
Efficacy and Safety of Native and Recombinant Zona Pellucida Immunocontraceptive Vaccines Formulated with Non-Freund’s Adjuvants in Donkeys
 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Management
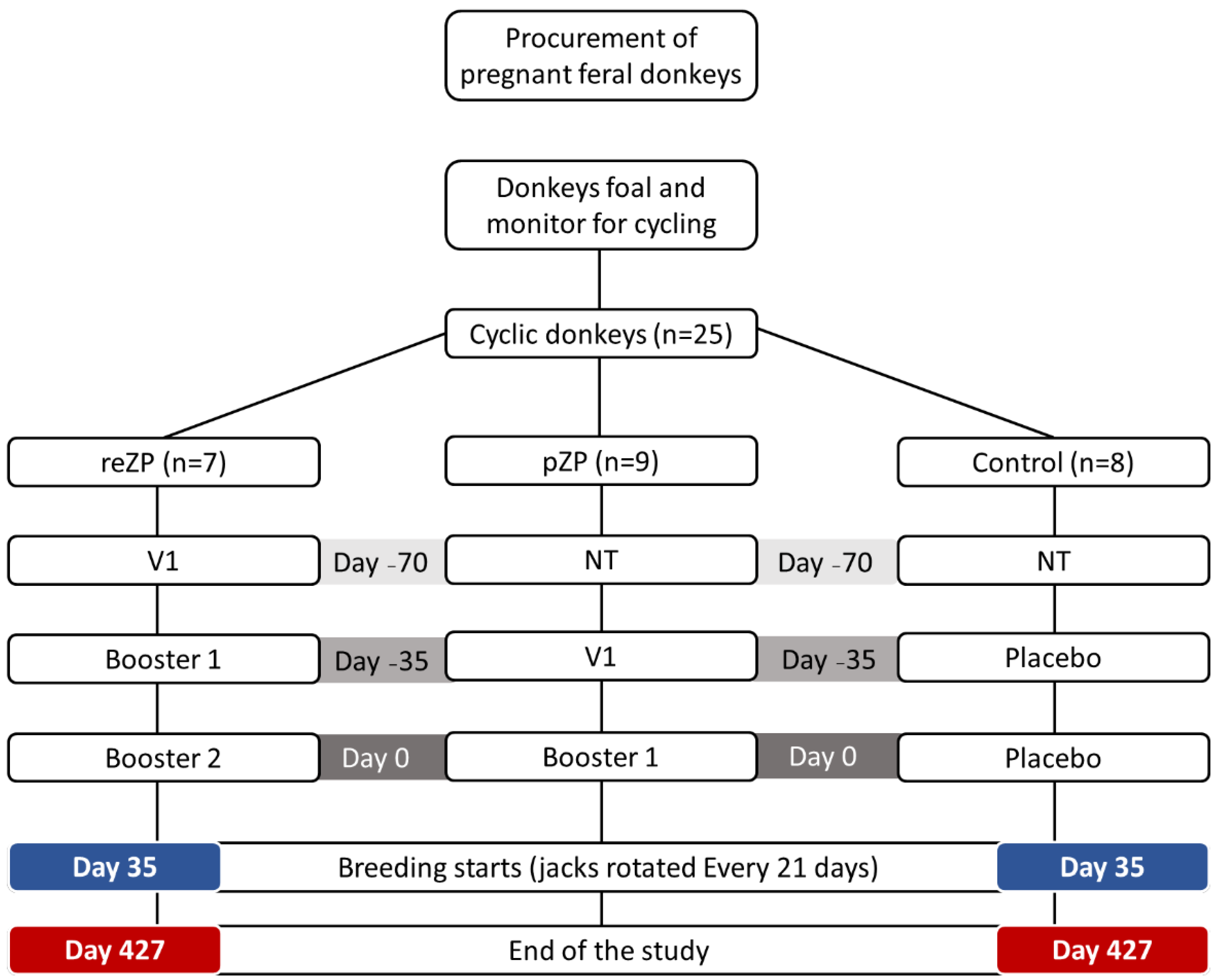
2.2. Study Design
2.3. Treatments
2.3.1. Preparation of Vaccines
2.3.2. Vaccinations
2.4. Observations and Sample Collection
2.4.1. Transrectal Monitoring of the Reproductive Tract
2.4.2. Monitoring of Injection Sites
2.4.3. Collection of Serum Samples
2.4.4. Anti-pZP Antibody Titre Assays
2.4.5. Serum Progesterone Assays
2.5. Data Analysis
