
Targeting Secreted Protease/Anti-Protease Balance as a Vaccine Strategy against the Helminth Fasciola hepatica

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statements
2.2. Production of Functional Recombinant F. hepatica Stefins and Kunitz-Type Inhibitor
2.3. Whole Mount NEJ Immunolocalisation of FhStf-1, FhStf-2 and FhStf-3 by Confocal Microscopy
2.4. Inhibition of Cysteine Peptidase Activity in F. hepatica NEJ and Adult Secreted/Excreted Proteins (ES) by rFhStf-1, rFhStf-2 and rFhStf-3
2.5. Vaccine Trial Design
2.6. Haematological Analysis and Liver Enzyme Assays
2.7. Egg Hatch Assay (EHA)
2.8. Analysis of Anti-F. hepatica Antigen Specific Antibodies (Total IgG) in Sheep Sera Samples by ELISA
2.9. Analysis of Anti-F. hepatica Antigen Specific IgG1 and IgG2 Isotypes in Sheep Sera Samples by ELISA
2.10. Isolation of Ovine Peripheral Blood Mononuclear Cell (PBMC) and Lymphocyte Stimulation Assays (LSA)
2.11. Statistical Analysis
3. Results
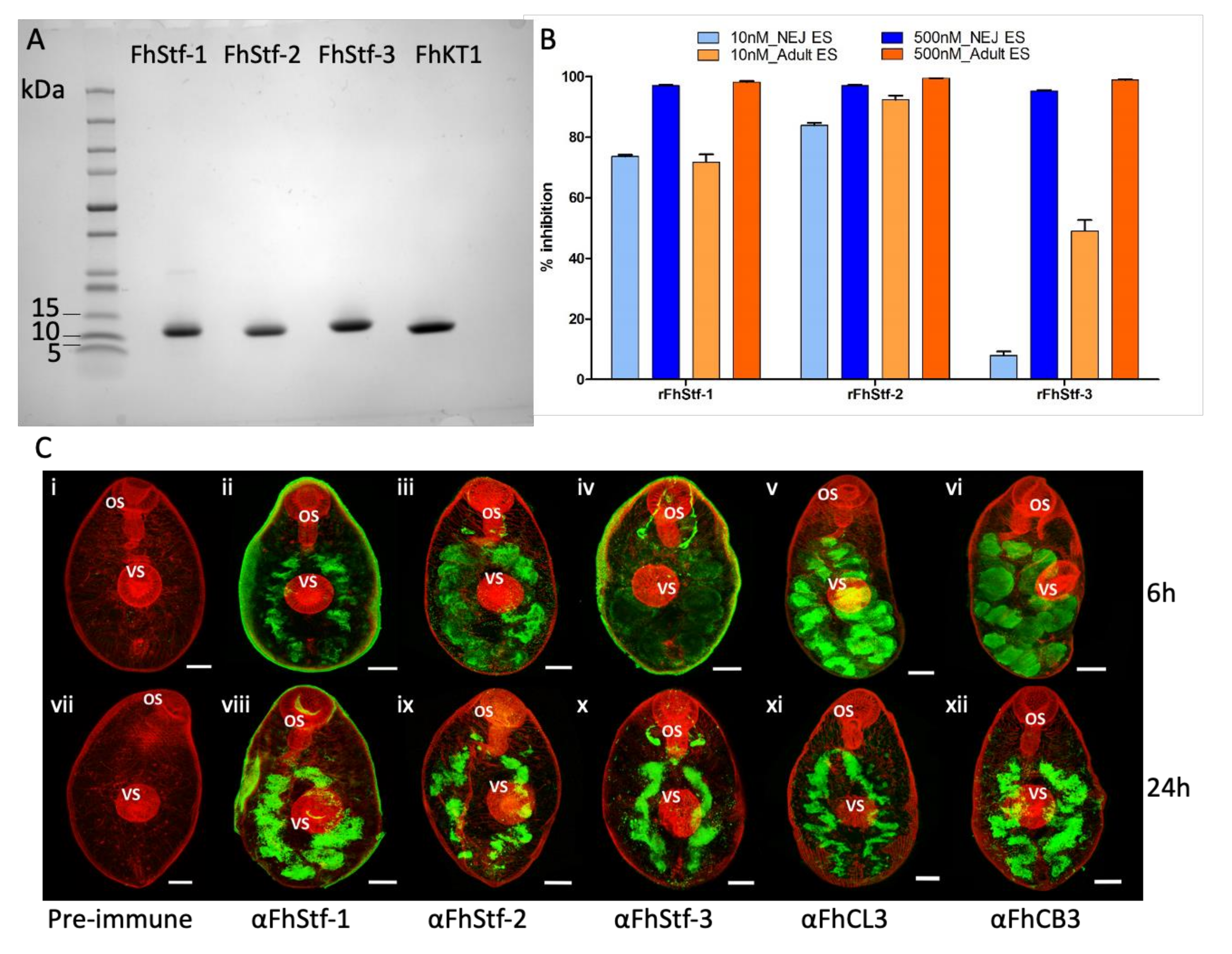
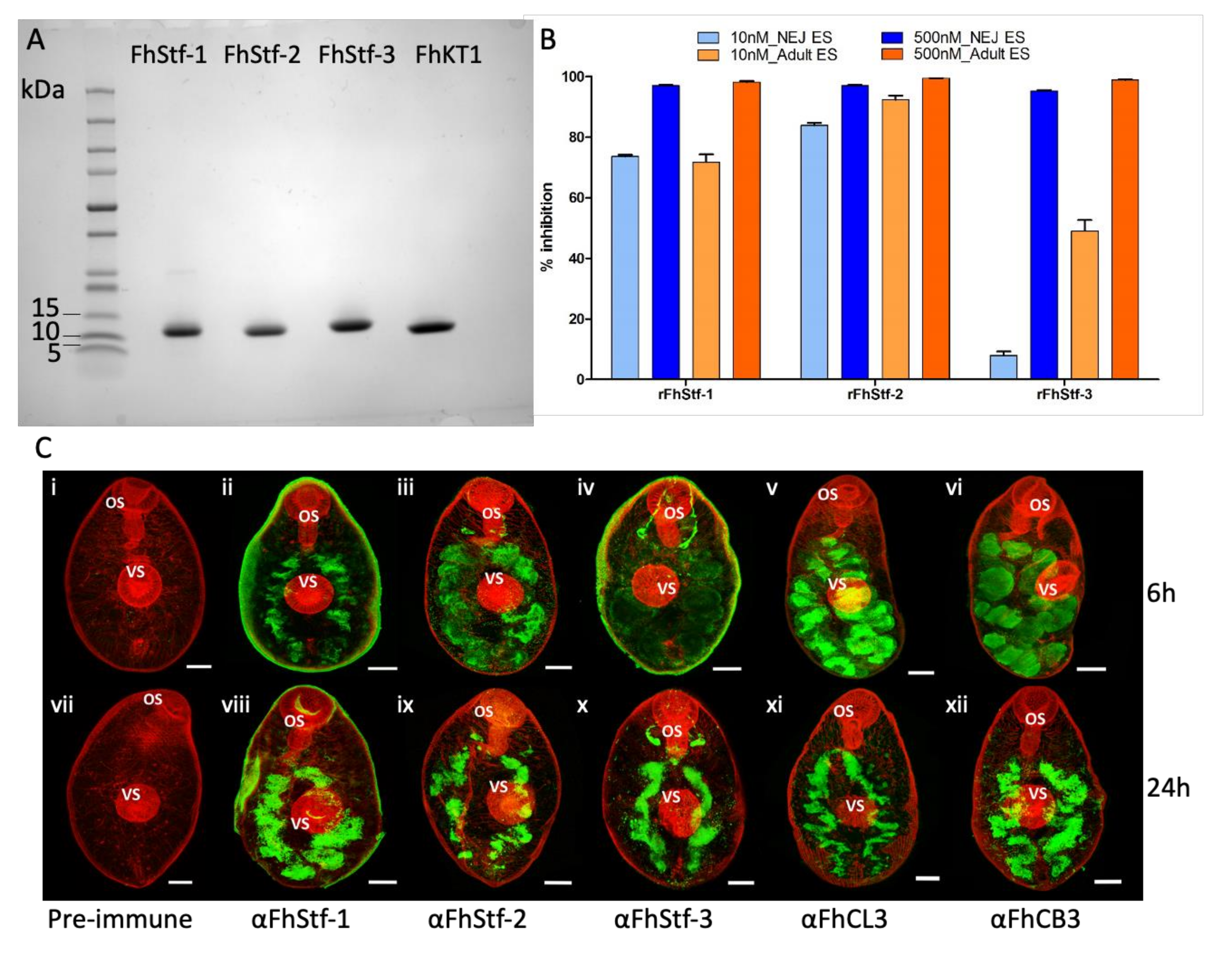
3.1. Recombinant Production of Functionally Active Proteins
3.1.1. Recombinant Expression of Three F. hepatica Stefins and Kunitz-Type Inhibitor
3.1.2. The Recombinant F. hepatica Stefins and Kunitz-Type Inhibitor Are Potent Inhibitors of Parasite Cysteine Peptidases
