
Inactivated Whole Virus Particle Influenza Vaccine Induces Anti-Neuraminidase Antibodies That May Contribute to Cross-Protection against Heterologous Virus Infection

 , , and
, , and 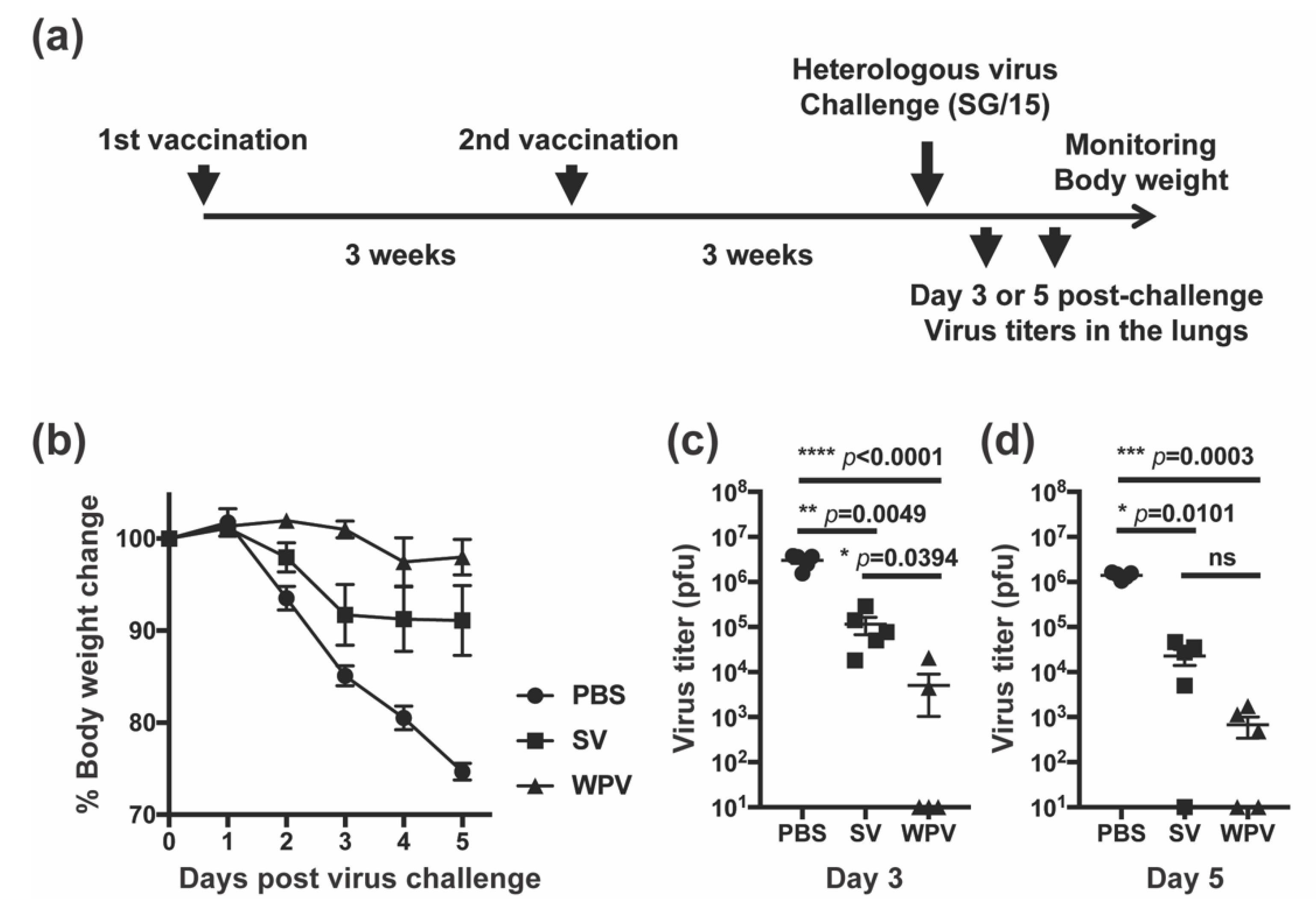{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Vaccine Preparation
2.2. Cells and Viruses
2.3. Animals
2.4. First Animal Experiment-Vaccination and Virus Challenge Study
2.5. Second Animal Experiment-Vaccination and Serum Collection
2.6. HI Assay
2.7. Neuraminidase and NI Assays
2.8. Neutralization of Viral Infectivity Test
2.9. Plaque Assay
2.10. Ethics Statement
2.11. Statistical Analysis
3. Results
3.1. Mice Vaccinated with WPV Exhibited Superior Cross-Protection Compared with Those Vaccinated with SV

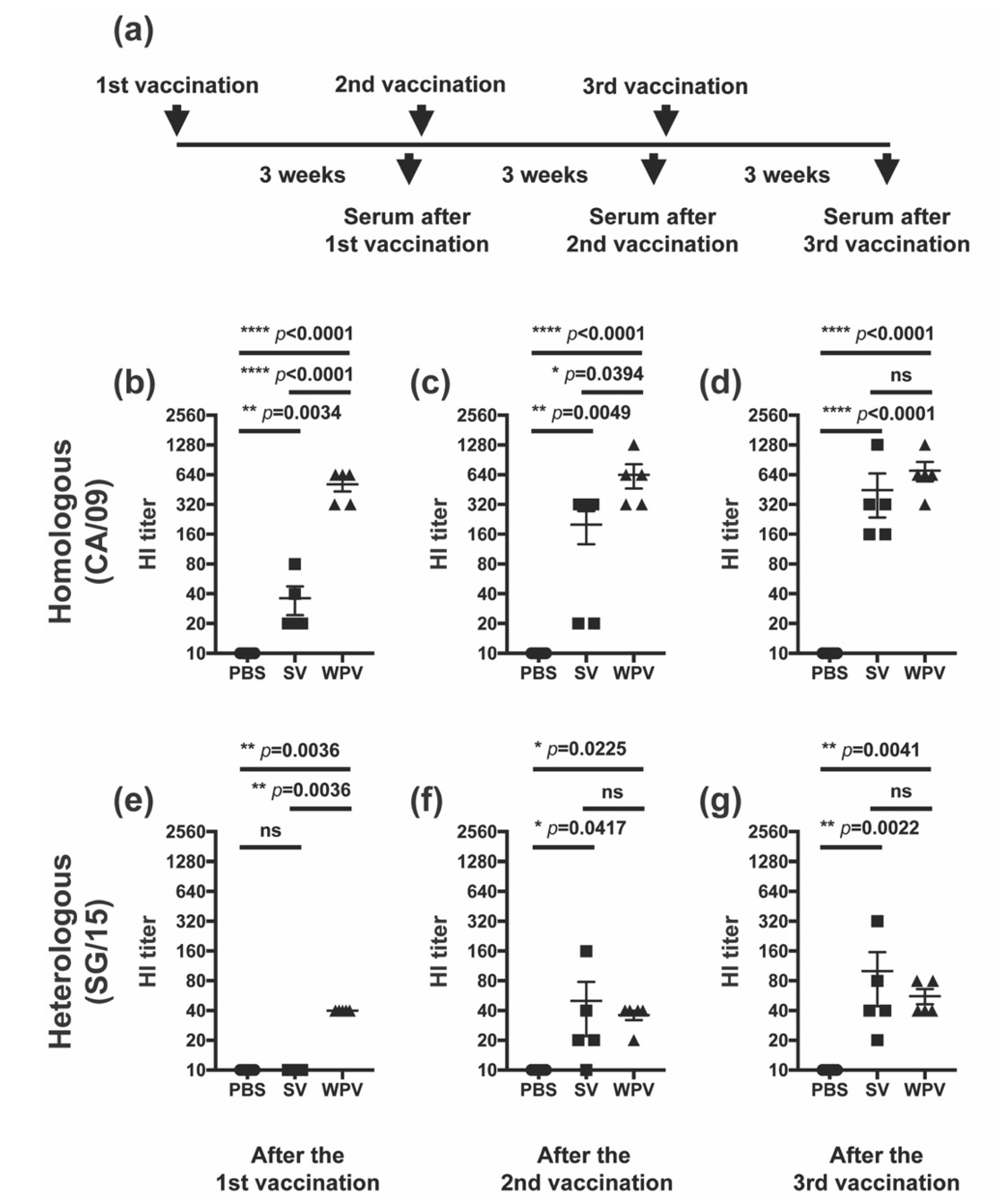
3.2. After Multiple Vaccinations, Mice Inoculated with SV Induced HI Antibodies Comparable to Those of Mice Vaccinated with WPV

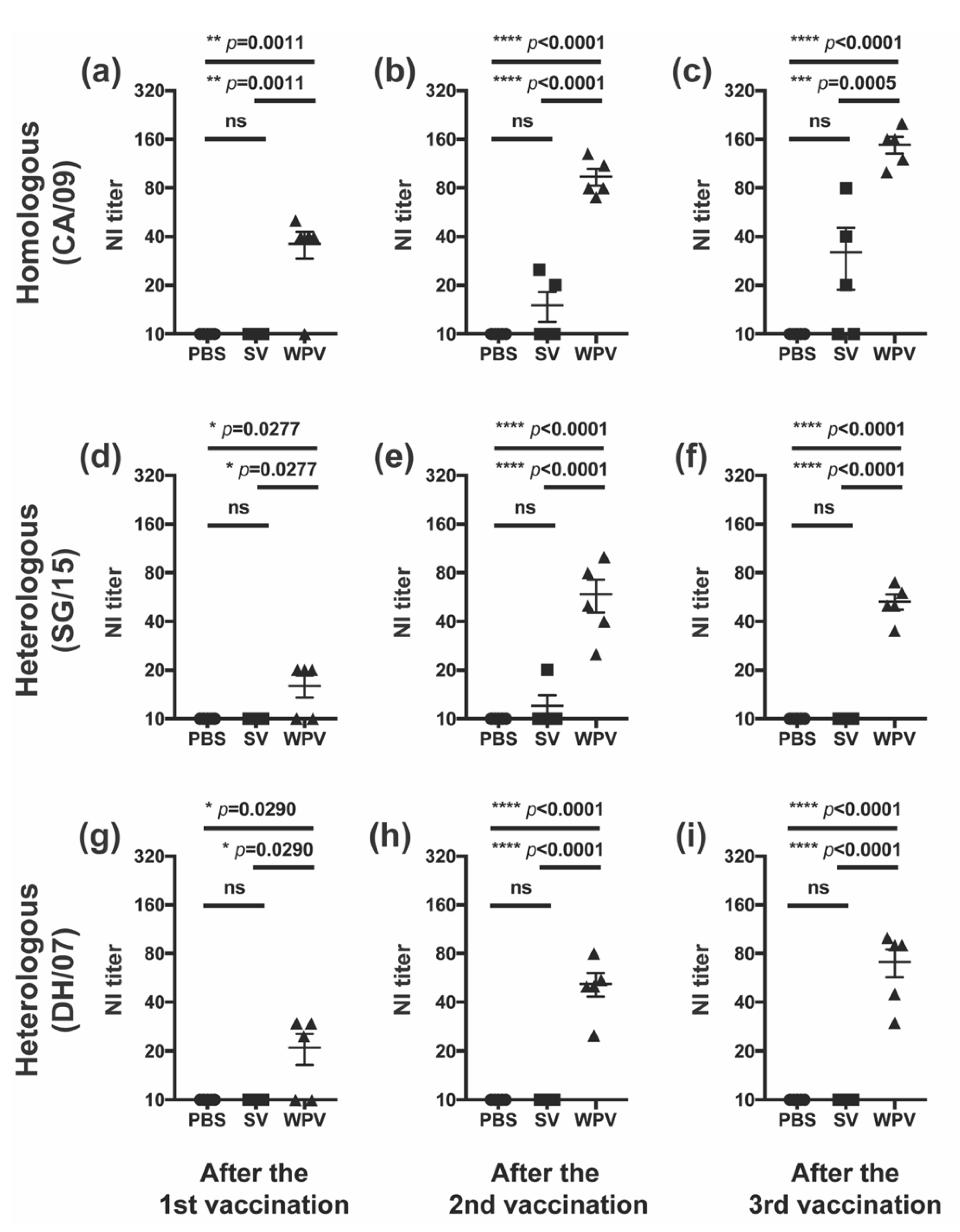
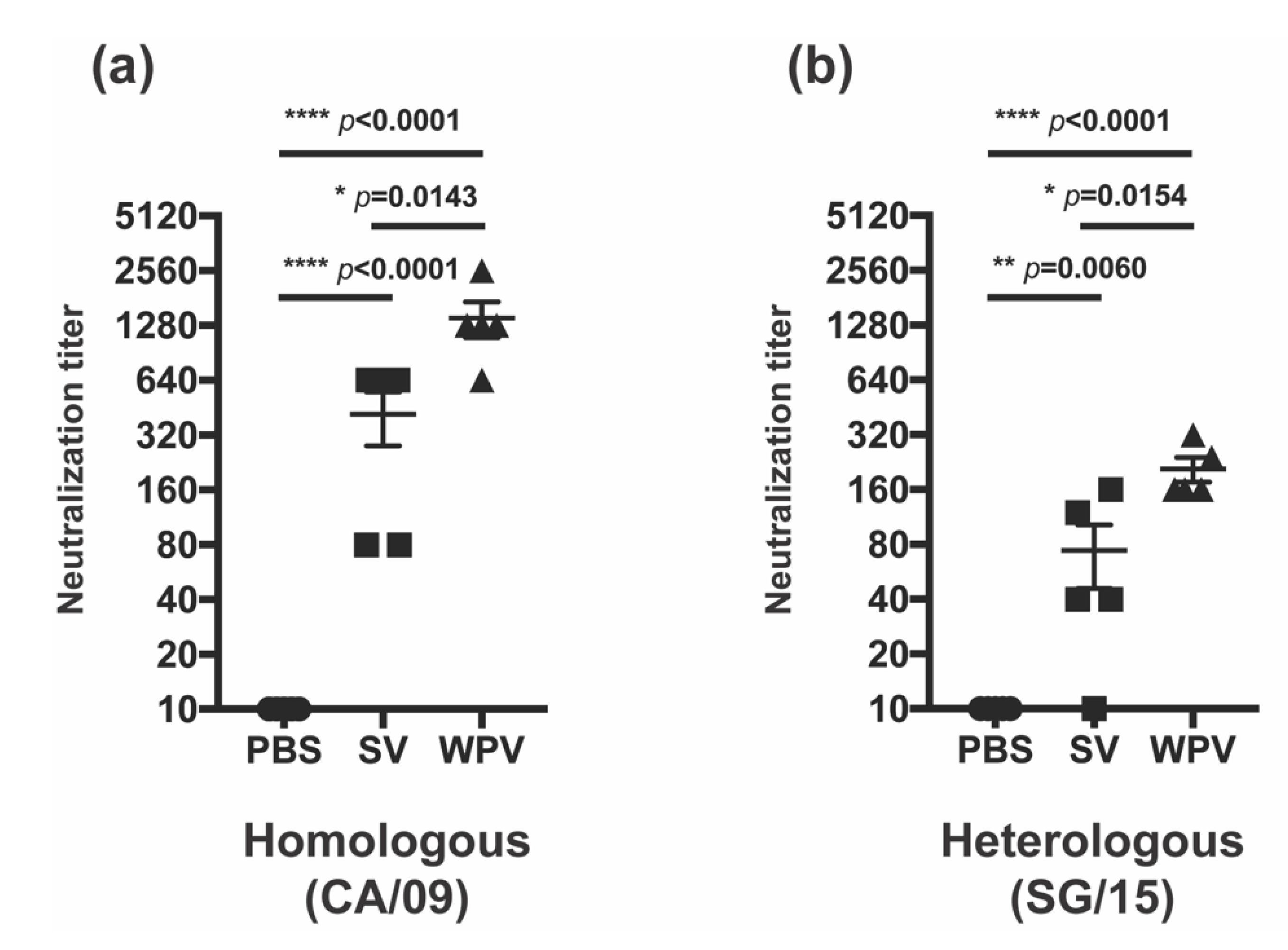
3.3. WPV Induced Better NI Antibodies than SV
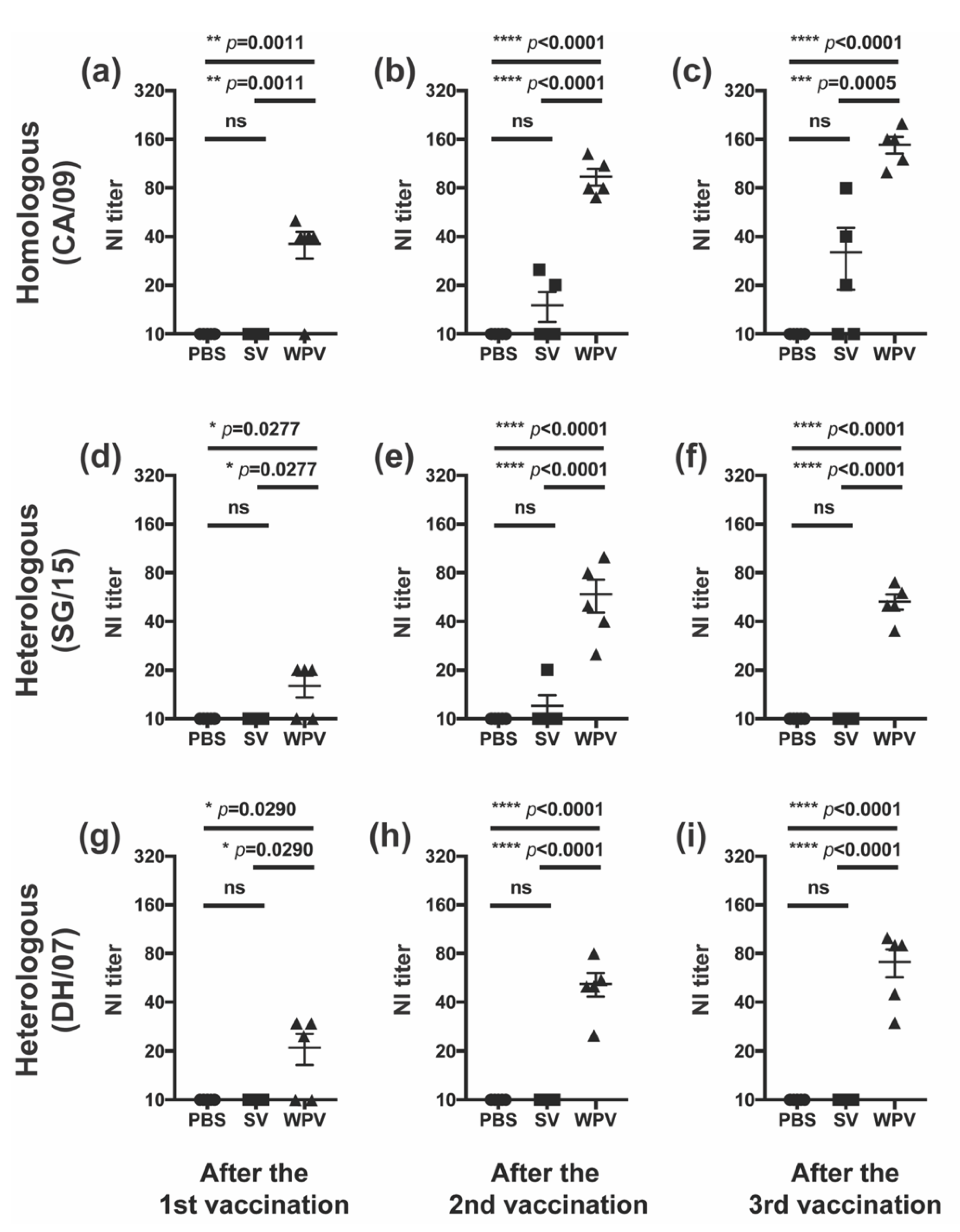
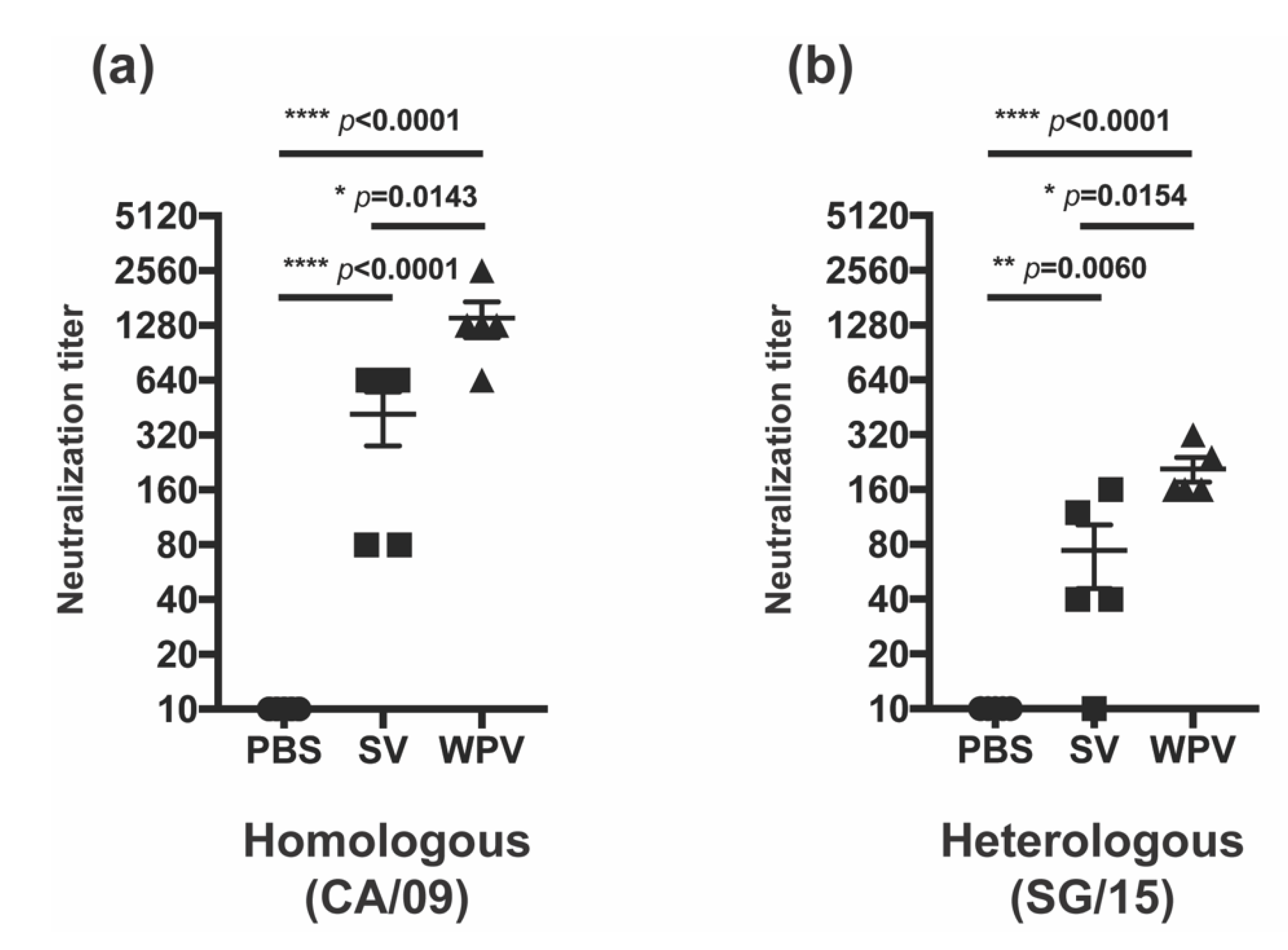
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stöhr, K. Influenza–WHO cares. Lancet Infect. Dis. 2002, 2, 517. [Google Scholar] [CrossRef]
- World Health Organization. Manual for the Laboratory Diagnosis and Virological Surveillance of Influenza; World Health Organization: Geneva, Switzerland, 2011; p. 153. [Google Scholar]