3. Results
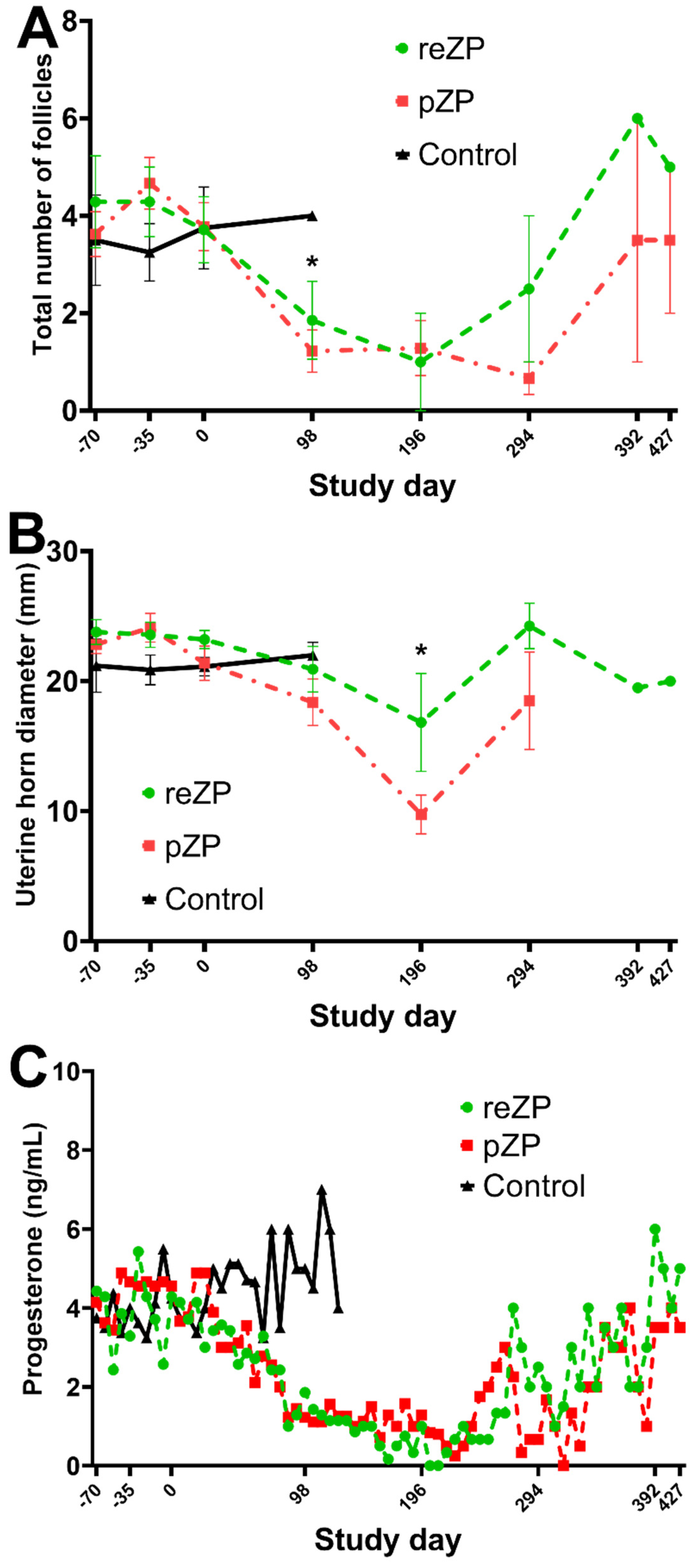
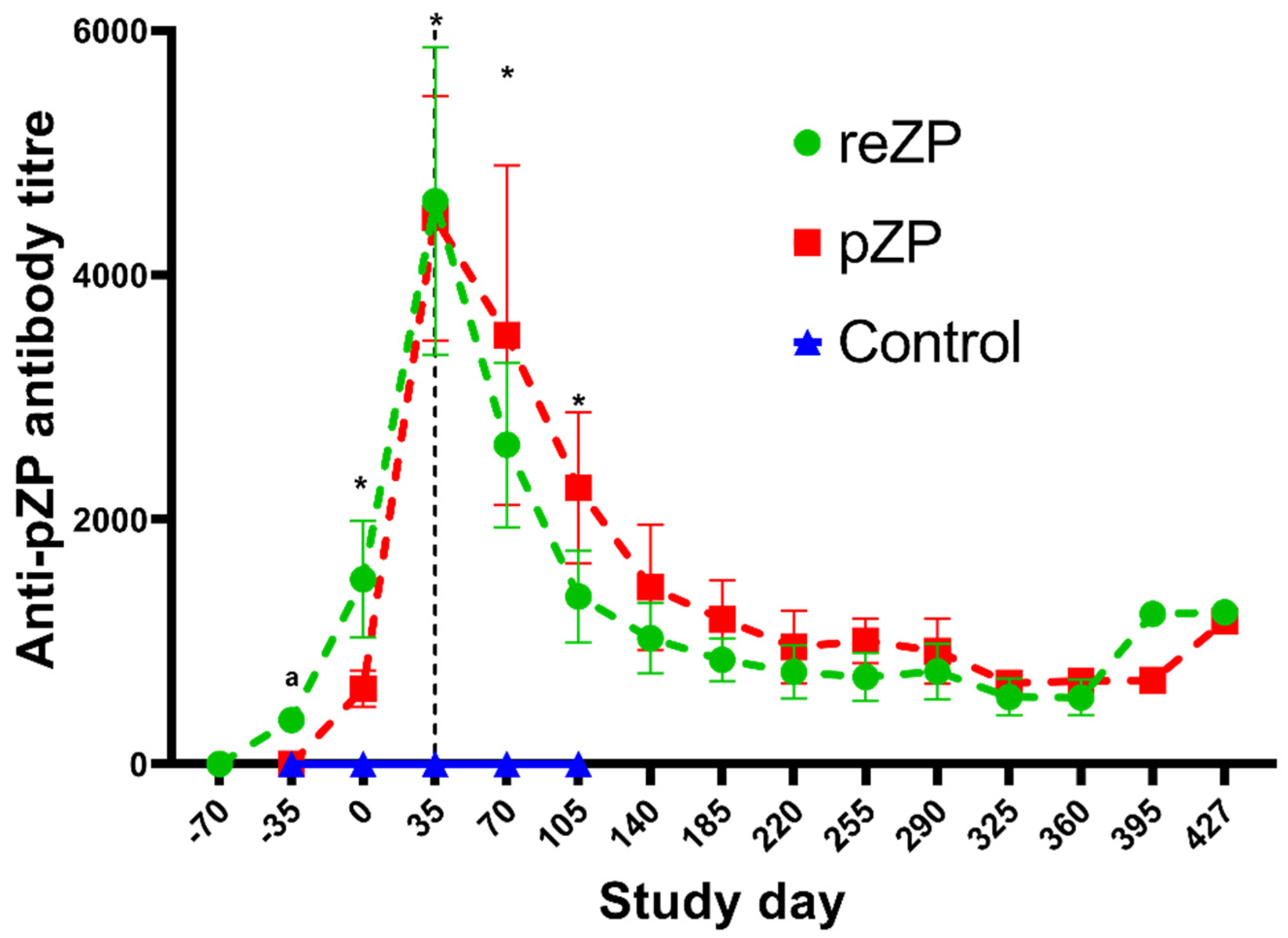
3.1. Effects of Treatments on Uterine Features, Ovarian Function, and Pregnancy Rates
3.2. Injection Site Reactions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feral Donkeys. How the Move from Animal Power to Mechanisation Turns Working Donkeys into Abandoned Donkeys. 2021. Available online: https://www.thedonkeysanctuary.org.uk/about-us/our-international-work/issues/feral-donkeys#:~:text=How%20the%20move%20from%20animal,come%20into%20conflict%20with%20humans (accessed on 17 September 2022).