3.1.3. Immunolocalisation Studies
3.2. Vaccine Trials
3.2.1. Assessment of Protection Based on Total Adult Fluke Enumeration and Egg Viability
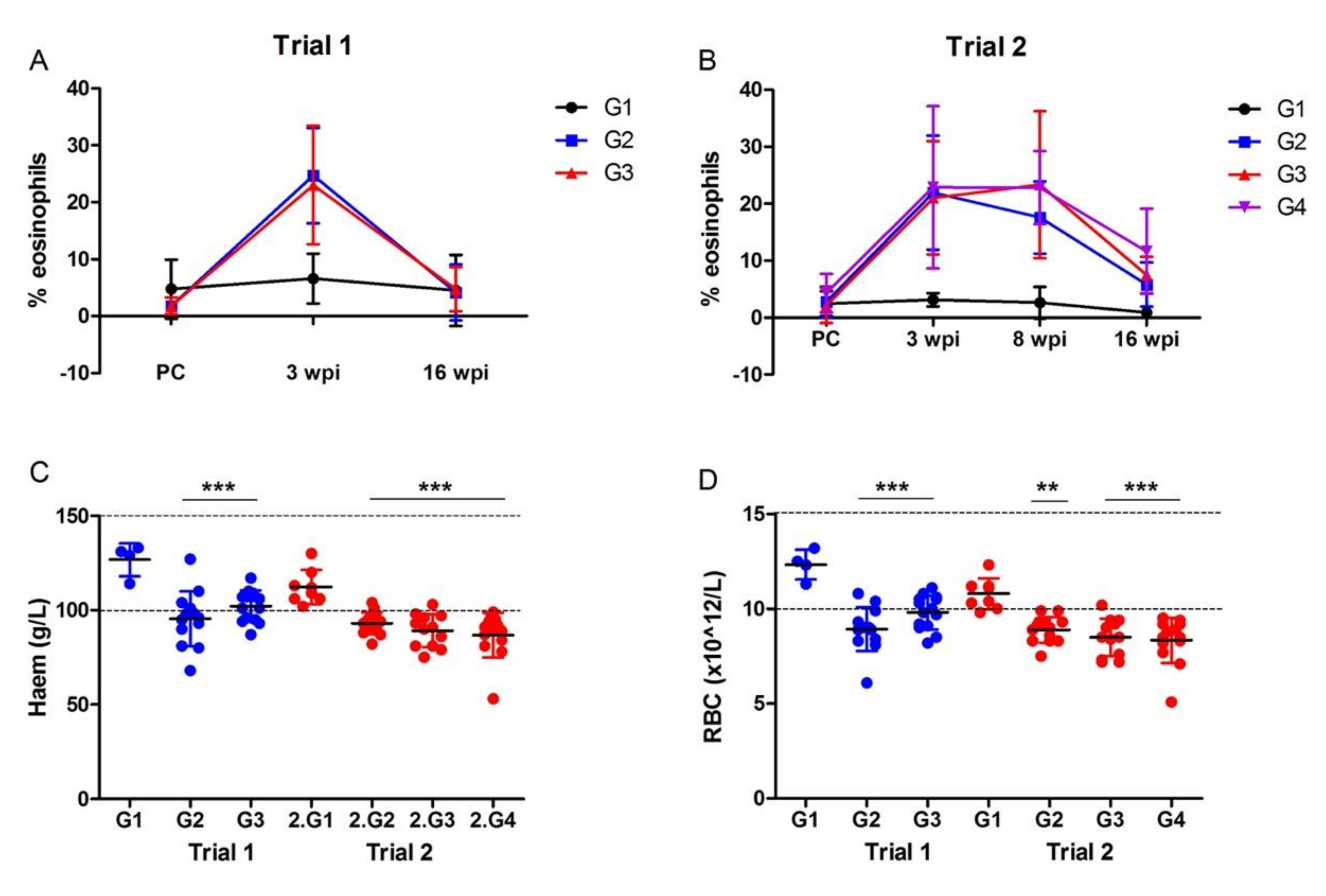
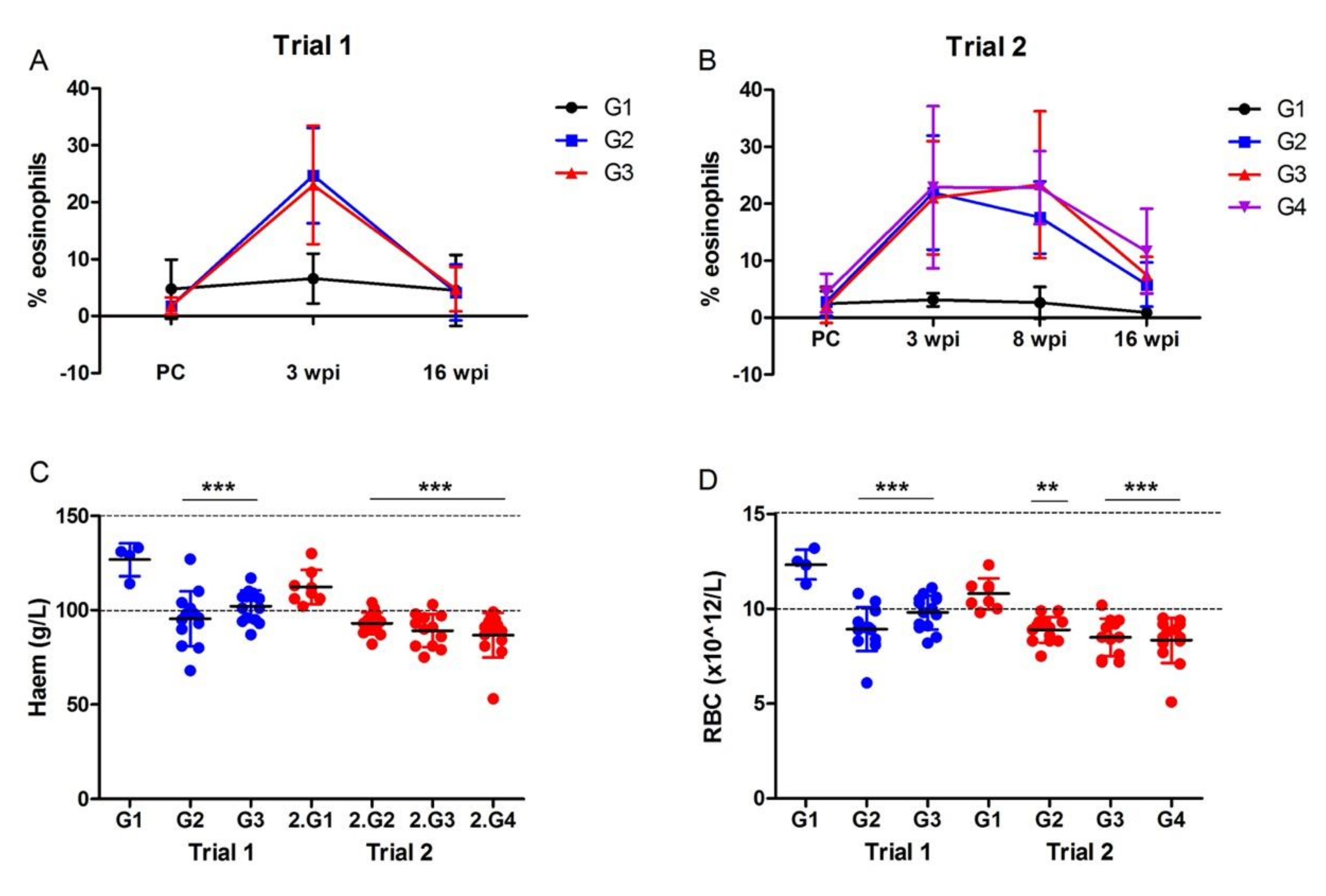
3.2.2. Haematological Analysis
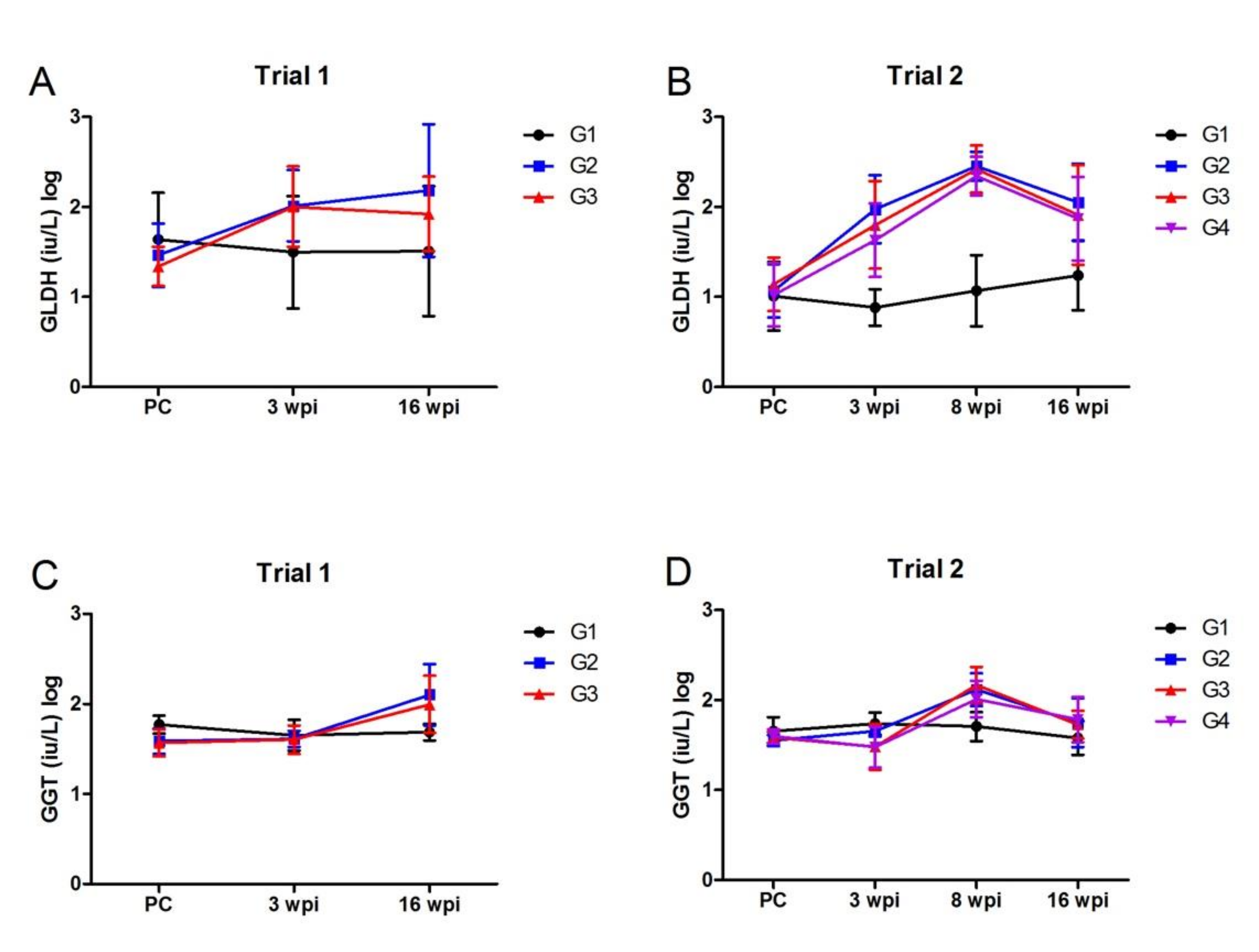
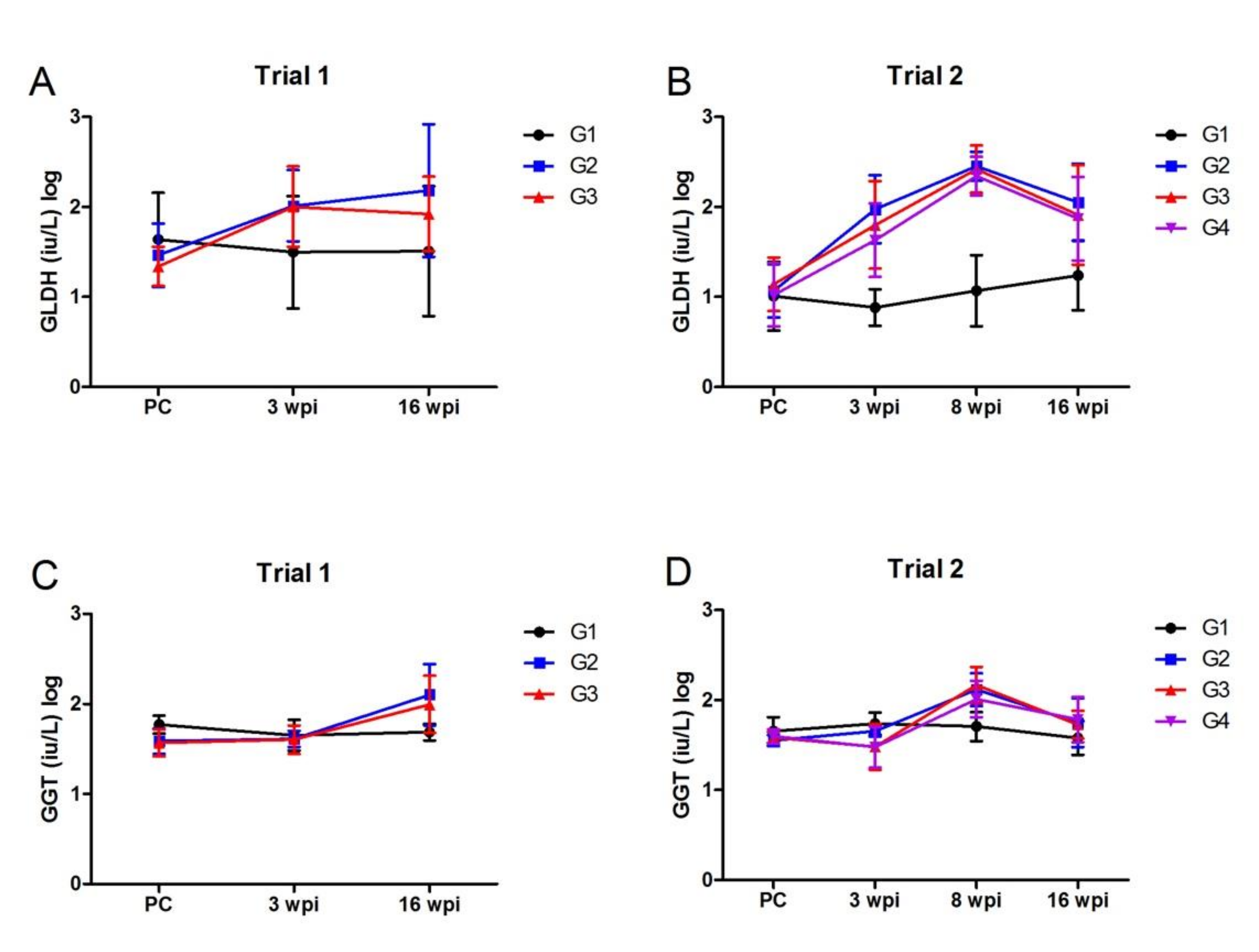
3.2.3. Liver Enzyme Assays
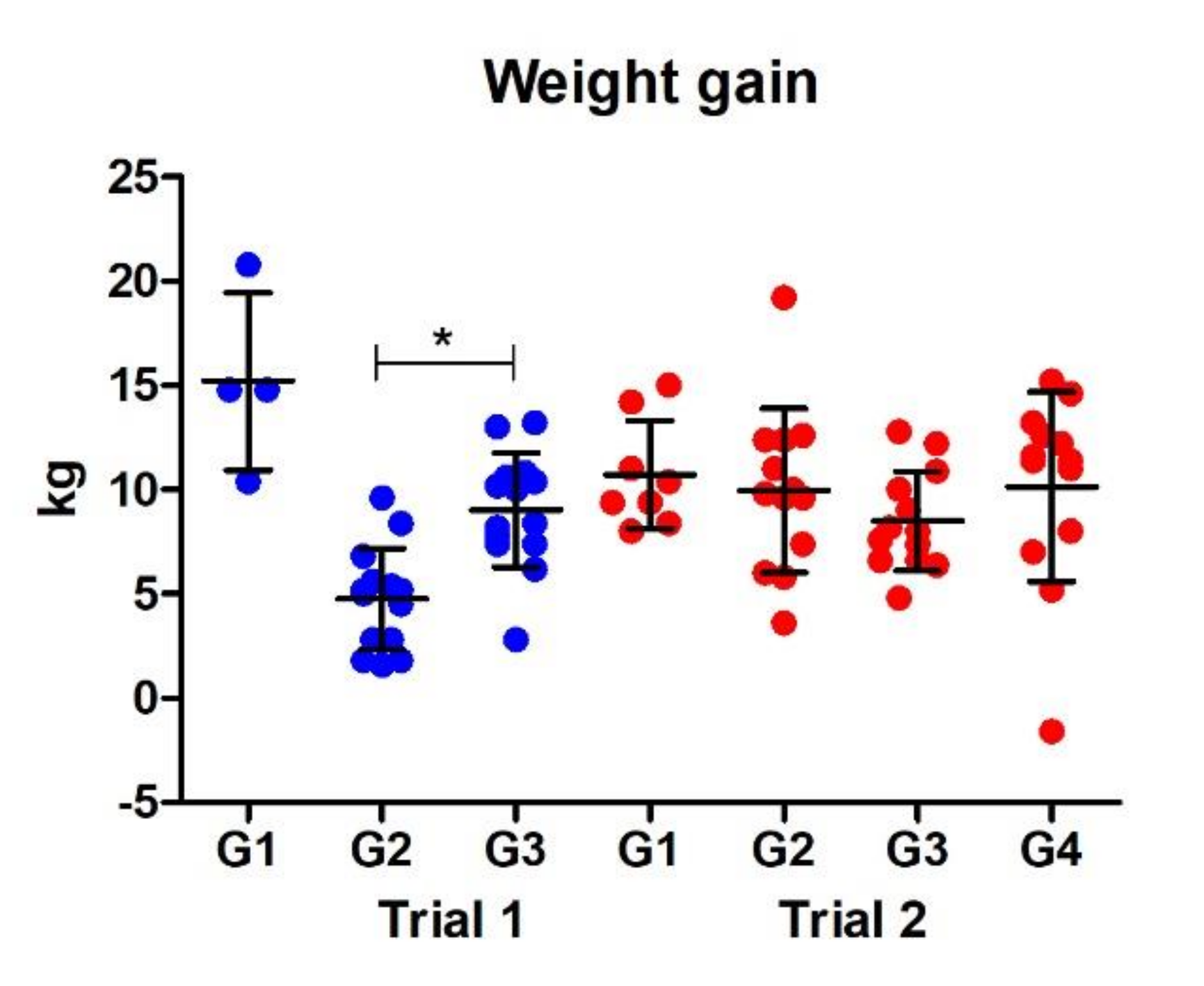
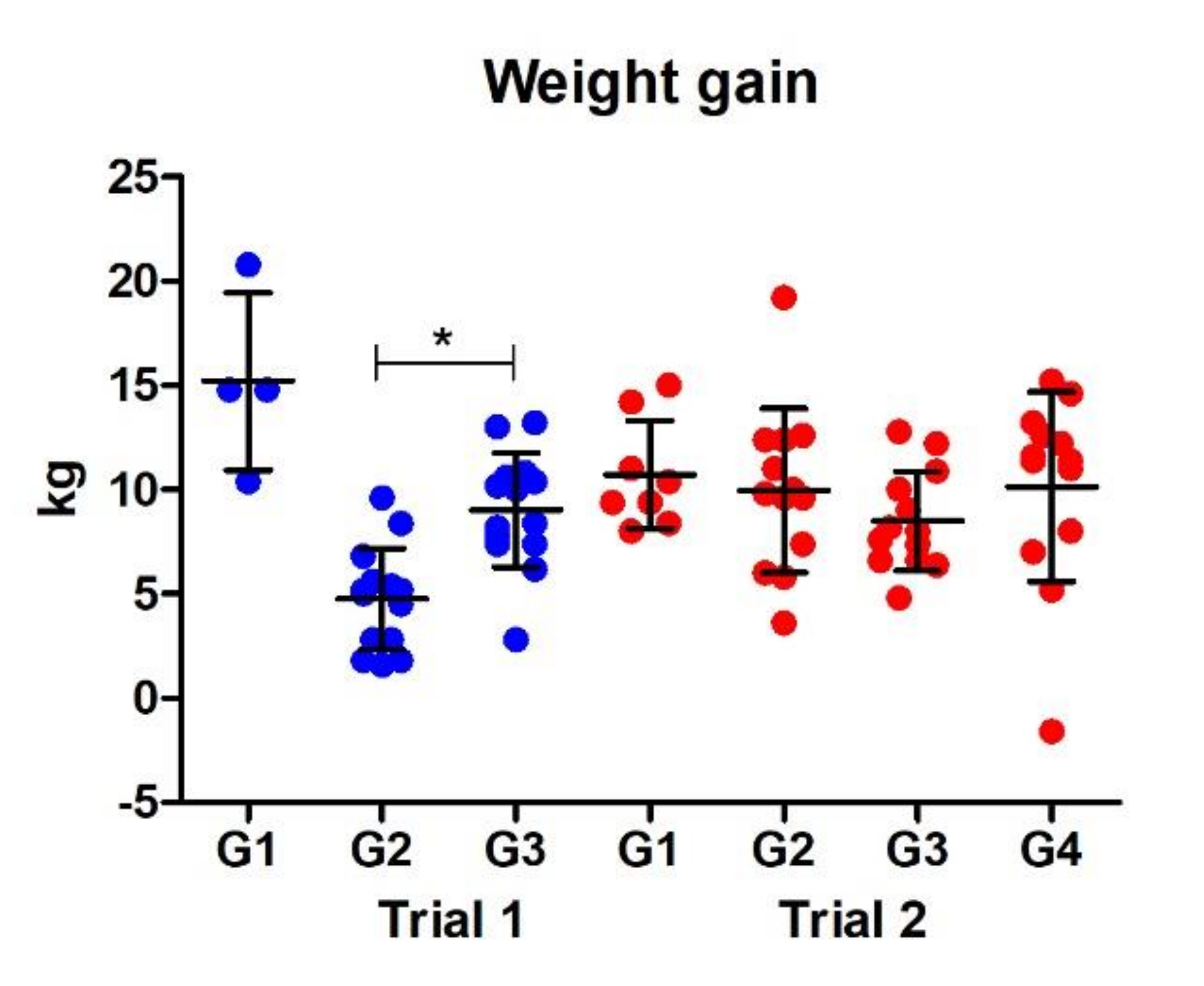