- Zambon, M.C. Epidemiology and pathogenesis of influenza. J. Antimicrob. Chemother. 1999, 44, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Petrova, V.N.; Russell, C.A. The evolution of seasonal influenza viruses. Nat. Rev. Microbiol. 2018, 16, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Grohskopf, L.A.; Alyanak, E.; Broder, K.R.; Blanton, L.H.; Fry, A.M.; Jernigan, D.B.; Atmar, R.L. Prevention and Control of Seasonal Influenza with Vaccines: Recommendations of the Advisory Committee on Immunization Practices—United States, 2020–2021 Influenza Season. MMWR Recomm. Rep. 2020, 69, 1–24. [Google Scholar] [CrossRef]
- Salomon, R.; Webster, R.G. The Influenza Virus Enigma. Cell 2009, 136, 402–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, J.M. Selection of influenza vaccine strains and developing pandemic vaccines. Vaccine 2002, 20, 40–44. [Google Scholar] [CrossRef]
- Coudeville, L.; Bailleux, F.; Riche, B.; Megas, F.; Andre, P.; Ecochard, R. Relationship between haemagglutination-inhibiting antibody titres and clinical protection against influenza: Development and application of a bayesian random-effects model. BMC Med. Res. Methodol. 2010, 10, 18. [Google Scholar] [CrossRef] [Green Version]
- Belongia, E.A.; Simpson, M.D.; King, J.P.; Sundaram, M.E.; Kelley, N.S.; Osterholm, M.T.; McLean, H.Q. Variable influenza vaccine effectiveness by subtype: A systematic review and meta-analysis of test-negative design studies. Lancet Infect. Dis. 2016, 16, 942–951. [Google Scholar] [CrossRef]