- Muller, H.; Bourne, A. Minimum population size and potential impact of feral and semi-feral donkeys and horses in an arid rangeland. Afr. Zool. 2018, 53, 139–144. [Google Scholar] [CrossRef]
- Bertschinger, H.J.; Delsink, A.; Van Altena, J.J.; Kirkpatrick, J.F. Porcine zona pellucida vaccine immunocontraception of African elephant (Loxodonta africana) cows: A review of 22 years of research. Bothalia 2018, 48, a2324. [Google Scholar] [CrossRef]
- Gionfriddo, J.P.; Denicola, A.J.; Miller, L.A.; Fagerstone, K.A. Efficacy of GnRH immunocontraception of wild white-tailed deer in New Jersey. Wildl. Soc. Bull. 2011, 35, 142–148. [Google Scholar] [CrossRef]
- Bertschinger, H.J.; Caldwell, P. Fertility suppression of some wildlife species in southern Africa—A review. Reprod. Domest. Anim. 2016, 51, 18–24. [Google Scholar] [CrossRef]
- Garside, D.; Gebril, A.; Alsaadi, M.; Ferro, V.A. Fertility control in wildlife: Review of current status, including novel and future technologies. In Reproductive Sciences in Animal Conservation; Springer: New York, NY, USA, 2014; pp. 467–488. [Google Scholar]
- Riley, S. Model code of practice for the humane control of feral donkeys. J. Int. Wildl. Law Policy 2015, 18, 276–288. [Google Scholar] [CrossRef]
- Hayward, M.W.; Kerley, G.I.H. Fencing for conservation: Restriction of evolutionary potential or a riposte to threatening processes? Biol. Conserv. 2009, 142, 1–13. [Google Scholar] [CrossRef]
- Van Aarde, R.; Whyte, I.; Pimm, S. Culling and the dynamics of the Kruger National Park African elephant population. Anim. Conserv. 1999, 2, 287–294. [Google Scholar] [CrossRef]
- Nicholson, A.J. Supplement: The Balance of Animal Populations. J. Anim. Ecol. 1933, 2, 131–178. [Google Scholar] [CrossRef]
- Wolff, J.O. Population regulation in mammals: An evolutionary perspective. J. Anim. Ecol. 1997, 66, 1–13. [Google Scholar] [CrossRef]
- Kirkpatrick, J.F.; Lyda, R.O.; Frank, K.M. Contraceptive vaccines for wildlife: A review. Am. J. Reprod. Immunol. 2011, 66, 40–50. [Google Scholar] [CrossRef]
- Miller, L.A.; Johns, B.E.; Killian, G.J. Immunocontraception of white-tailed deer using native and recombinant zona pellucida vaccines. Anim. Reprod. Sci. 2000, 63, 187–195. [Google Scholar] [CrossRef][Green Version]
- Schulman, M.L.; Botha, A.E.; Muenscher, S.B.; Annandale, C.H.; Guthrie, A.J.; Bertschinger, H.J. Reversibility of the effects of GnRH-vaccination used to suppress reproduction function in mares. Equine Vet. J. 2013, 45, 111–113. [Google Scholar] [CrossRef] [PubMed]
- Levy, J.K.; Miller, L.A.; Crawford, P.C.; Ritchey, J.W.; Ross, M.K.; Fagerstone, K.A. GnRH immunocontraception of male cats. Theriogenology 2004, 62, 1116–1130. [Google Scholar] [CrossRef] [PubMed]
- Hall, S.E.; Nixon, B.; Aitken, R.J. Non-surgical sterilisation methods may offer a sustainable solution to feral horse (Equus caballus) overpopulation. Reprod. Fertil. Dev. 2017, 29, 1655–1666. [Google Scholar] [CrossRef] [PubMed]
- Killian, G.; Wagner, D.; Miller, L. Observations on the use of the GnRH vaccine GonaconTM in male white-tailed deer (Odocoileus virginianus). In Proceedings of the 11th Wildlife Damage Management Conference, Traverse City, MI, USA, 16–19 May 2005; pp. 256–263. [Google Scholar]
- Lueders, I.; Young, D.; Maree, L.; Van Der Horst, G.; Luther, I.; Botha, S.; Tindall, B.; Fosgate, G.; Ganswindt, A.; Bertschinger, H.J. Effects of GnRH vaccination in wild and captive African Elephant bulls (Loxodonta africana) on reproductive organs and semen quality. PLoS ONE 2017, 12, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Barber, M.R.; Fayrer-Hosken, R.A. Possible mechanisms of mammalian immunocontraception. J. Reprod. Immunol. 2000, 46, 103–124. [Google Scholar] [CrossRef] [PubMed]
- Frank, K.M.; Lyda, R.O.; Kirkpatrick, J.F. Immunocontraception of captive exotic species IV. Species differences in response to the porcine zona pellucida vaccine, timing of booster inoculations, and procedural failures. Zoo Biol. 2005, 24, 349–358. [Google Scholar] [CrossRef]
- French, H.; Peterson, E.; Schulman, M.; Roth, R.; Crampton, M.; Conan, A.; Marchi, S.; Knobel, D.; Bertschinger, H. Efficacy and safety of native and recombinant zona pellucida immunocontraceptive vaccines in donkeys. Theriogenology 2020, 153, 27–33. [Google Scholar] [CrossRef]
- Joonè, C.J.; Bertschinger, H.J.; Gupta, S.K.; Fosgate, G.T.; Arukha, A.P.; Minhas, V.; Dieterman, E.; Schulman, M.L. Ovarian function and pregnancy outcome in pony mares following immunocontraception with native and recombinant porcine zona pellucida vaccines. Equine Vet. J. 2017, 49, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, M.L.; Shellam, G.R.; Papadimitriou, J.M.; Lawson, M.A. Immunocontraception is induced in BALB/c mice inoculated with murine cytomegalovirus expressing mouse zona pellucida 3. Biol. Reprod. 2003, 68, 2024–2032. [Google Scholar] [CrossRef] [PubMed]
