
Roles of the Tol/Pal System in Bacterial Pathogenesis and Its Application to Antibacterial Therapy



Abstract
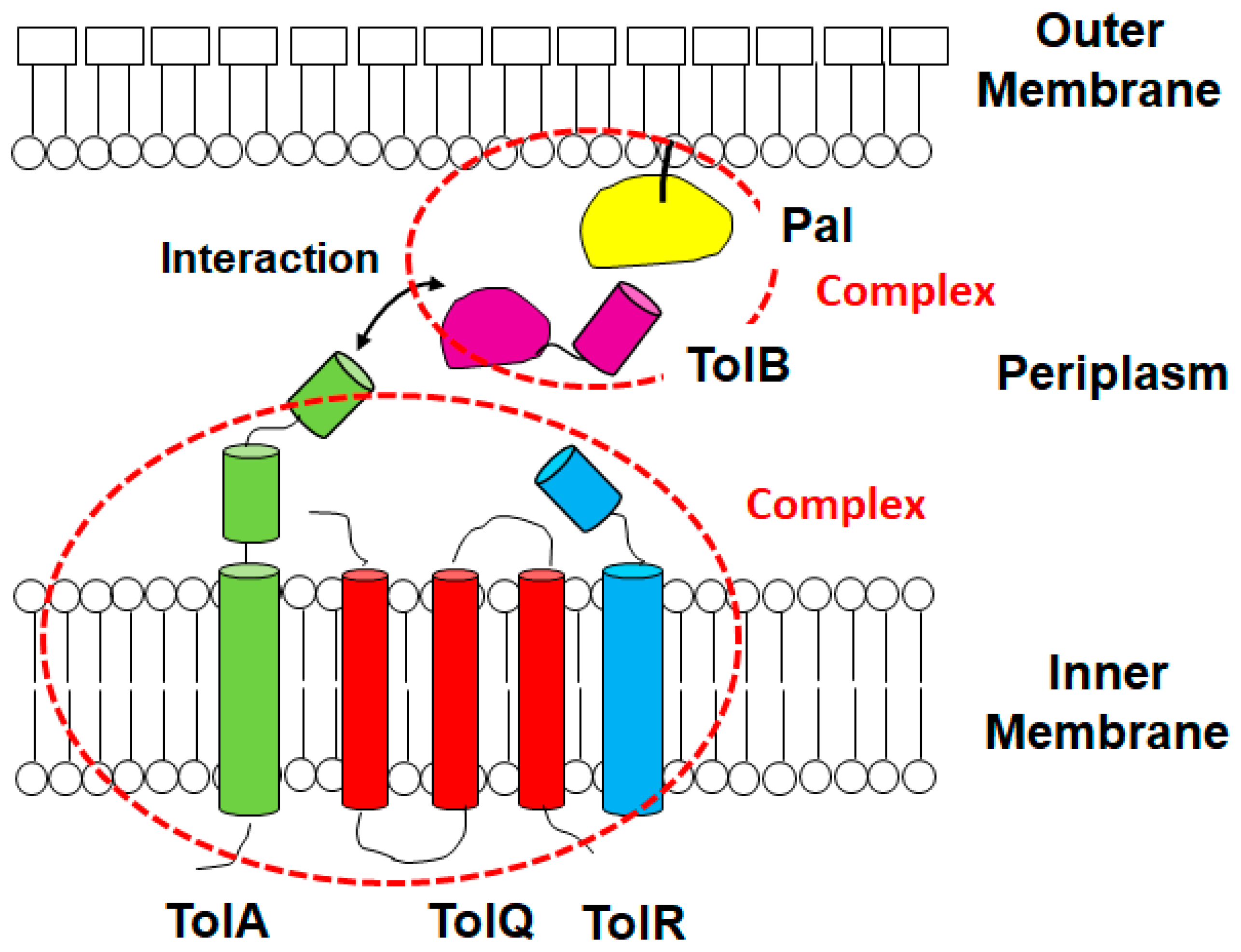
1. Introduction
2. Roles of the Tol/Pal System in Pathogenesis

{kind=link}
| Bacterial Species | Pathogenesis | References |
|---|---|---|
| Enterohemorrhagic Escherichia coli (EHEC) | Secretion of the T3SS effector proteins, A/E lesion formation, Flagellar synthesis | [34] |
| Citrobacter rodentium | Lethality and Enteritis in mouse | [34] |
| Uropathogenic Escherichia coli (UPEC) | Flagellar synthesis, Bacterial colonization within bladder epithelial cells and in the urinary tract of mice | [35] |
| Escherichia coli | Development of sepsis | [36] |
| Salmonella enterica (Typhimurium) | Bacterial survival in macrophage and mouse, Innate tolerance to bile acid and serum | [37,38,39,40] |
| Salmonella enterica (Choleraesuis) | Lethality in mice, innate tolerance to deoxycholate and vancomycin | [41] |
| Shigella flexneri | Bacterial invasion and growth in epithelial cells, Innate tolerance to antibiotics, bile acid and human complement | [42] |
| Pseudomonas aeruginosa | Bacterial growth | [43,44,45] |
| Burkholderia cenocepacia | Bacterial adhesion to lung cells, Lethality in G. mellonella | [46] |
| Klebsiella pneumoniae | Innate tolerance to serum and phagocytosis, Lethality in mice | [47,48] |
| Haemophilus ducreyi | Papules formation, Bacterial survival | [49,50] |
| Vibrio cholerae | Uptake of the CTXphi phase, Bacterial growth at high temperature | [33] |
| Edwardsiella ictaluri | Mortality in catfish | [51] |
| Dickeya dadantii (Erwinia chrysanthemi) | Activity of pectinolytic enzyme, Motility, Tissue injury on plant leaf | [52] |
2.1. Escherichia coli Pathogenic Subgroups
2.2. Salmonella enterica
2.3. Shigella flexneri
2.4. Pseudomonas aeruginosa
2.5. Burkholderia cenocepacia
2.6. Klebsiella pneumoniae
2.7. Haemophilus ducreyi
2.8. Vibrio cholerae
2.9. Edwardsiella ictaluri
2.10. Dickeya dadantii (Formerly Named Erwinia chrysanthemi)
3. Immunogenicity of the tol/pal Mutants and Outer Membrane Vesicles (OMVs)
4. Pal Proteins for Vaccine Strategies
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zgurskaya, H.I.; Lopez, C.A.; Gnanakaran, S. Permeability Barrier of Gram-Negative Cell Envelopes and Approaches to Bypass It. ACS Infect. Dis. 2015, 1, 512–522. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Z.; Plesiat, P.; Nikaido, H. The challenge of efflux-mediated antibiotic resistance in Gram-negative bacteria. Clin. Microbiol. Rev. 2015, 28, 337–418. [Google Scholar] [CrossRef] [PubMed]
- Delcour, A.H. Outer membrane permeability and antibiotic resistance. Biochim. Biophys. Acta 2009, 1794, 808–816. [Google Scholar] [CrossRef] [PubMed]
- Lazzaroni, J.C.; Germon, P.; Ray, M.C.; Vianney, A. The Tol proteins of Escherichia coli and their involvement in the uptake of biomolecules and outer membrane stability. FEMS Microbiol. Lett. 1999, 177, 191–197. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Krachler, A.M.; Sharma, A.; Cauldwell, A.; Papadakos, G.; Kleanthous, C. TolA modulates the oligomeric status of YbgF in the bacterial periplasm. J. Mol. Biol. 2010, 403, 270–285. [Google Scholar] [CrossRef]
- Muller, M.M.; Webster, R.E. Characterization of the tol-pal and cyd region of Escherichia coli K-12: Transcript analysis and identification of two new proteins encoded by the cyd operon. J. Bacteriol. 1997, 179, 2077–2080. [Google Scholar] [CrossRef][Green Version]