- Tenforde, M.W.; Kondor, R.J.G.; Chung, J.R.; Zimmerman, R.K.; Nowalk, M.P.; Jackson, M.L.; Jackson, L.A.; Monto, A.S.; Martin, E.T.; Belongia, E.A.; et al. Effect of Antigenic Drift on Influenza Vaccine Effectiveness in the United States—2019–2020. Clin. Infect. Dis. 2021, 73, e4244–e4250. [Google Scholar] [CrossRef]
- Sekiya, T.; Ohno, M.; Nomura, N.; Handabile, C.; Shingai, M.; Jackson, D.C.; Brown, L.E.; Kida, H. Selecting and Using the Appropriate Influenza Vaccine for Each Individual. Viruses 2021, 13, 971. [Google Scholar] [CrossRef]
- Sekiya, T.; Mifsud, E.J.; Ohno, M.; Nomura, N.; Sasada, M.; Fujikura, D.; Daito, T.; Shingai, M.; Ohara, Y.; Nishimura, T.; et al. Inactivated whole virus particle vaccine with potent immunogenicity and limited IL-6 induction is ideal for influenza. Vaccine 2019, 37, 2158–2166. [Google Scholar] [CrossRef]
- Shingai, M.; Nomura, N.; Sekiya, T.; Ohno, M.; Fujikura, D.; Handabile, C.; Omori, R.; Ohara, Y.; Nishimura, T.; Endo, M.; et al. Potent priming by inactivated whole influenza virus particle vaccines is linked to viral RNA uptake into antigen presenting cells. Vaccine 2021, 39, 3940–3951. [Google Scholar] [CrossRef] [PubMed]
