ko, E.J.; Kwon, Y.M.; Joo, K.H.; Compans, R.W.; Kang, S.M. Mucosal adjuvants for influenza virus-like particle vaccine. Viral Immunol. 2013, 26, 385–395. [Google Scholar] [CrossRef]
- Quan, F.S.; Huang, C.; Compans, R.W.; Kang, S.M. Virus-like particle vaccine induces protective immunity against homologous and heterologous strains of influenza virus. J. Virol. 2007, 81, 3514–3524. [Google Scholar] [CrossRef]
- Xie, X.; Muruato, A.; Lokugamage, K.G.; Narayanan, K.; Zhang, X.; Zou, J.; Liu, J.; Schindewolf, C.; Bopp, N.E.; Aguilar, P.V.; et al. An Infectious cDNA Clone of SARS-CoV-2. Cell Host Microbe 2020, 27, 841–848.e3. [Google Scholar] [CrossRef]
- Vanderheiden, A.; Edara, V.V.; Floyd, K.; Kauffman, R.C.; Mantus, G.; Anderson, E.; Rouphael, N.; Edupuganti, S.; Shi, P.Y.; Menachery, V.D.; et al. Development of a Rapid Focus Reduction Neutralization Test Assay for Measuring SARS-CoV-2 Neutralizing Antibodies. Curr. Protoc. Immunol. 2020, 131, e116. [Google Scholar] [CrossRef]
- Katzelnick, L.C.; Coello Escoto, A.; McElvany, B.D.; Chávez, C.; Salje, H.; Luo, W.; Rodriguez-Barraquer, I.; Jarman, R.; Durbin, A.P.; Diehl, S.A.; et al. Viridot: An automated virus plaque (immunofocus) counter for the measurement of serological neutralizing responses with application to dengue virus. PLoS Negl. Trop. Dis. 2018, 12, e0006862. [Google Scholar] [CrossRef]
- Stone, S.; Rothan, H.A.; Natekar, J.P.; Kumari, P.; Sharma, S.; Pathak, H.; Arora, K.; Auroni, T.T.; Kumar, M. SARS-CoV-2 Variants of Concern Infect the Respiratory Tract and Induce Inflammatory Response in Wild-Type Laboratory Mice. Viruses 2021, 14, 27. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Chen, C.; Drelich, A.; Martinez, D.R.; Gralinski, L.E.; Sun, Z.; Schäfer, A.; Kulkarni, S.S.; Liu, X.; Leist, S.R.; et al. Rapid identification of a human antibody with high prophylactic and therapeutic efficacy in three animal models of SARS-CoV-2 infection. Proc. Natl. Acad. Sci. USA 2020, 117, 29832–29838. [Google Scholar] [CrossRef] [PubMed]
- Leist, S.R.; Dinnon, K.H., 3rd; Schäfer, A.; Tse, L.V.; Okuda, K.; Hou, Y.J.; West, A.; Edwards, C.E.; Sanders, W.; Fritch, E.J.; et al. A Mouse-Adapted SARS-CoV-2 Induces Acute Lung Injury and Mortality in Standard Laboratory Mice. Cell 2020, 183, 1070–1085.e12. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Pardi, N.; Hogan, M.J.; Weissman, D. Recent advances in mRNA vaccine technology. Curr. Opin. Immunol. 2020, 65, 14–20. [Google Scholar] [CrossRef]
- Rashidzadeh, H.; Danafar, H.; Rahimi, H.; Mozafari, F.; Salehiabar, M.; Rahmati, M.A.; Rahamooz-Haghighi, S.; Mousazadeh, N.; Mohammadi, A.; Ertas, Y.N.; et al. Nanotechnology against the novel coronavirus (severe acute respiratory syndrome coronavirus 2): Diagnosis, treatment, therapy and future perspectives. Nanomedicine 2021, 16, 497–516. [Google Scholar] [CrossRef]
- Carvalho, S.B.; Silva, R.J.S.; Sousa, M.F.Q.; Peixoto, C.; Roldão, A.; Carrondo, M.J.T.; Alves, P.M. Bioanalytics for Influenza Virus-Like Particle Characterization and Process Monitoring. Front. Bioeng. Biotechnol. 2022, 10, 805176. [Google Scholar] [CrossRef]
- Mohsen, M.O.; Zha, L.; Cabral-Miranda, G.; Bachmann, M.F. Major findings and recent advances in virus-like particle (VLP)-based vaccines. Semin Immunol . 2017, 34, 123–132. [Google Scholar] [CrossRef]
- Kang, H.J.; Chu, K.B.; Yoon, K.W.; Eom, G.D.; Mao, J.; Kim, M.J.; Lee, S.H.; Moon, E.K.; Quan, F.S. Multiple Neuraminidase Containing Influenza Virus-like Particle Vaccines Protect Mice from Avian and Human Influenza Virus Infection. Viruses 2022, 14, 429. [Google Scholar] [CrossRef]
- Patel, J.M.; Vartabedian, V.F.; Selvaraj, P. Lipid-Mediated Cell Surface Engineering. In Micro- and Nanoengineering of the Cell Surface; Karp, J., Zhao, W., Eds.; Elsevier Inc.: Alpharetta, GA, USA, 2014; pp. 121–141. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bommireddy, R.; Stone, S.; Bhatnagar, N.; Kumari, P.; Munoz, L.E.; Oh, J.; Kim, K.-H.; Berry, J.T.L.; Jacobsen, K.M.; Jaafar, L.; et al. Influenza Virus-like Particle-Based Hybrid Vaccine Containing RBD Induces Immunity against Influenza and SARS-CoV-2 Viruses. Vaccines 2022, 10, 944. https://doi.org/10.3390/vaccines10060944
Bommireddy R, Stone S, Bhatnagar N, Kumari P, Munoz LE, Oh J, Kim K-H, Berry JTL, Jacobsen KM, Jaafar L, et al. Influenza Virus-like Particle-Based Hybrid Vaccine Containing RBD Induces Immunity against Influenza and SARS-CoV-2 Viruses. Vaccines. 2022; 10(6):944. https://doi.org/10.3390/vaccines10060944
Chicago/Turabian StyleBommireddy, Ramireddy, Shannon Stone, Noopur Bhatnagar, Pratima Kumari, Luis E. Munoz, Judy Oh, Ki-Hye Kim, Jameson T. L. Berry, Kristen M. Jacobsen, Lahcen Jaafar, and et al. 2022. "Influenza Virus-like Particle-Based Hybrid Vaccine Containing RBD Induces Immunity against Influenza and SARS-CoV-2 Viruses" Vaccines 10, no. 6: 944. https://doi.org/10.3390/vaccines10060944
APA StyleBommireddy, R., Stone, S., Bhatnagar, N., Kumari, P., Munoz, L. E., Oh, J., Kim, K.-H., Berry, J. T. L., Jacobsen, K. M., Jaafar, L., Naing, S.-H., Blackerby, A. N., Gaag, T. V. d., Wright, C. N., Lai, L., Pack, C. D., Ramachandiran, S., Suthar, M. S., Kang, S.-M., ... Selvaraj, P. (2022). Influenza Virus-like Particle-Based Hybrid Vaccine Containing RBD Induces Immunity against Influenza and SARS-CoV-2 Viruses. Vaccines, 10(6), 944. https://doi.org/10.3390/vaccines10060944