- Vianney, A.; Muller, M.M.; Clavel, T.; Lazzaroni, J.C.; Portalier, R.; Webster, R.E. Characterization of the tol-pal region of Escherichia coli K-12: Translational control of tolR expression by TolQ and identification of a new open reading frame downstream of pal encoding a periplasmic protein. J. Bacteriol. 1996, 178, 4031–4038. [Google Scholar] [CrossRef][Green Version]
- Clavel, T.; Lazzaroni, J.C.; Vianney, A.; Portalier, R. Expression of the tolQRA genes of Escherichia coli K-12 is controlled by the RcsC sensor protein involved in capsule synthesis. Mol. Microbiol. 1996, 19, 19–25. [Google Scholar] [CrossRef]
- Duan, K.; Lafontaine, E.R.; Majumdar, S.; Sokol, P.A. RegA, iron, and growth phase regulate expression of the Pseudomonas aeruginosa tol-oprL gene cluster. J. Bacteriol. 2000, 182, 2077–2087. [Google Scholar] [CrossRef]
- Liu, L.; Li, F.; Xu, L.; Wang, J.; Li, M.; Yuan, J.; Wang, H.; Yang, R.; Li, B. Cyclic AMP-CRP Modulates the Cell Morphology of Klebsiella pneumoniae in High-Glucose Environment. Front. Microbiol. 2019, 10, 2984. [Google Scholar] [CrossRef]
- Bouveret, E.; Derouiche, R.; Rigal, A.; Lloubes, R.; Lazdunski, C.; Benedetti, H. Peptidoglycan-associated lipoprotein-TolB interaction. A possible key to explaining the formation of contact sites between the inner and outer membranes of Escherichia coli. J. Biol. Chem. 1995, 270, 11071–11077. [Google Scholar] [CrossRef] [PubMed]
- Derouiche, R.; Benedetti, H.; Lazzaroni, J.C.; Lazdunski, C.; Lloubes, R. Protein complex within Escherichia coli inner membrane. TolA N-terminal domain interacts with TolQ and TolR proteins. J. Biol. Chem. 1995, 270, 11078–11084. [Google Scholar] [CrossRef] [PubMed]
- Eick-Helmerich, K.; Braun, V. Import of biopolymers into Escherichia coli: Nucleotide sequences of the exbB and exbD genes are homologous to those of the tolQ and tolR genes, respectively. J. Bacteriol. 1989, 171, 5117–5126. [Google Scholar] [CrossRef] [PubMed]
- Cascales, E.; Lloubes, R.; Sturgis, J.N. The TolQ-TolR proteins energize TolA and share homologies with the flagellar motor proteins MotA-MotB. Mol. Microbiol. 2001, 42, 795–807. [Google Scholar] [CrossRef] [PubMed]
- Walburger, A.; Lazdunski, C.; Corda, Y. The Tol/Pal system function requires an interaction between the C-terminal domain of TolA and the N-terminal domain of TolB. Mol. Microbiol. 2002, 44, 695–708. [Google Scholar] [CrossRef] [PubMed]
- Bonsor, D.A.; Hecht, O.; Vankemmelbeke, M.; Sharma, A.; Krachler, A.M.; Housden, N.G.; Lilly, K.J.; James, R.; Moore, G.R.; Kleanthous, C. Allosteric beta-propeller signalling in TolB and its manipulation by translocating colicins. EMBO J. 2009, 28, 2846–2857. [Google Scholar] [CrossRef]
- Szczepaniak, J.; Holmes, P.; Rajasekar, K.; Kaminska, R.; Samsudin, F.; Inns, P.G.; Rassam, P.; Khalid, S.; Murray, S.M.; Redfield, C.; et al. The lipoprotein Pal stabilises the bacterial outer membrane during constriction by a mobilisation-and-capture mechanism. Nat. Commun. 2020, 11, 1305. [Google Scholar] [CrossRef]
- Clavel, T.; Germon, P.; Vianney, A.; Portalier, R.; Lazzaroni, J.C. TolB protein of Escherichia coli K-12 interacts with the outer membrane peptidoglycan-associated proteins Pal, Lpp and OmpA. Mol. Microbiol. 1998, 29, 359–367. [Google Scholar] [CrossRef]
- Zhuang, Z.; Song, F.; Martin, B.M.; Dunaway-Mariano, D. The YbgC protein encoded by the ybgC gene of the tol-pal gene cluster of Haemophilus influenzae catalyzes acyl-coenzyme A thioester hydrolysis. FEBS Lett. 2002, 516, 161–163. [Google Scholar] [CrossRef]