- Hobson, D.; Curry, R.L.; Beare, A.S.; Ward-Gardner, A. The role of serum haemagglutination-inhibiting antibody in protection against challenge infection with influenza A2 and B viruses. J. Hyg. 1972, 70, 767–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohmit, S.E.; Petrie, J.G.; Cross, R.T.; Johnson, E.; Monto, A.S. Influenza hemagglutination-inhibition antibody titer as a correlate of vaccine-induced protection. J. Infect. Dis. 2011, 204, 1879–1885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunning, A.J.; DiazGranados, C.A.; Voloshen, T.; Hu, B.; Landolfi, V.A.; Talbot, H.K. Correlates of Protection against Influenza in the Elderly: Results from an Influenza Vaccine Efficacy Trial. Clin. Vaccine Immunol. 2016, 23, 228–235. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Fang, V.J.; Ohmit, S.E.; Monto, A.S.; Cook, A.R.; Cowling, B.J. Quantifying Protection Against Influenza Virus Infection Measured by Hemagglutination-inhibition Assays in Vaccine Trials. Epidemiology 2016, 27, 143–151. [Google Scholar] [CrossRef] [Green Version]
- Hay, A.J.; Gregory, V.; Douglas, A.R.; Lin, Y.P. The evolution of human influenza viruses. Philos. Trans. R. Soc. B Biol. Sci. 2001, 356, 1861–1870. [Google Scholar] [CrossRef] [Green Version]
- Itoh, Y.; Ozaki, H.; Tsuchiya, H.; Okamoto, K.; Torii, R.; Sakoda, Y.; Kawaoka, Y.; Ogasawara, K.; Kida, H. A vaccine prepared from a non-pathogenic H5N1 avian influenza virus strain confers protective immunity against highly pathogenic avian influenza virus infection in cynomolgus macaques. Vaccine 2008, 26, 562–572. [Google Scholar] [CrossRef]