- Skinner, S.M.; Kirchick, T.M.H.J.; Dunbar, B.S. Immunization with zona pellucida proteins results in abnormal ovarian follicular differentiation and inhibition of gonadotropin-induce steroid secretion. Endocrinology 1984, 115, 2418–2432. [Google Scholar] [CrossRef] [PubMed]
- Mahi-Brown, C.A.; Yanagimachi, R.; Hoffman, J.C.; Huang, T.T.F. Fertility control in the bitch by active immunization with porcine zonae pellucidae: Use of different adjuvants and patterns of estradiol and progesterone levels in estrous cycles. Biol. Reprod. 1985, 32, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Stoops, M.A.; Liu, I.K.M.; Shideler, S.E.; Lasley, B.L.; Fayrer-Hosken, R.A.; Benirschke, K.; Murata, K.; Van Leeuwen, E.M.G.; Anderson, G.B. Effect of porcine zonae pellucidae immunisation on ovarian follicular development and endocrine function in domestic ewes (Ovis aries). Reprod. Fertil. Dev. 2006, 18, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Tatematsu, K.I.; Ikeda, M.; Wakabayashi, Y.; Yamamura, T.; Kikuchi, K.; Noguchi, J.; Sezutsu, H. Silkworm recombinant bovine zona pellucida protein 4 (bZP4) as a potential female immunocontraceptive antigen; impaired sperm-oocyte interaction and ovarian dysfunction. J. Reprod. Dev. 2021, 67, 402–406. [Google Scholar] [CrossRef] [PubMed]
- Dunbar, B.S.; Lo, C.; Stevens, V. Effect of immunization with purified porcine zona pellucida proteins on ovarian function in baboons. Fertil. Steril. 1989, 52, 311–318. [Google Scholar] [CrossRef]
- Liu, I.K.M.; Bernoco, M.; Feldman, M. Contraception in mares heteroimmunized with pig zonae pellucidae. J. Reprod. Fertil. 1989, 85, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Kirkpatrick, J.F.; Liu, I.K.M.; Turner, J.W. Remotely-delivered immunocontraception in feral horses. Wildl. Soc. Bull. 1990, 18, 326–330. [Google Scholar]
- Kirkpatrick, J.F.; Naugle, R.; Liu, I.K.M.; Bernoco, M.; Turner, J.W. Effects of seven consecutive years of porcine zona pellucida contraception on ovarian function in feral mares. Biol. Reprod. 1995, 52, 411–418. [Google Scholar] [CrossRef]
- Kirkpatrick, J.; Liu, I.; Turner, J.; Naugle, R.; Keiper, R. Long-term effects of porcine zonae pellucidae immunocontraception on ovarian function in feral horses (Equus caballus). J. Reprod. Fertil. 1992, 94, 437–444. [Google Scholar] [CrossRef][Green Version]
- Cain, J.W., III; Marshal, J.P. A porcine zona pellucida immunocontraception of African elephants (Loxodonta africana): Beyond the experimental stage. In Proceedings of the IVth International Wildlife Management Congress, Durban, South Africa, 9–12 July 2012; pp. 95–102. [Google Scholar]
- Fayrer-Hosken, R.A.; Grobler, D.; Van Altena, J.J.; Bertschinger, H.J.; Kirkpatrick, J.F. Immunocontraception of African elephants. Nature 2000, 407, 149. [Google Scholar] [CrossRef]
- Ahlers, M.J.; Ganswindt, A.; Münscher, S.; Bertschinger, H.J. Fecal 20-oxo-pregnane concentrations in free-ranging African elephants (Loxodonta africana) treated with porcine zona pellucida vaccine. Theriogenology 2012, 78, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Nolan, M.B.; Bertschinger, H.J.; Roth, R.; Crampton, M.; Martins, I.S.; Fosgate, G.T.; Stout, T.A.; Schulman, M.L. Ovarian function following immunocontraceptive vaccination of mares using native porcine and recombinant zona pellucida vaccines formulated with a non-Freund’s adjuvant and anti-GnRH vaccines. Theriogenology 2018, 120, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Joonè, C.J.; Schulman, M.L.; Bertschinger, H.J. Ovarian dysfunction associated with zona pellucida–based immunocontraceptive vaccines. Theriogenology 2017, 89, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Billiau, A.; Matthys, P. Modes of action of Freund’s adjuvants in experimental models of autoimmune diseases. J. Leukoc. Biol. 2001, 70, 849–860. [Google Scholar] [CrossRef]
- Ovarian, A.Z.; Thillaikoothan, P. Role of adjuvants in inhibitory influence of immunization in Bonnet Monkeys: A morphological study. Biol. Reprod. 1989, 41, 665–673. [Google Scholar]
- Nolan, M.B.; Schulman, M.L.; Botha, A.E.; Human, A.M.; Roth, R.; Crampton, M.C.; Bertschinger, H.J. Serum antibody immunoreactivity and safety of native porcine and recombinant zona pellucida vaccines formulated with a non-Freund’s adjuvant in horses. Vaccine 2019, 37, 1299–1306. [Google Scholar] [CrossRef]
- Roelle, J.E.; Ransom, J.I. Injection-Site Reactions in Wild Horses (Equus caballus) Receiving an Immunocontraceptive Vaccine; US Geological Survey: Reston, VA, USA, 2009; pp. 1–15.