- Gray, A.N.; Egan, A.J.; Van’t Veer, I.L.; Verheul, J.; Colavin, A.; Koumoutsi, A.; Biboy, J.; Altelaar, A.F.; Damen, M.J.; Huang, K.C.; et al. Coordination of peptidoglycan synthesis and outer membrane constriction during Escherichia coli cell division. eLife 2015, 4, e07118. [Google Scholar] [CrossRef]
- Szczepaniak, J.; Press, C.; Kleanthous, C. The multifarious roles of Tol-Pal in Gram-negative bacteria. FEMS Microbiol. Rev. 2020, 44, 490–506. [Google Scholar] [CrossRef] [PubMed]
- Nagel de Zwaig, R.; Luria, S.E. Genetics and physiology of colicin-tolerant mutants of Escherichia coli. J. Bacteriol. 1967, 94, 1112–1123. [Google Scholar] [CrossRef] [PubMed]
- Cascales, E.; Bernadac, A.; Gavioli, M.; Lazzaroni, J.C.; Lloubes, R. Pal lipoprotein of Escherichia coli plays a major role in outer membrane integrity. J. Bacteriol. 2002, 184, 754–759. [Google Scholar] [CrossRef] [PubMed]
- Riechmann, L.; Holliger, P. The C-terminal domain of TolA is the coreceptor for filamentous phage infection of E. coli. Cell 1997, 90, 351–360. [Google Scholar] [CrossRef]
- Mouslim, C.; Latifi, T.; Groisman, E.A. Signal-dependent requirement for the co-activator protein RcsA in transcription of the RcsB-regulated ugd gene. J. Biol. Chem. 2003, 278, 50588–50595. [Google Scholar] [CrossRef]
- Click, E.M.; Webster, R.E. The TolQRA proteins are required for membrane insertion of the major capsid protein of the filamentous phage f1 during infection. J. Bacteriol. 1998, 180, 1723–1728. [Google Scholar] [CrossRef]
- Lazdunski, C.; Shapiro, B.M. Isolation and some properties of cell envelope altered mutants of Escherichia coli. J. Bacteriol. 1972, 111, 495–498. [Google Scholar] [CrossRef]
- Davies, J.K.; Reeves, P. Genetics of resistance to colicins in Escherichia coli K-12: Cross-resistance among colicins of group A. J. Bacteriol. 1975, 123, 102–117. [Google Scholar] [CrossRef]
- Onodera, K.; Rolfe, B.; Bernstein, A. Demonstration of missing membrane proteins in deletion mutants of E. coli K12. Biochem. Biophys. Res. Commun. 1970, 39, 969–975. [Google Scholar] [CrossRef]
- Foulds, J.; Barrett, C. Characterization of Escherichia coli mutants tolerant to bacteriocin JF246: Two new classes of tolerant mutants. J. Bacteriol. 1973, 116, 885–892. [Google Scholar] [CrossRef]
- Meury, J.; Devilliers, G. Impairment of cell division in tolA mutants of Escherichia coli at low and high medium osmolarities. Biol. Cell 1999, 91, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Llamas, M.A.; Ramos, J.L.; Rodriguez-Herva, J.J. Mutations in each of the tol genes of Pseudomonas putida reveal that they are critical for maintenance of outer membrane stability. J. Bacteriol. 2000, 182, 4764–4772. [Google Scholar] [CrossRef] [PubMed]
- Heilpern, A.J.; Waldor, M.K. CTXphi infection of Vibrio cholerae requires the tolQRA gene products. J. Bacteriol. 2000, 182, 1739–1747. [Google Scholar] [CrossRef] [PubMed]
- Hueck, C.J. Type III protein secretion systems in bacterial pathogens of animals and plants. Microbiol. Mol. Biol. Rev. 1998, 62, 379–433. [Google Scholar] [CrossRef]
- Galan, J.E.; Wolf-Watz, H. Protein delivery into eukaryotic cells by type III secretion machines. Nature 2006, 444, 567–573. [Google Scholar] [CrossRef]
- Kenny, B.; DeVinney, R.; Stein, M.; Reinscheid, D.J.; Frey, E.A.; Finlay, B.B. Enteropathogenic E. coli (EPEC) transfers its receptor for intimate adherence into mammalian cells. Cell 1997, 91, 511–520. [Google Scholar] [CrossRef]