- Takada, A.; Matsushita, S.; Ninomiya, A.; Kawaoka, Y.; Kida, H. Intranasal immunization with formalin-inactivated virus vaccine induces a broad spectrum of heterosubtypic immunity against influenza A virus infection in mice. Vaccine 2003, 21, 3212–3218. [Google Scholar] [CrossRef]
- Aymard-Henry, M.; Coleman, M.T.; Dowdle, W.R.; Laver, W.G.; Schild, G.C.; Webster, R.G. Influenza virus neuraminidase and neuraminidase inhibition test procedures. Bull. World Health Organ. 1973, 48, 199–202. [Google Scholar]
- Chen, Y.Q.; Wohlbold, T.J.; Zheng, N.Y.; Huang, M.; Huang, Y.; Neu, K.E.; Lee, J.; Wan, H.; Rojas, K.T.; Kirkpatrick, E.; et al. Influenza Infection in Humans Induces Broadly Cross-Reactive and Protective Neuraminidase-Reactive Antibodies. Cell 2018, 173, 417–429.e10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, S.; Nachbagauer, R.; Balmaseda, A.; Stadlbauer, D.; Ojeda, S.; Patel, M.; Rajabhathor, A.; Lopez, R.; Guglia, A.F.; Sanchez, N.; et al. Novel correlates of protection against pandemic H1N1 influenza A virus infection. Nat. Med. 2019, 25, 962–967. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Lee, Y.; Park, S.; Jung, Y.; Lee, Y.; Ko, E.; Kim, Y.; Li, X.; Kang, S. Neuraminidase expressing virus-like particle vaccine provides effective cross-protection against influenza virus. Virology 2019, 535, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Wohlbold, T.J.; Nachbagauer, R.; Xu, H.; Tan, G.S.; Hirsh, A.; Brokstad, K.A.; Cox, R.J.; Palese, P.; Krammer, F. Vaccination