3.2.4. Weight Gain
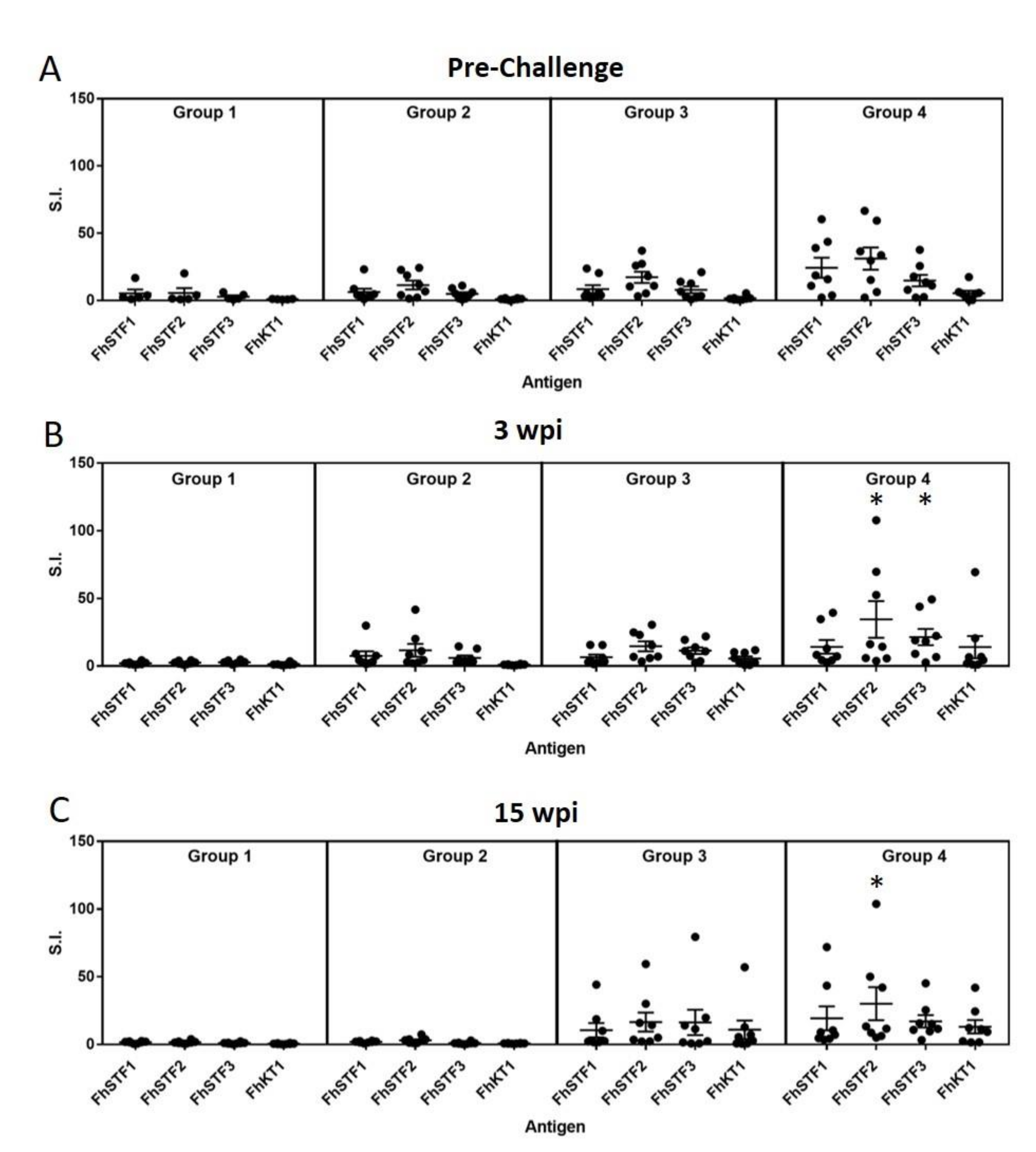
3.3. Analysis of Immunogenicity of the Vaccine Antigens by Lymphocyte Stimulation Assays
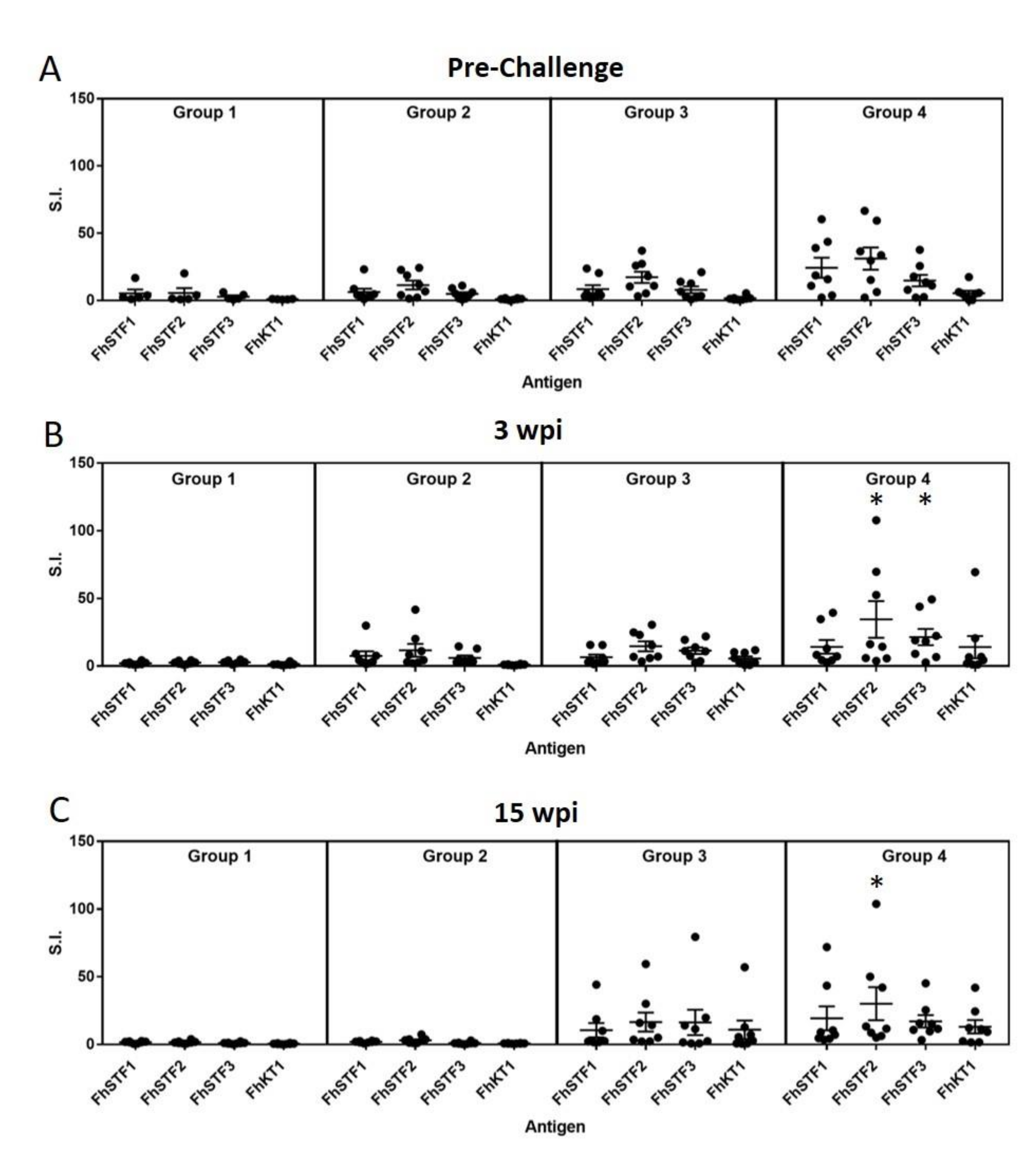
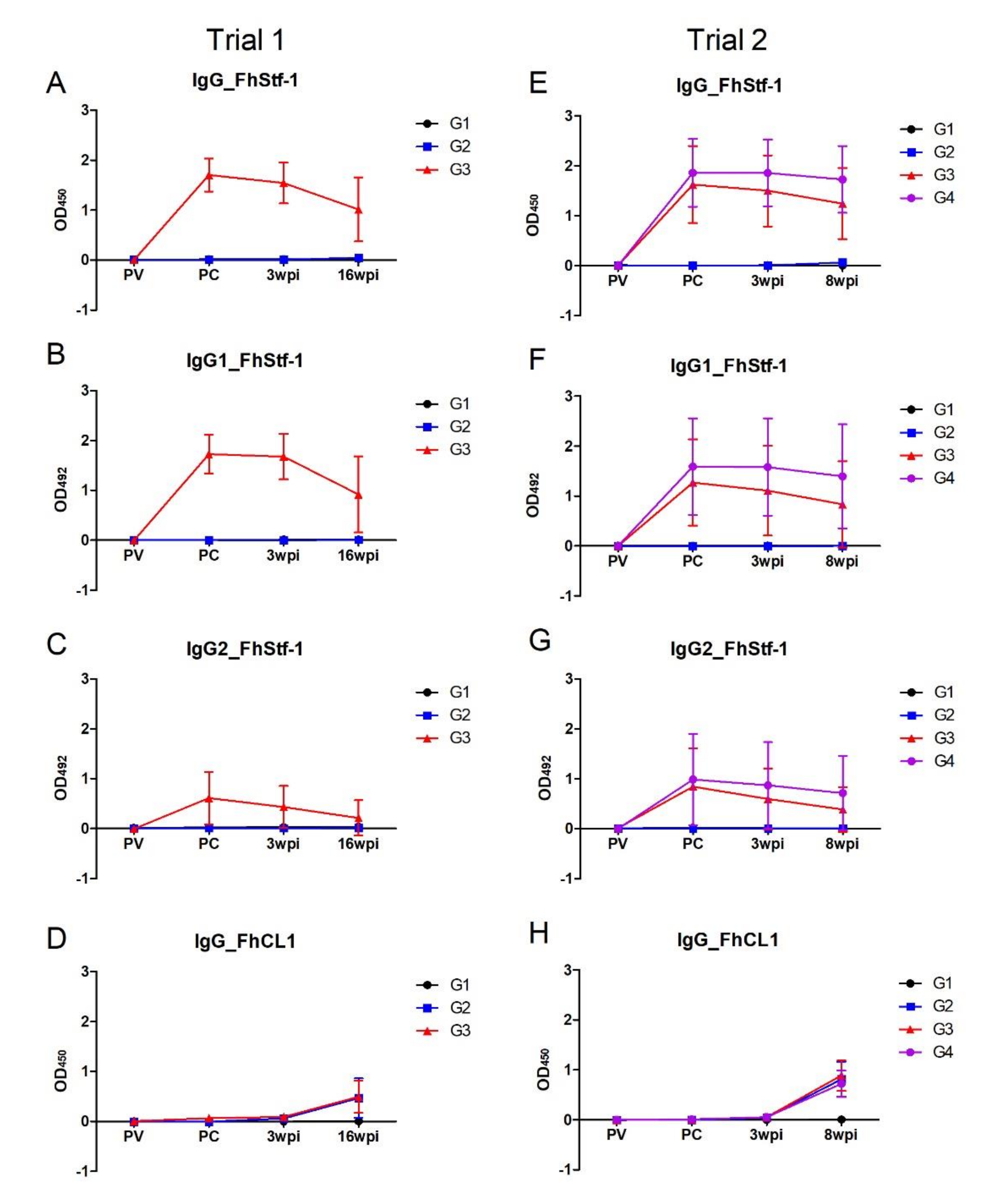
3.4. Analysis of Immunogenicity of the Vaccine Antigens by ELISA
3.5. Correlation Analysis
- Vaccination was associated with an average weight gain of 3.5 kg and increased haemoglobin levels of 7.18 g/L.