- Hirakawa, H.; Suzue, K.; Takita, A.; Awazu, C.; Kurushima, J.; Tomita, H. Roles of the Tol-Pal system in the Type III secretion system and flagella-mediated virulence in enterohemorrhagic Escherichia coli. Sci. Rep. 2020, 10, 15173. [Google Scholar] [CrossRef]
- Flores-Mireles, A.L.; Walker, J.N.; Caparon, M.; Hultgren, S.J. Urinary tract infections: Epidemiology, mechanisms of infection and treatment options. Nat. Rev. Microbiol. 2015, 13, 269–284. [Google Scholar] [CrossRef]
- Anderson, G.G.; Palermo, J.J.; Schilling, J.D.; Roth, R.; Heuser, J.; Hultgren, S.J. Intracellular bacterial biofilm-like pods in urinary tract infections. Science 2003, 301, 105–107. [Google Scholar] [CrossRef]
- Wright, K.J.; Seed, P.C.; Hultgren, S.J. Uropathogenic Escherichia coli flagella aid in efficient urinary tract colonization. Infect. Immun. 2005, 73, 7657–7668. [Google Scholar] [CrossRef]
- Lane, M.C.; Lockatell, V.; Monterosso, G.; Lamphier, D.; Weinert, J.; Hebel, J.R.; Johnson, D.E.; Mobley, H.L. Role of motility in the colonization of uropathogenic Escherichia coli in the urinary tract. Infect. Immun. 2005, 73, 7644–7656. [Google Scholar] [CrossRef] [PubMed]
- Hirakawa, H.; Suzue, K.; Kurabayashi, K.; Tomita, H. The Tol-Pal System of Uropathogenic Escherichia coli Is Responsible for Optimal Internalization into and Aggregation Within Bladder Epithelial Cells, Colonization of the Urinary Tract of Mice, and Bacterial Motility. Front. Microbiol. 2019, 10, 1827. [Google Scholar] [CrossRef] [PubMed]
- Pichon, C.; Hechard, C.; du Merle, L.; Chaudray, C.; Bonne, I.; Guadagnini, S.; Vandewalle, A.; Le Bouguenec, C. Uropathogenic Escherichia coli AL511 requires flagellum to enter renal collecting duct cells. Cell Microbiol. 2009, 11, 616–628. [Google Scholar] [CrossRef] [PubMed]
- Hirakawa, H.; Suzue, K.; Takita, A.; Kamitani, W.; Tomita, H. Roles of OmpX, an Outer Membrane Protein, on Virulence and Flagellar Expression in Uropathogenic Escherichia coli. Infect. Immun. 2021, 89, e00721-20. [Google Scholar] [CrossRef] [PubMed]
- Hellman, J.; Roberts, J.D., Jr.; Tehan, M.M.; Allaire, J.E.; Warren, H.S. Bacterial peptidoglycan-associated lipoprotein is released into the bloodstream in gram-negative sepsis and causes inflammation and death in mice. J. Biol. Chem. 2002, 277, 14274–14280. [Google Scholar] [CrossRef]
- Sabbagh, S.C.; Forest, C.G.; Lepage, C.; Leclerc, J.M.; Daigle, F. So similar, yet so different: Uncovering distinctive features in the genomes of Salmonella enterica serovars Typhimurium and Typhi. FEMS Microbiol. Lett. 2010, 305, 1–13. [Google Scholar] [CrossRef]
- Palmer, A.D.; Slauch, J.M. Mechanisms of Salmonella pathogenesis in animal models. Hum. Ecol. Risk Assess. 2017, 23, 1877–1892. [Google Scholar] [CrossRef]
- Bowe, F.; Lipps, C.J.; Tsolis, R.M.; Groisman, E.; Heffron, F.; Kusters, J.G. At least four percent of the Salmonella typhimurium genome is required for fatal infection of mice. Infect. Immun. 1998, 66, 3372–3377. [Google Scholar] [CrossRef]
- Masilamani, R.; Cian, M.B.; Dalebroux, Z.D. Salmonella Tol-Pal Reduces Outer Membrane Glycerophospholipid Levels for Envelope Homeostasis and Survival during Bacteremia. Infect. Immun. 2018, 86, e00173-18. [Google Scholar] [CrossRef]
- Prouty, A.M.; Van Velkinburgh, J.C.; Gunn, J.S. Salmonella enterica serovar typhimurium resistance to bile: Identification and characterization of the tolQRA cluster. J. Bacteriol. 2002, 184, 1270–1276. [Google Scholar] [CrossRef]