with adjuvanted recombinant neuraminidase induces broad heterologous, but not heterosubtypic, cross-protection against influenza virus infection in mice. MBio 2015, 6, e02556. [Google Scholar] [CrossRef] [Green Version]
- Rockman, S.; Brown, L.E.; Barr, I.G.; Gilbertson, B.; Lowther, S.; Kachurin, A.; Kachurina, O.; Klippel, J.; Bodle, J.; Pearse, M.; et al. Neuraminidase-Inhibiting Antibody Is a Correlate of Cross-Protection against Lethal H5N1 Influenza Virus in Ferrets Immunized with Seasonal Influenza Vaccine. J. Virol. 2013, 87, 3053–3061. [Google Scholar] [CrossRef] [Green Version]
- Gilchuk, I.M.; Bangaru, S.; Gilchuk, P.; Irving, R.P.; Kose, N.; Bombardi, R.G.; Thornburg, N.J.; Creech, C.B.; Edwards, K.M.; Li, S.; et al. Influenza H7N9 Virus Neuraminidase-Specific Human Monoclonal Antibodies Inhibit Viral Egress and Protect from Lethal Influenza Infection in Mice. Cell Host Microbe 2019, 26, 715–728.e8. [Google Scholar] [CrossRef]
- Sandbulte, M.R.; Jimenez, G.S.; Boon, A.C.M.; Smith, L.R.; Treanor, J.J.; Webby, R.J. Cross-reactive neuraminidase antibodies afford partial protection against H5N1 in mice and are present in unexposed humans. PLoS Med. 2007, 4, 0265–0272. [Google Scholar] [CrossRef] [Green Version]