- Turner, J.W.; Liu, I.K.M.; Kirkpatrick, J.F. Remotely delivered immunocontraception in free-roaming feral burros (Equus asinus). J. Reprod. Fertil. 1996, 107, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Cauchard, S.; Bertrand, F.; Barrier-Battut, I.; Jacquet, S.; Laurentie, M.; Barbey, C.; Laugier, C.; Deville, S.; Cauchard, J. Assessment of the safety and immunogenicity of Rhodococcus equi-secreted proteins combined with either a liquid nanoparticle (IMS 3012) or a polymeric (PET GEL A) water-based adjuvant in adult horses and foals-Identification of promising new candidate ant. Vet. Immunol. Immunopathol. 2014, 157, 164–174. [Google Scholar] [CrossRef]
- Stahl-Hennig, C.; Eisenblätter, M.; Jasny, E.; Rzehak, T.; Tenner-Racz, K.; Trumpfheller, C.; Salazar, A.M.; Überla, K.; Nieto, K.; Kleinschmidt, J.; et al. Synthetic double-stranded RNAs are adjuvants for the induction of t helper 1 and humoral immune responses to human papillomavirus in rhesus macaques. PLoS Pathog. 2009, 5, e1000373. [Google Scholar] [CrossRef] [PubMed]
- Pulko, V.; Liu, X.; Krco, C.J.; Harris, K.J.; Frigola, X.; Kwon, E.D.; Dong, H. TLR3-stimulated dendritic cells up-regulate B7-H1 expression and influence the magnitude of CD8 T cell responses to tumor vaccination. J. Immunol. 2009, 183, 3634–3641. [Google Scholar] [CrossRef] [PubMed]
- Apostólico, J.D.S.; Alves, V.; Lunardelli, S.; Coirada, F.C.; Boscardin, S.B.; Rosa, D.S. Adjuvants: Classification, modus operandi, and licensing. J. Immunol. Res. 2016, 2016, 1459394. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Nguyen, M.T. Recent advances of vaccine adjuvants for infectious diseases. Immune Netw. 2015, 15, 51–57. [Google Scholar] [CrossRef]
- Oliveira, S.N.; Canuto, L.; Segabinazzi, L.G.T.M.; Dell’Aqua Junior, J.A.; Papa, P.; Fonseca, M.; de Lisboa Ribeiro Filho, A.; Papa, F. Histrelin acetate-induced ovulation in Brazilian Northeastern jennies (Equus asinus) with different follicle diameters. Theriogenology 2019, 136, 95–100. [Google Scholar] [CrossRef] [PubMed]
- du Toit, N.; Burden, F.A.; Dixon, P.M. Clinical dental examinations of 357 donkeys in the UK. Part 1: Prevalence of dental disorders. Equine Vet. J. 2009, 41, 390–394. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Shrestha, A.; Panda, A.K.; Gupta, S.K. Production of tag-free recombinant fusion protein encompassing promiscuous T cell epitope of tetanus toxoid and dog zona pellucida glycoprotein-3 for contraceptive vaccine development. Mol. Biotechnol. 2013, 54, 853–862. [Google Scholar] [CrossRef]