- Mouslim, C.; Groisman, E.A. Control of the Salmonella ugd gene by three two-component regulatory systems. Mol. Microbiol. 2003, 47, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Li, Z.; Fei, X.; Tian, Y.; Zhou, G.; Hu, Y.; Wang, S.; Shi, H. The role of TolA, TolB, and TolR in cell morphology, OMVs production, and virulence of Salmonella choleraesuis. AMB Express 2022, 12, 5. [Google Scholar] [CrossRef] [PubMed]
- Jennison, A.V.; Verma, N.K. Shigella flexneri infection: Pathogenesis and vaccine development. FEMS Microbiol. Rev. 2004, 28, 43–58. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Sasakawa, C. Molecular basis of the intracellular spreading of Shigella. Infect. Immun. 2001, 69, 5959–5966. [Google Scholar] [CrossRef] [PubMed]
- Pastor, Y.; Camacho, A.I.; Zuniga-Ripa, A.; Merchan, A.; Rosas, P.; Irache, J.M.; Gamazo, C. Towards a subunit vaccine from a Shigella flexneri DeltatolR mutant. Vaccine 2018, 36, 7509–7519. [Google Scholar] [CrossRef]
- Poole, K. Pseudomonas aeruginosa: Resistance to the max. Front. Microbiol. 2011, 2, 65. [Google Scholar] [CrossRef]
- Harmsen, M.; Yang, L.; Pamp, S.J.; Tolker-Nielsen, T. An update on Pseudomonas aeruginosa biofilm formation, tolerance, and dispersal. FEMS Immunol. Med. Microbiol. 2010, 59, 253–268. [Google Scholar] [CrossRef]
- Hoiby, N.; Ciofu, O.; Bjarnsholt, T. Pseudomonas aeruginosa biofilms in cystic fibrosis. Future Microbiol. 2010, 5, 1663–1674. [Google Scholar] [CrossRef]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial biofilms: A common cause of persistent infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef]
- Ciofu, O.; Tolker-Nielsen, T. Tolerance and Resistance of Pseudomonas aeruginosa Biofilms to Antimicrobial Agents-How P. aeruginosa Can Escape Antibiotics. Front. Microbiol. 2019, 10, 913. [Google Scholar] [CrossRef]
- Imperi, F.; Ciccosanti, F.; Perdomo, A.B.; Tiburzi, F.; Mancone, C.; Alonzi, T.; Ascenzi, P.; Piacentini, M.; Visca, P.; Fimia, G.M. Analysis of the periplasmic proteome of Pseudomonas aeruginosa, a metabolically versatile opportunistic pathogen. Proteomics 2009, 9, 1901–1915. [Google Scholar] [CrossRef] [PubMed]
- Whiteley, M.; Bangera, M.G.; Bumgarner, R.E.; Parsek, M.R.; Teitzel, G.M.; Lory, S.; Greenberg, E.P. Gene expression in Pseudomonas aeruginosa biofilms. Nature 2001, 413, 860–864. [Google Scholar] [CrossRef] [PubMed]
- Dennis, J.J.; Lafontaine, E.R.; Sokol, P.A. Identification and characterization of the tolQRA genes of Pseudomonas aeruginosa. J. Bacteriol. 1996, 178, 7059–7068. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Jacobs, M.A.; Alwood, A.; Thaipisuttikul, I.; Spencer, D.; Haugen, E.; Ernst, S.; Will, O.; Kaul, R.; Raymond, C.; Levy, R.; et al. Comprehensive transposon mutant library of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2003, 100, 14339–14344. [Google Scholar] [CrossRef]
- Liberati, N.T.; Urbach, J.M.; Miyata, S.; Lee, D.G.; Drenkard, E.; Wu, G.; Villanueva, J.; Wei, T.; Ausubel, F.M. An ordered, nonredundant library of Pseudomonas aeruginosa strain PA14 transposon insertion mutants. Proc. Natl. Acad. Sci. USA 2006, 103, 2833–2838. [Google Scholar] [CrossRef]
- Skurnik, D.; Roux, D.; Aschard, H.; Cattoir, V.; Yoder-Himes, D.; Lory, S.; Pier, G.B. A comprehensive analysis of in vitro and in vivo genetic fitness of Pseudomonas aeruginosa using high-throughput sequencing of transposon libraries. PLoS Pathog. 2013, 9, e1003582. [Google Scholar] [CrossRef]