- Easterbrook, J.D.; Schwartzman, L.M.; Gao, J.; Kash, J.C.; Morens, D.M.; Couzens, L.; Wan, H.; Eichelberger, M.C.; Taubenberger, J.K. Protection against a lethal H5N1 influenza challenge by intranasal immunization with virus-like particles containing 2009 pandemic H1N1 neuraminidase in mice. Virology 2012, 432, 39–44. [Google Scholar] [CrossRef]
- Schulman, J.L.; Khakpour, M.; Kilbourne, E.D. Protective Effects of Specific Immunity to Viral Neuraminidase on Influenza Virus Infection of Mice. J. Virol. 1968, 2, 778–786. [Google Scholar] [CrossRef] [Green Version]
- Barry, D.W.; Mayner, R.E.; Hochstein, H.D.; Dunlap, R.C.; Rastogi, S.C.; Hannah, J.E.; Blackburn, R.J.; Sullivan, J.L.; Gerety, R.J. Comparative Trial of Influenza Vaccines. Am. J. Epidemiol. 1976, 104, 47–59. [Google Scholar] [CrossRef]
- Barry, D.W.; Staton, E.; Mayner, R.E. Inactivated influenza vaccine efficacy: Diminished antigenicity of split product vaccines in mice. Infect. Immun. 1974, 10, 1329–1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcelin, G.; DuBois, R.; Rubrum, A.; Russell, C.J.; McElhaney, J.E.; Webby, R.J. A contributing role for anti-neuraminidase antibodies on immunity to pandemic H1N1 2009 influenza a virus. PLoS ONE 2011, 6, e26335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monto, A.S.; Kendal, A.P. Effect of Neuraminidase Antibody on Hong Kong Influenza. Lancet 1973, 301, 623–625. [Google Scholar] [CrossRef] [Green Version]