- Kollipara, S.; Bende, G.; Agarwal, N.; Varshney, B.; Paliwal, J. International guidelines for bioanalytical method validation: A comparison and discussion on current scenario. Chromatographia 2011, 73, 201–217. [Google Scholar] [CrossRef]
- Bechert, U.; Bartell, J.; Kutzler, M.; Menino, A.; Bildfell, R.; Anderson, M.; Fraker, M. Effects of two porcine zona pellucida immunocontraceptive vaccines on ovarian activity in horses. J. Wildl. Manag. 2013, 77, 1386–1400. [Google Scholar] [CrossRef]
- Peterson, E.W.; Segabinazzi, L.G.T.M.; Gilbert, R.O.; Bergfelt, D.R.; French, H.M. Evaluation of stress accompanying immunocontraceptive vaccination in donkeys. Animals 2022, 12, 457. [Google Scholar] [CrossRef] [PubMed]
- Ransom, J.I.; Hobbs, N.T.; Bruemmer, J. Contraception can lead to trophic asynchrony between birth pulse and resources. PLoS ONE 2013, 8, e54972. [Google Scholar] [CrossRef] [PubMed]
- Powell, D.M.; Monfort, S.L. Assessment: Effects of porcine zona pellucida immunocontraception on estrous cyclicity in feral horses. J. Appl. Anim. Welf. Sci. 2001, 4, 271–284. [Google Scholar] [CrossRef]
- Joonè, C.J.; Schulman, M.L.; Fosgate, G.T.; Claes, A.N.J.; Gupta, S.K.; Botha, A.E.; Human, A.; Bertschinger, H.J. Serum anti-Müllerian hormone dynamics in mares following immunocontraception with anti-zona pellucida or -GnRH vaccines. Theriogenology 2018, 106, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Stefaniuk-Szmukier, M.; Ropka-Molik, K.; Zagrajczuk, A.; Piórkowska, K.; Szmatoła, T.; Łuszczyński, J.; Bugno-Poniewierska, M. Genetic variability in equine GDF9 and BMP15 genes in Arabian and thoroughbred mares. Ann. Anim. Sci. 2018, 18, 39–52. [Google Scholar] [CrossRef]
- Barbaccia, M.L.; Serra, M.; Purdy, R.H.; Biggio, G. Stress and neuroactive steroids. Int. Rev. Neurobiol. 2001, 46, 243–272. [Google Scholar] [CrossRef] [PubMed]
- Genazzani, A.R.; Petraglia, F.; Bernardi, F.; Casarosa, E.; Salvestroni, C.; Tonetti, A.; Nappi, R.E.; Luisi, S.; Palumbo, M.; Purdy, R.H.; et al. Circulating levels of allopregnanolone in humans: Gender, age, and endocrine influences. J. Clin. Endocrinol. Metab. 1998, 83, 2099–2103. [Google Scholar] [CrossRef]
- Purdy, R.H.; Morrow, A.L.; Moore, P.H.; Paul, S.M. Stress-induced elevations of γ-aminobutyric acid type a receptor-active steroids in the rat brain. Proc. Natl. Acad. Sci. USA 1991, 88, 4553–4557. [Google Scholar] [CrossRef] [PubMed]