- Lo Sciuto, A.; Fernandez-Pinar, R.; Bertuccini, L.; Iosi, F.; Superti, F.; Imperi, F. The periplasmic protein TolB as a potential drug target in Pseudomonas aeruginosa. PLoS ONE 2014, 9, e103784. [Google Scholar] [CrossRef]
- Drevinek, P.; Mahenthiralingam, E. Burkholderia cenocepacia in cystic fibrosis: Epidemiology and molecular mechanisms of virulence. Clin. Microbiol. Infect. 2010, 16, 821–830. [Google Scholar] [CrossRef]
- Dennehy, R.; Romano, M.; Ruggiero, A.; Mohamed, Y.F.; Dignam, S.L.; Mujica Troncoso, C.; Callaghan, M.; Valvano, M.A.; Berisio, R.; McClean, S. The Burkholderia cenocepacia peptidoglycan-associated lipoprotein is involved in epithelial cell attachment and elicitation of inflammation. Cell Microbiol. 2017, 19, e12691. [Google Scholar] [CrossRef]
- Podschun, R.; Ullmann, U. Klebsiella spp. as nosocomial pathogens: Epidemiology, taxonomy, typing methods, and pathogenicity factors. Clin. Microbiol. Rev. 1998, 11, 589–603. [Google Scholar] [CrossRef]
- Hsieh, P.F.; Liu, J.Y.; Pan, Y.J.; Wu, M.C.; Lin, T.L.; Huang, Y.T.; Wang, J.T. Klebsiella pneumoniae peptidoglycan-associated lipoprotein and murein lipoprotein contribute to serum resistance, antiphagocytosis, and proinflammatory cytokine stimulation. J. Infect. Dis. 2013, 208, 1580–1589. [Google Scholar] [CrossRef] [PubMed]
- Hellman, J.; Warren, H.S. Outer membrane protein A (OmpA), peptidoglycan-associated lipoprotein (PAL), and murein lipoprotein (MLP) are released in experimental Gram-negative sepsis. J. Endotoxin Res. 2001, 7, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Bauer, M.E.; Spinola, S.M. Localization of Haemophilus ducreyi at the pustular stage of disease in the human model of infection. Infect. Immun. 2000, 68, 2309–2314. [Google Scholar] [CrossRef]
- Spinola, S.M.; Hiltke, T.J.; Fortney, K.; Shanks, K.L. The conserved 18,000-molecular-weight outer membrane protein of Haemophilus ducreyi has homology to PAL. Infect. Immun. 1996, 64, 1950–1955. [Google Scholar] [CrossRef] [PubMed]
- Fortney, K.R.; Young, R.S.; Bauer, M.E.; Katz, B.P.; Hood, A.F.; Munson, R.S., Jr.; Spinola, S.M. Expression of peptidoglycan-associated lipoprotein is required for virulence in the human model of Haemophilus ducreyi infection. Infect. Immun. 2000, 68, 6441–6448. [Google Scholar] [CrossRef] [PubMed]
- Hiltke, T.J.; Bauer, M.E.; Klesney-Tait, J.; Hansen, E.J.; Munson, R.S., Jr.; Spinola, S.M. Effect of normal and immune sera on Haemophilus ducreyi 35000HP and its isogenic MOMP and LOS mutants. Microb. Pathog. 1999, 26, 93–102. [Google Scholar] [CrossRef]
- Deen, J.; Mengel, M.A.; Clemens, J.D. Epidemiology of cholera. Vaccine 2020, 38 (Suppl. S1), A31–A40. [Google Scholar] [CrossRef]
- Harris, J.B.; LaRocque, R.C.; Qadri, F.; Ryan, E.T.; Calderwood, S.B. Cholera. Lancet 2012, 379, 2466–2476. [Google Scholar] [CrossRef]
- Waldor, M.K.; Mekalanos, J.J. Lysogenic conversion by a filamentous phage encoding cholera toxin. Science 1996, 272, 1910–1914. [Google Scholar] [CrossRef]
- Klesius, P.H.; Shoemaker, C.A. Development and use of modified live Edwardsiella ictaluri vaccine against enteric septicemia of catfish. Adv. Vet. Med. 1999, 41, 523–537. [Google Scholar] [CrossRef]
- Abdelhamed, H.; Lu, J.; Lawrence, M.L.; Karsi, A. Involvement of tolQ and tolR genes in Edwardsiella ictaluri virulence. Microb. Pathog. 2016, 100, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Hugouvieux-Cotte-Pattat, N.; Condemine, G.; Nasser, W.; Reverchon, S. Regulation of pectinolysis in Erwinia chrysanthemi. Annu. Rev. Microbiol. 1996, 50, 213–257. [Google Scholar] [CrossRef] [PubMed]