- The IgG2 antibodies elicited by vaccination were associated with average increased haemoglobin levels of 13.44 g/L.
- Infection with the Italian liver fluke isolate, which comprised of on average 42 more adult parasites than infections with the South Gloucester isolate, was associated with an average weight loss of 4.4 kg.
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Charlier, J.; Rinaldi, L.; Musella, V.; Ploeger, H.W.; Chartier, C.; Vineer, H.R.; Hinney, B.; von Samson-Himmelstjerna, G.; Băcescu, B.; Mickiewicz, M.; et al. Initial assessment of the economic burden of major parasitic helminth infections to the ruminant livestock industry in Europe. Prev. Vet. Med. 2020, 182, 105103. [Google Scholar] [CrossRef]
- Morgan, E.R.; Charlier, J.; Hendrickx, G.; Biggeri, A.; Catalan, D.; von Samson-Himmelstjerna, G.; Demeler, J.; Müller, E.; Van Dijk, J.; Kenyon, F.; et al. Global Change and Helminth Infections in Grazing Ruminants in Europe: Impacts, Trends and Sustainable Solutions. Agriculture 2013, 3, 484–502. [Google Scholar] [CrossRef] [Green Version]
- Mehmood, K.; Zhang, H.; Sabir, A.J.; Abbas, R.Z.; Ijaz, M.; Durrani, A.Z.; Saleem, M.H.; Ur Rehman, M.; Iqbal, M.K.; Wang, Y.; et al. A review on epidemiology, global prevalence and economical losses of fasciolosis in ruminants. Microb. Pathog. 2017, 109, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Animal Health Ireland. Liver Fluke—The Facts. Available online: https://animalhealthireland.ie/?ahi-publication=liver-fluke-the-facts (accessed on 1 November 2021).
- Hayward, A.D.; Skuce, P.J.; McNeilly, T.N. The influence of liver fluke infection on production in sheep and cattle: A meta-analysis. Int. J. Parasitol. 2021, 51, 913–924. [Google Scholar] [CrossRef] [PubMed]
- Castro-Hermida, J.A.; González-Warleta, M.; Martínez-Sernández, V.; Ubeira, F.M.; Mezo, M. Current challenges for fasciolicide treatment in ruminant livestock. Trends Parasitol. 2021, 37, 430–444. [Google Scholar] [CrossRef]
- Overend, D.J.; Bowen, F.L. Resistance of Fasciola hepatica to triclabendazole. Aust. Vet. J. 1995, 72, 275–276. [Google Scholar] [CrossRef]
- Fairweather, I.; Brennan, G.P.; Hanna, R.E.B.; Robinson, M.W.; Skuce, P.J. Drug resistance in liver flukes. Int. J. Parasitol. Drugs Drug Resist. 2020, 12, 39–59. [Google Scholar] [CrossRef] [PubMed]
- Dalton, J.P.; Mulcahy, G. Parasite vaccines—A reality? Vet. Parasitol. 2001, 98, 149–167. [Google Scholar] [CrossRef]
- Perera, D.J.; Ndao, M. Promising technologies in the field of helminth vaccines. Front. Immunol. 2021, 12, 711650. [Google Scholar] [CrossRef]
- Cwiklinski, K.; De Marco Verissimo, C.; McVeigh, P.; Donnelly, S.; Dalton, J.P. Applying ‘omics’ technologies to understand Fasciola spp. biology. In Fasciolosis, 2nd ed.; Dalton, J.P., Ed.; CABI Publishing: Wallingford, UK, 2021; pp. 338–378. [Google Scholar]
- Cwiklinski, K.; Jewhurst, H.; McVeigh, P.; Barbour, T.; Maule, A.G.; Tort, J.; O’Neill, S.M.; Robinson, M.W.; Donnelly, S.; Dalton, J.P. Infection by the helminth parasite Fasciola hepatica requires rapid regulation of metabolic, virulence, and invasive factors to adjust to its mammalian host. Mol. Cell. Proteom. 2018, 17, 792–809. [Google Scholar] [CrossRef] [Green Version]
- Cwiklinski, K.; Robinson, M.W.; Donnelly, S.; Dalton, J.P. Complementary transcriptomic and proteomic analyses reveal the cellular and molecular processes that drive growth and development of Fasciola hepatica in the host liver. BMC Genom. 2021, 22, 46. [Google Scholar] [CrossRef]
- Dalton, J.P.; O’Neill, S.; Stack, C.; Collins, P.; Walshe, A.; Sekiya, M.; Doyle, S.; Mulcahy, G.; Hoyle, D.; Khaznadji, E.; et al. Fasciola hepatica cathepsin L-like proteases: Biology, function, and potential in the development of first generation liver fluke vaccines. Int. J. Parasitol. 2003, 33, 1173–1181. [Google Scholar] [CrossRef]
- Robinson, M.W.; Corvo, I.; Jones, P.M.; George, A.M.; Padula, M.P.; To, J.; Cancela, M.; Rinaldi, G.; Tort, J.F.; Roche, L.; et al. Collagenolytic activities of the major secreted cathepsin L peptidases involved in the virulence of the helminth pathogen, Fasciola hepatica. PLoS NTD 2011, 5, e1012. [Google Scholar] [CrossRef] [Green Version]
- Cwiklinski, K.; Donnelly, S.; Drysdale, O.; Jewhurst, H.; Smith, D.; De Marco Verissimo, C.; Pritsch, I.C.; O’Neill, S.; Dalton, J.P.; Robinson, M.W. The cathepsin-like cysteine peptidases of trematodes of the genus Fasciola. Adv. Parasitol. 2019, 104, 113–164. [Google Scholar] [PubMed]
- Barbour, T.; Cwiklinski, K.; Lalor, R.; Dalton, J.P.; De Marco Verissimo, C. The zoonotic helminth parasite Fasciola hepatica: Virulence-associated cathepsin B and cathepsin L cysteine peptidases secreted by infective newly excysted juveniles (NEJ). Animals 2021, 11, 3495. [Google Scholar] [CrossRef]
- Caffrey, C.R.; McKerrow, J.H.; Salter, J.P.; Sajid, M. Blood ‘n’ guts: An update on schistosome digestive peptidases. Trends Parasitol. 2004, 20, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Dalton, J.P.; Caffrey, C.R.; Sajid, M.; Stack, C.; Donnelly, S.; Loukas, A.; Don, T.; McKerrow, J.; Halton, D.W.; Brindley, P.J. Proteases in trematode biology. In Parasitic Flatworms: Molecular Biology, Biochemistry, Immunology and Physiology; Maule, A.G., Marks, N.J., Eds.; CAB International: Wallingford, UK, 2006; pp. 348–368. [Google Scholar]
- Tort, J.; Brindley, P.J.; Knox, D.; Wolfe, K.H.; Dalton, J.P. Proteinases and associated genes of parasitic helminths. Adv. Parasitol. 1999, 43, 161–266. [Google Scholar]
- Smith, D.; Cwiklinski, K.; Jewhurst, H.; Tikhonova, I.G.; Dalton, J.P. An atypical and functionally diverse family of Kunitz-type cysteine/serine proteinase inhibitors secreted by the helminth parasite Fasciola hepatica. Sci. Rep. 2020, 10, 20657. [Google Scholar] [CrossRef]