- Tsuma, V.T.; Einarsson, S.; Madej, A.; Forsberg, M.; Lundeheim, N. Plasma levels of progesterone and cortisol after ACTH administration in lactating primiparous sows. Acta Vet. Scand. 1998, 39, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Van Lier, E.; Anderson, H.; Clariget, R.P.; Forsberg, M. Effects of administration of adrenocorticotrophic hormone (ACTH) on extragonadal progesterone levels in sheep. Reprod. Domest. Anim. 1998, 33, 55–59. [Google Scholar] [CrossRef]
- Bolanos, J.M.; Molina, J.R.; Forsberg, M. Effect of blood sampling and administration of ACTH on cortisol and progesterone levels in ovariectomized Zebu cows (Bos indicus). Acta Vet. Scand. 1997, 38, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Herrera, A.Y.; Nielsen, S.E.; Mather, M. Stress-induced increases in progesterone and cortisol in naturally cycling women. Neurobiol. Stress 2016, 3, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Wirth, M.M.; Gaffey, A.E. Stress, rejection, and hormones: Cortisol and progesterone reactivity to laboratory speech and rejection tasks in women and men. F1000Research 2014, 3, 208. [Google Scholar] [CrossRef]
- Chatdarong, K.; Ponglowhapan, S.; Karlsson, Å.; Linde-Forsberg, C. The effect of ACTH stimulation on cortisol and progesterone concentrations in intact and ovariohysterectomized domestic cats. Theriogenology 2006, 66, 1482–1487. [Google Scholar] [CrossRef] [PubMed]
- Hydbring-Sandberg, E.; Larsson, E.; Madej, A.; Höglund, O.V. Short-term effect of ovariohysterectomy on urine serotonin, cortisol, testosterone and progesterone in bitches. BMC Res. Notes 2021, 14, 265. [Google Scholar] [CrossRef] [PubMed]
- Romeo, R.D.; Lee, S.J.; McEwen, B.S. Differential stress reactivity in intact and ovariectomized prepubertal and adult female rats. Neuroendocrinology 2004, 80, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Araujo, R.R.; Ginther, O.J.; Ferreira, J.C.; Palhão, M.M.; Beg, M.A.; Wiltbank, M.C. Role of follicular estradiol-17beta in timing of luteolysis in heifers. Biol. Reprod. 2009, 81, 426–437. [Google Scholar] [CrossRef]
- Griffin, P.G.; Ginther, O.J. Dynamics of uterine diameter and endometrial morphology during the estrous cycle and early pregnancy in mares. Anim. Reprod. Sci. 1991, 25, 133–142. [Google Scholar] [CrossRef]
- Bergfelt, D.R.; Ginther, O.J. Ovarian, uterine and embryo dynamics in horses versus ponies. J. Equine Vet. Sci. 1996, 16, 66–72. [Google Scholar] [CrossRef]
- McKinnon, A.O.; Squires, E.L.; Pickett, B.W. Equine Reproductive Ultrasonography; Bulletin No.4 for Colorado State University; Animal Reproduction Laboratory: Fort Collins, CO, USA, 1988; pp. 31–40. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
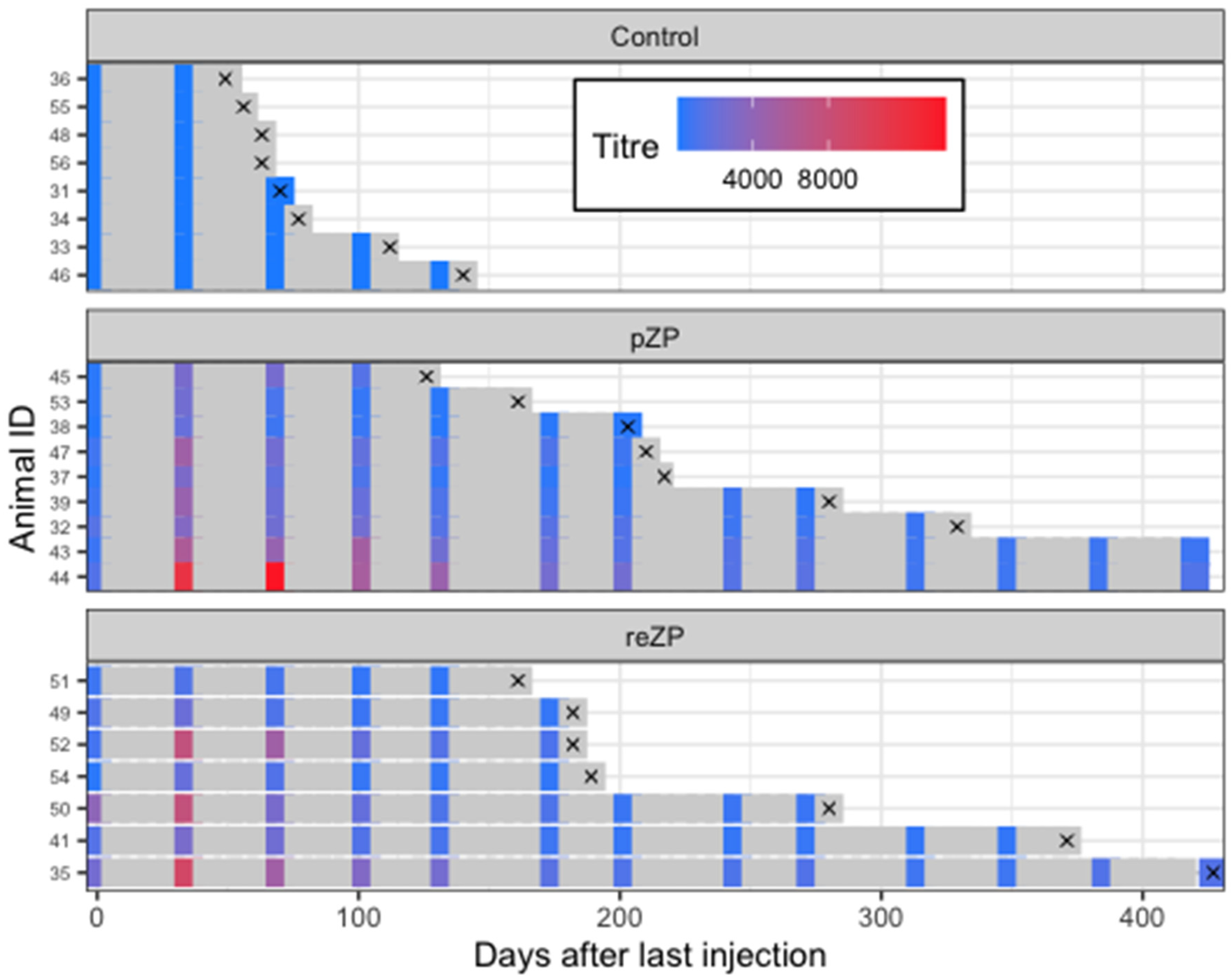{kind=link}