- Hugouvieux-Cotte-Pattat, N.; Blot, N.; Reverchon, S. Identification of TogMNAB, an ABC transporter which mediates the uptake of pectic oligomers in Erwinia chrysanthemi 3937. Mol. Microbiol. 2001, 41, 1113–1123. [Google Scholar] [CrossRef]
- Dubuisson, J.F.; Vianney, A.; Hugouvieux-Cotte-Pattat, N.; Lazzaroni, J.C. Tol-Pal proteins are critical cell envelope components of Erwinia chrysanthemi affecting cell morphology and virulence. Microbiology 2005, 151, 3337–3347. [Google Scholar] [CrossRef] [PubMed]
- Grenier, A.M.; Duport, G.; Pages, S.; Condemine, G.; Rahbe, Y. The phytopathogen Dickeya dadantii (Erwinia chrysanthemi 3937) is a pathogen of the pea aphid. Appl. Environ. Microbiol. 2006, 72, 1956–1965. [Google Scholar] [CrossRef] [PubMed]
- Paterson, G.K.; Northen, H.; Cone, D.B.; Willers, C.; Peters, S.E.; Maskell, D.J. Deletion of tolA in Salmonella typhimurium generates an attenuated strain with vaccine potential. Microbiology 2009, 155, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Kulp, A.; Kuehn, M.J. Biological functions and biogenesis of secreted bacterial outer membrane vesicles. Annu. Rev. Microbiol. 2010, 64, 163–184. [Google Scholar] [CrossRef]
- Camacho, A.I.; de Souza, J.; Sanchez-Gomez, S.; Pardo-Ros, M.; Irache, J.M.; Gamazo, C. Mucosal immunization with Shigella flexneri outer membrane vesicles induced protection in mice. Vaccine 2011, 29, 8222–8229. [Google Scholar] [CrossRef]
- Micoli, F.; Rondini, S.; Alfini, R.; Lanzilao, L.; Necchi, F.; Negrea, A.; Rossi, O.; Brandt, C.; Clare, S.; Mastroeni, P.; et al. Comparative immunogenicity and efficacy of equivalent outer membrane vesicle and glycoconjugate vaccines against nontyphoidal Salmonella. Proc. Natl. Acad. Sci. USA 2018, 115, 10428–10433. [Google Scholar] [CrossRef]
- Mitra, S.; Sinha, R.; Mitobe, J.; Koley, H. Development of a cost-effective vaccine candidate with outer membrane vesicles of a tolA-disrupted Shigella boydii strain. Vaccine 2016, 34, 1839–1846. [Google Scholar] [CrossRef]
- Turner, L.; Praszkier, J.; Hutton, M.L.; Steer, D.; Ramm, G.; Kaparakis-Liaskos, M.; Ferrero, R.L. Increased Outer Membrane Vesicle Formation in a Helicobacter pylori tolB Mutant. Helicobacter 2015, 20, 269–283. [Google Scholar] [CrossRef] [PubMed]
- Takaki, K.; Tahara, Y.O.; Nakamichi, N.; Hasegawa, Y.; Shintani, M.; Ohkuma, M.; Miyata, M.; Futamata, H.; Tashiro, Y. Multilamellar and Multivesicular Outer Membrane Vesicles Produced by a Buttiauxella agrestis tolB Mutant. Appl. Environ. Microbiol. 2020, 86, e01131-20. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.D.; Bagchi, A.; Warren, H.S.; Tehan, M.M.; Trigilio, J.A.; Beasley-Topliffe, L.K.; Tesini, B.L.; Lazzaroni, J.C.; Fenton, M.J.; Hellman, J. Bacterial peptidoglycan-associated lipoprotein: A naturally occurring toll-like receptor 2 agonist that is shed into serum and has synergy with lipopolysaccharide. J. Infect. Dis. 2005, 191, 939–948. [Google Scholar] [CrossRef]
- Shim, H.K.; Kim, J.Y.; Kim, M.J.; Sim, H.S.; Park, D.W.; Sohn, J.W.; Kim, M.J. Legionella lipoprotein activates toll-like receptor 2 and induces cytokine production and expression of costimulatory molecules in peritoneal macrophages. Exp. Mol. Med. 2009, 41, 687–694. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mobarez, A.M.; Rajabi, R.A.; Salmanian, A.H.; Khoramabadi, N.; Hosseini Doust, S.R. Induction of protective immunity by recombinant peptidoglycan associated lipoprotein (rPAL) protein of Legionella pneumophila in a BALB/c mouse model. Microb. Pathog. 2019, 128, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Sin, J.I.; Kim, M.J. CD8(+) T Cells Directed Against a Peptide Epitope Derived from Peptidoglycan-Associated Lipoprotein of Legionella pneumophila Confer Disease Protection. Front. Immunol. 2020, 11, 604413. [Google Scholar] [CrossRef] [PubMed]