- Smith, D.; Tikhonova, I.G.; Jewhurst, H.L.; Drysdale, O.C.; Dvořák, J.; Robinson, M.W.; Cwiklinski, K.; Dalton, J.P. Unexpected Activity of a Novel Kunitz-type Inhibitor: Inhibition of cysteine proteases but not serine proteases. J. Biol. Chem. 2016, 291, 19220–19234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cancela, M.; Corvo, I.; da Silva, E.; Teichmann, A.; Roche, L.; Díaz, A.; Tort, J.F.; Ferreira, H.B.; Zaha, A. Functional characterization of single-domain cystatin-like cysteine proteinase inhibitors expressed by the trematode Fasciola hepatica. Parasitology 2017, 144, 1695–1707. [Google Scholar] [CrossRef]
- Collins, P.R.; Stack, C.M.; O’Neill, S.M.; Doyle, S.; Ryan, T.; Brennan, G.P.; Mousley, A.; Stewart, M.; Maule, A.G.; Dalton, J.P.; et al. Cathepsin L1, the major protease involved in liver fluke (Fasciola hepatica) virulence: Propetide cleavage sites and autoactivation of the zymogen secreted from gastrodermal cells. J. Biol. Chem. 2004, 279, 17038–17046. [Google Scholar] [CrossRef] [Green Version]
- López Corrales, J.; Cwiklinski, K.; De Marco Verissimo, C.; Dorey, A.; Lalor, R.; Jewhurst, H.; McEvoy, A.; Diskin, M.; Duffy, C.; Cosby, S.L.; et al. Diagnosis of sheep fasciolosis caused by Fasciola hepatica using cathepsin L enzyme-linked immunosorbent assays (ELISA). Vet. Parasitol. 2021, 298, 109517. [Google Scholar] [CrossRef] [PubMed]
- Dalton, J.P.; Tom, T.D.; Strand, M. Fasciola hepatica: Comparison of immature and mature immunoreactive glycoproteins. Parasite Immunol. 1985, 7, 643–657. [Google Scholar] [CrossRef]
- Robinson, M.W.; Menon, R.; Donnelly, S.M.; Dalton, J.P.; Ranganathan, S. An integrated transcriptomics and proteomics analysis of the secretome of the helminth pathogen Fasciola hepatica: Proteins associated with invasion and infection of the mammalian host. Mol. Cell. Proteomics. 2009, 8, 1891–1907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, A.; Cwiklinski, K.; Lalor, R.; O’Connell, B.; Robinson, M.W.; Gerlach, J.; Joshi, L.; Kilcoyne, M.; Dalton, J.P.; O’Neill, S.M. Fasciola hepatica Extracellular Vesicles isolated from excretory-secretory products using a gravity flow method modulate dendritic cell phenotype and activity. PLoS NTD 2020, 14, e0008626. [Google Scholar] [CrossRef]
- Toet, H.; Piedrafita, D.M.; Spithill, T.W. Liver fluke vaccines in ruminants: Strategies, progress and future opportunities. Int. J. Parasitol. 2014, 44, 915–927. [Google Scholar] [CrossRef] [PubMed]
- Fairweather, I.; McShane, D.D.; Shaw, L.; Ellison, S.E.; O’Hagan, N.T.; York, E.A.; Trudgett, A.; Brennan, G.P. Development of an egg hatch assay for the diagnosis of triclabendazole resistance in Fasciola hepatica: Proof of concept. Vet. Parasitol. 2012, 183, 249–259. [Google Scholar] [CrossRef] [PubMed]
- López-Otín, C.; Bond, J.S. Proteases: Multifunctional enzymes in life and disease. J. Biol. Chem. 2008, 283, 30433–30437. [Google Scholar] [CrossRef] [Green Version]
- Dey, T.; Kalita, J.; Weldon, S.; Taggart, C.C. Proteases and their inhibitors in chronic obstructive pulmonary disease. J. Clin. Med. 2018, 7, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eatemadi, A.; Aiyelabegan, H.T.; Negahdari, B.; Mazlomi, M.A.; Daraee, H.; Daraee, N.; Eatemadi, R.; Sadroddiny, E. Role of protease and protease inhibitors in cancer pathogenesis and treatment. Biomed. Pharmacother. 2017, 86, 221–231. [Google Scholar] [CrossRef]
- Reiser, J.; Adair, B.; Reinheckel, T. Specialized roles for cysteine cathepsins in health and disease. J. Clin. Investig. 2010, 120, 3421–3431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleh, Y.; Wnukiewicz, J.; Andrzejak, R.; Trziszka, T.; Siewinski, M.; Ziolkowski, P.; Kopec, W. Cathepsin B and cysteine protease inhibitors in human tongue cancer: Correlation with tumor staging and in vitro inhibition of cathepsin B by chicken cystatin. J. Cancer Mol. 2006, 2, 67–72. [Google Scholar]
- Leto, G.; Sepporta, M.V. The potential of cystatin C as a predictive biomarker in breast cancer. Expert Rev. Anticancer. Ther. 2020, 20, 1049–1056. [Google Scholar] [CrossRef]
- Eide, T.J.; Jarvinen, M.; Hopsu-Havu, V.K.; Maltau, J.; Rinne, A. Immunolocalization of cystatin A in neoplastic, virus and inflammatory lesions of the uterine cervix. Acta Histochem. 1992, 93, 241–248. [Google Scholar] [CrossRef]
- Strojnik, T.; Zajc, I.; Bervar, A.; Židanik, B.; Golouh, R.; Kos, J.; Dolenc, V.; Lah, T. Cathepsin B and its inhibitor stefin A in brain tumors. Pflugers Arch. 2000, 439, R122–R123. [Google Scholar] [CrossRef]
- Zajc, I.; Sever, N.; Bervar, A.; Lan, T.T. Expression of cysteine peptidase cathepsin L and its inhibitors stefins A and B in relation to tumorigenicity of breast cancer cell lines. Cancer Lett. 2002, 187, 185–190. [Google Scholar] [CrossRef]
- Mirtti, T.; Alanen, K.; Kallajoki, M.; Rinne, A.; Söderström, K.O. Expression of cystatins, high molecular weight cytokeratin, and proliferation markers in prostatic adenocarcinoma and hyperplasia. Prostate 2003, 54, 290–298. [Google Scholar] [CrossRef]
- Spithill, T.W.; Toet, H.; Rathinasamy, V.; Zerna, G.; Swan, J.; Cameron, T.; Smooker, P.M.; Piedrafita, D.M.; Dempster, R.; Beddoe, T. Vaccines for Fasciola (Liver fluke): New thinking for an old problem. In Fasciolosis, 2nd ed.; Dalton, J.P., Ed.; CABI Publishing: Wallingford, UK, 2021; pp. 379–422. [Google Scholar]
- Dalton, J.P.; McGonigle, S.; Rolph, T.P.; Andrews, S.J. Induction of protective immunity in cattle against infection with Fasciola hepatica by vaccination with cathepsin L proteinases and with hemoglobin. Infect. Immun. 1996, 64, 5066–5074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wijffels, G.L.; Salvatore, L.; Dosen, M.; Waddington, J.; Wilson, L.; Thompson, C.; Campbell, N.; Sexton, J.; Wicker, J.; Bowen, F.; et al. Vaccination of sheep with purified cysteine proteinases of Fasciola hepatica decreases worm fecundity. Exp. Parasitol. 1994, 78, 132–148. [Google Scholar] [CrossRef] [PubMed]