| End Points | reZP (n = 7) | pZP (n = 9) | Overall Vaccinated Jennies | Control (n = 8) |
|---|---|---|---|---|
| Interval from Day 0 to shutdown | 118 ± 33 days (4/7) | 91 ± 20 days (4/9) | 104 ± 29 days | - |
| Duration of ovarian shutdown | 75 ± 59 days | 146 ± 92 days | 114 ± 83 days | - |
| Interval to pregnancy from Day 0 | 244 ± 104 days (7/7) a | 218 ± 69 days (7/9) a | 232 ± 87 days (14/16) | 78 ± 31 days (8/8) b |
| Interval between the end of the ovarian shutdown and pregnancy | 75 ± 84 days (4/7) | 102 ± 89 days (4/9) | 88 ± 82 days (8/16) | - |
| Anti-pZP Antibody | reZP (n = 7) | pZP (n = 9) | Overall Vaccinated Jennies | Control (n = 8) |
|---|---|---|---|---|
| Titres at Day 35 of the study | 4604 ± 3325 a | 4465 ± 3001 a | 4526 ± 3038 a | 0 b |
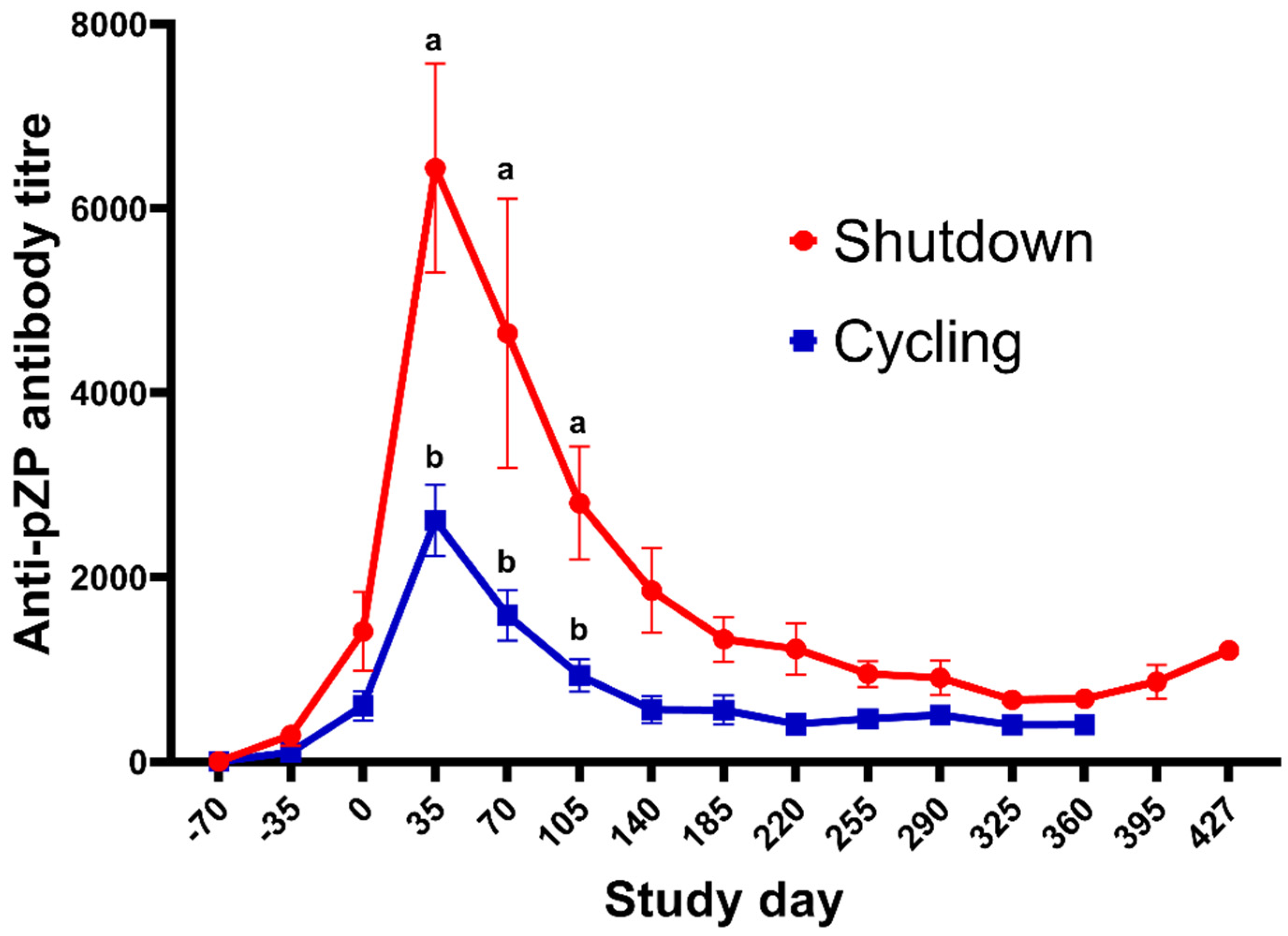
| Titres at Day 35 of jennies with ovarian shutdown | 6550 ± 3144 (4/7) | 6322 ± 3742 (4/9) | 6436 ± 3202 Y | - |
| Titres at Day 35 of jennies with no ovarian shutdown | 2008 ± 806 (3/7) | 2980 ± 511 (5/9) | 2612 ± 1088 X | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
French, H.; Segabinazzi, L.; Middlebrooks, B.; Peterson, E.; Schulman, M.; Roth, R.; Crampton, M.; Conan, A.; Marchi, S.; Gilbert, T.; et al. Efficacy and Safety of Native and Recombinant Zona Pellucida Immunocontraceptive Vaccines Formulated with Non-Freund’s Adjuvants in Donkeys. Vaccines 2022, 10, 1999. https://doi.org/10.3390/vaccines10121999
French H, Segabinazzi L, Middlebrooks B, Peterson E, Schulman M, Roth R, Crampton M, Conan A, Marchi S, Gilbert T, et al. Efficacy and Safety of Native and Recombinant Zona Pellucida Immunocontraceptive Vaccines Formulated with Non-Freund’s Adjuvants in Donkeys. Vaccines. 2022; 10(12):1999. https://doi.org/10.3390/vaccines10121999
Chicago/Turabian StyleFrench, Hilari, Lorenzo Segabinazzi, Brittany Middlebrooks, Erik Peterson, Martin Schulman, Robyn Roth, Michael Crampton, Anne Conan, Silvia Marchi, Trevor Gilbert, and et al. 2022. "Efficacy and Safety of Native and Recombinant Zona Pellucida Immunocontraceptive Vaccines Formulated with Non-Freund’s Adjuvants in Donkeys" Vaccines 10, no. 12: 1999. https://doi.org/10.3390/vaccines10121999
APA StyleFrench, H., Segabinazzi, L., Middlebrooks, B., Peterson, E., Schulman, M., Roth, R., Crampton, M., Conan, A., Marchi, S., Gilbert, T., Knobel, D., & Bertschinger, H. (2022). Efficacy and Safety of Native and Recombinant Zona Pellucida Immunocontraceptive Vaccines Formulated with Non-Freund’s Adjuvants in Donkeys. Vaccines, 10(12), 1999. https://doi.org/10.3390/vaccines10121999