- McMahon, M.; Murphy, T.F.; Kyd, J.; Thanavala, Y. Role of an immunodominant T cell epitope of the P6 protein of nontypeable Haemophilus influenzae in murine protective immunity. Vaccine 2005, 23, 3590–3596. [Google Scholar] [CrossRef]
- Spinola, S.M.; Griffiths, G.E.; Bogdan, J.; Menegus, M.A. Characterization of an 18,000-molecular-weight outer membrane protein of Haemophilus ducreyi that contains a conserved surface-exposed epitope. Infect. Immun. 1992, 60, 385–391. [Google Scholar] [CrossRef]
- Yoon, W.S.; Park, S.H.; Park, Y.K.; Park, S.C.; Sin, J.I.; Kim, M.J. Comparison of responses elicited by immunization with a Legionella species common lipoprotein delivered as naked DNA or recombinant protein. DNA Cell Biol. 2002, 21, 99–107. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, Z.; Dong, Y.; Chen, Y. Recombinant PAL/PilE/FlaA DNA vaccine provides protective immunity against Legionella pneumophila in BALB/c mice. BMC Biotechnol. 2020, 20, 28. [Google Scholar] [CrossRef]
- Lei, L.; Yang, F.; Zou, J.; Jing, H.; Zhang, J.; Xu, W.; Zou, Q.; Zhang, J.; Wang, X. DNA vaccine encoding OmpA and Pal from Acinetobacter baumannii efficiently protects mice against pulmonary infection. Mol. Biol. Rep. 2019, 46, 5397–5408. [Google Scholar] [CrossRef] [PubMed]
- Hirakawa, H.; Suzue, K.; Takita, A.; Tomita, H. Roles of OmpA in Type III Secretion System-Mediated Virulence of Enterohemorrhagic Escherichia coli. Pathogens 2021, 10, 1496. [Google Scholar] [CrossRef] [PubMed]
- Prasadarao, N.V.; Wass, C.A.; Weiser, J.N.; Stins, M.F.; Huang, S.H.; Kim, K.S. Outer membrane protein A of Escherichia coli contributes to invasion of brain microvascular endothelial cells. Infect. Immun. 1996, 64, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Sukumaran, S.K.; Shimada, H.; Prasadarao, N.V. Entry and intracellular replication of Escherichia coli K1 in macrophages require expression of outer membrane protein A. Infect. Immun. 2003, 71, 5951–5961. [Google Scholar] [CrossRef]
- Nicholson, T.F.; Watts, K.M.; Hunstad, D.A. OmpA of uropathogenic Escherichia coli promotes postinvasion pathogenesis of cystitis. Infect. Immun. 2009, 77, 5245–5251. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hirakawa, H.; Suzue, K.; Tomita, H. Roles of the Tol/Pal System in Bacterial Pathogenesis and Its Application to Antibacterial Therapy. Vaccines 2022, 10, 422. https://doi.org/10.3390/vaccines10030422
Hirakawa H, Suzue K, Tomita H. Roles of the Tol/Pal System in Bacterial Pathogenesis and Its Application to Antibacterial Therapy. Vaccines. 2022; 10(3):422. https://doi.org/10.3390/vaccines10030422
Chicago/Turabian StyleHirakawa, Hidetada, Kazutomo Suzue, and Haruyoshi Tomita. 2022. "Roles of the Tol/Pal System in Bacterial Pathogenesis and Its Application to Antibacterial Therapy" Vaccines 10, no. 3: 422. https://doi.org/10.3390/vaccines10030422
APA StyleHirakawa, H., Suzue, K., & Tomita, H. (2022). Roles of the Tol/Pal System in Bacterial Pathogenesis and Its Application to Antibacterial Therapy. Vaccines, 10(3), 422. https://doi.org/10.3390/vaccines10030422