- Schulman, J.L. The role of antineuraminidase antibody in immunity to influenza virus infection. Bull. World Health Organ. 1969, 41, 647–650. [Google Scholar] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Handabile, C.; Sekiya, T.; Nomura, N.; Ohno, M.; Kawakita, T.; Shingai, M.; Kida, H. Inactivated Whole Virus Particle Influenza Vaccine Induces Anti-Neuraminidase Antibodies That May Contribute to Cross-Protection against Heterologous Virus Infection. Vaccines 2022, 10, 804. https://doi.org/10.3390/vaccines10050804
Handabile C, Sekiya T, Nomura N, Ohno M, Kawakita T, Shingai M, Kida H. Inactivated Whole Virus Particle Influenza Vaccine Induces Anti-Neuraminidase Antibodies That May Contribute to Cross-Protection against Heterologous Virus Infection. Vaccines. 2022; 10(5):804. https://doi.org/10.3390/vaccines10050804
Chicago/Turabian StyleHandabile, Chimuka, Toshiki Sekiya, Naoki Nomura, Marumi Ohno, Tomomi Kawakita, Masashi Shingai, and Hiroshi Kida. 2022. "Inactivated Whole Virus Particle Influenza Vaccine Induces Anti-Neuraminidase Antibodies That May Contribute to Cross-Protection against Heterologous Virus Infection" Vaccines 10, no. 5: 804. https://doi.org/10.3390/vaccines10050804
APA StyleHandabile, C., Sekiya, T., Nomura, N., Ohno, M., Kawakita, T., Shingai, M., & Kida, H. (2022). Inactivated Whole Virus Particle Influenza Vaccine Induces Anti-Neuraminidase Antibodies That May Contribute to Cross-Protection against Heterologous Virus Infection. Vaccines, 10(5), 804. https://doi.org/10.3390/vaccines10050804