- Wesołowska, A.; Kozak Ljunggren, M.; Jedlina, L.; Basałaj, K.; Legocki, A.; Wedrychowicz, H.; Kesik-Brodacka, M. A preliminary study of a lettuce-based edible vaccine expressing the cysteine proteinase of Fasciola hepatica for fasciolosis control in livestock. Front. Immunol. 2018, 9, 2592. [Google Scholar] [CrossRef]
- Golden, O.; Flynn, R.J.; Read, C.; Sekiya, M.; Donnelly, S.M.; Stack, C.; Dalton, J.P.; Mulcahy, G. Protection of cattle against a natural infection of Fasciola hepatica by vaccination with recombinant cathepsin L1 (rFhCL1). Vaccine 2010, 28, 5551–5557. [Google Scholar] [CrossRef]
- Zafra, R.; Buffoni, L.; Pérez-Caballero, R.; Molina-Hernández, V.; Ruiz-Campillo, M.T.; Pérez, J.; Martínez-Moreno, Á.; Martínez Moreno, F.J. Efficacy of a multivalent vaccine against Fasciola hepatica infection in sheep. Vet. Res. 2021, 52, 13. [Google Scholar] [CrossRef] [PubMed]
- Mulcahy, G.; O’Connor, F.; McGonigle, S.; Dowd, A.; Clery, D.G.; Andrews, S.J.; Dalton, J.P. Correlation of specific antibody titre and avidity with protection in cattle immunized against Fasciola hepatica. Vaccine 1998, 16, 932–939. [Google Scholar] [CrossRef]
- McCluskie, M.J.; Weeratna, R.D.; Davis, H.L. The potential of oligodeoxynucleotides as mucosal and parenteral adjuvants. Vaccine 2001, 19, 2657–2660. [Google Scholar] [CrossRef]
- Bode, C.; Zhao, G.; Steinhagen, F.; Kinjo, T.; Klinman, D.M. CpG DNA as a vaccine adjuvant. Expert Rev. Vaccines 2011, 10, 499–511. [Google Scholar] [CrossRef] [Green Version]
- Zafra, R.; Pérez-Écija, R.A.; Buffoni, L.; Moreno, P.; Bautista, M.J.; Martínez-Moreno, A.; Mulcahy, G.; Dalton, J.P.; Pérez, J. Early and late peritoneal and hepatic changes in goats immunized with recombinant cathepsin L1 and infected with Fasciola hepatica. J. Comp. Pathol. 2013, 148, 373–384. [Google Scholar] [CrossRef] [PubMed]
- O’Riordan, E.G.; Hanrahan, J.P. The effects of sex and castration on growth rate and carcass traits of lambs. Ir. J. Agric. Food Res. 1992, 31, 157–162. [Google Scholar]
- Mirderikvandi, M.; Masoudi, A.; Khaldari, M.; Bojarpour, M.; Nazari, P. The effect of some factors on growth performance of Lori Bakhtiari lambs. Livest. Res. Rural. Dev. 2016, 28, 212. [Google Scholar]
- Ghaderi–Zefrehei, M.; Safari, A.; Moridi, M.; Khanzadeh, H.; Rashidi Dehsaraei, A. Bayesian estimate of genetic parameters for growth traits in Lori Bakhtiari sheep. Trop. Anim. Health Prod. 2021, 53, 457. [Google Scholar] [CrossRef]
- Bunge, L.; Navajas, E.A.; Stevenson, L.; Lambe, N.R.; Maltin, C.A.; Simm, G.; Chang, K.C. Breed and sex effects on muscle fibre characterisitics in two contrasting sheep breeds: Scottish Blackface and Texel. Proc. Br. Soc. Anim. Sci. 2007, 2007, 111. [Google Scholar] [CrossRef]
- De Vargas Junior, F.M.; Martins, C.F.; dos Santos, P.; Ferreira, M.B.; de Almeida Ricardo, H.; Leão, A.G.; Fernandes, A.R.M.; Teixeira, A. The effect of sex and genotype on growth performance, feed efficiency, and carcass traits of local sheep group Pantaneiro and Texel or Santa Inês crossbred finished on feedlot. Trop. Anim. Health Prod. 2014, 46, 869–875. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trial | Group | Mean Fluke Burden ± SD | Range | Protection (%) | Length (cm) ± SD |
|---|---|---|---|---|---|
| 1 | G2 | 60.71 ± 12.43 | 38–78 | - | 1.71 ± 0.108 |
| 1 | G3 | 50.14 ± 16.27 | 29–81 | 17.4 | 1.62 ± 0.260 |
| 2 | G2 | 87.23 ± 24.63 | 49–143 | - | 2.26 ± 0.467 |
| 2 | G3 | 102.10 ± 24.32 | 65–145 | 0 | 2.14 ± 0.312 |
| 2 | G4 | 86.92 ± 22.86 | 46–124 | 0 | 2.12 ± 0.166 |
| Mean EPG ± SD | ||||
|---|---|---|---|---|
| Trial | Group | 12 wpi | 14 wpi | 16 wpi |
| 1 | G2 | 33.0 ± 17.6 | 38.3 ± 15.9 | 62.3 ± 30.7 |
| 1 | G3 | 32.4 ± 19.6 | 39.4 ± 20.9 | 61.4 ± 49.6 |
| 2 | G2 | 27.3 ± 13.0 | 56.7 ± 25.9 | 55.4 ± 26.8 |
| 2 | G3 | 24.4 ± 14.7 | 56.1 ± 40.5 | 64.4 ± 38.3 |
| 2 | G4 | 57.1 ± 30.2 | 60.0 ± 33.9 | 59.6 ± 26.6 |
| Trial | Group | Number of Eggs ± SD | Reduction (%) * |
|---|---|---|---|
| 1 | G2 | 2.06 × 105 ± 2.08 × 105 | - |
| 1 | G3 | 1.12 × 105 ± 1.17 × 105 | 46 |
| 2 | G2 | 1.01 × 106 ± 1.15 × 106 | - |
| 2 | G3 | 1.08 × 106 ± 1.07 × 106 | 0 |
| 2 | G4 | 0.58 × 106 ± 0.47 × 106 | 43 |
| Trial | Group | Embryonation (%) | Hatch Rate (%) | Protection (%) * |
|---|---|---|---|---|
| 1 | G2 | 52.7 ± 22.9 | 29.0 ± 18.7 | - |
| 1 | G3 | 49.1 ± 23.9 | 18.5 ± 12.2 | 36.2 |
| 2 | G2 | 58.8 ± 16.3 | 33.5 ± 13.0 | - |
| 2 | G3 | 60.0 ± 18.9 | 20.6 ± 10.8 | 38.5 |
| 2 | G4 | 67.2 ± 11.0 | 19.4 ± 6.8 | 42.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cwiklinski, K.; Drysdale, O.; López Corrales, J.; Corripio-Miyar, Y.; De Marco Verissimo, C.; Jewhurst, H.; Smith, D.; Lalor, R.; McNeilly, T.N.; Dalton, J.P. Targeting Secreted Protease/Anti-Protease Balance as a Vaccine Strategy against the Helminth Fasciola hepatica. Vaccines 2022, 10, 155. https://doi.org/10.3390/vaccines10020155
Cwiklinski K, Drysdale O, López Corrales J, Corripio-Miyar Y, De Marco Verissimo C, Jewhurst H, Smith D, Lalor R, McNeilly TN, Dalton JP. Targeting Secreted Protease/Anti-Protease Balance as a Vaccine Strategy against the Helminth Fasciola hepatica. Vaccines. 2022; 10(2):155. https://doi.org/10.3390/vaccines10020155
Chicago/Turabian StyleCwiklinski, Krystyna, Orla Drysdale, Jesús López Corrales, Yolanda Corripio-Miyar, Carolina De Marco Verissimo, Heather Jewhurst, David Smith, Richard Lalor, Tom N. McNeilly, and John P. Dalton. 2022. "Targeting Secreted Protease/Anti-Protease Balance as a Vaccine Strategy against the Helminth Fasciola hepatica" Vaccines 10, no. 2: 155. https://doi.org/10.3390/vaccines10020155
APA StyleCwiklinski, K., Drysdale, O., López Corrales, J., Corripio-Miyar, Y., De Marco Verissimo, C., Jewhurst, H., Smith, D., Lalor, R., McNeilly, T. N., & Dalton, J. P. (2022). Targeting Secreted Protease/Anti-Protease Balance as a Vaccine Strategy against the Helminth Fasciola hepatica. Vaccines, 10(2), 155. https://doi.org/10.3390/vaccines10020155