
Subcutaneous Immunization with Unaltered Axenic Malaria Parasite Liver Stages Induces Sterile Protection against Infectious Sporozoite Challenge

Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Parasite
2.2. Salivary Gland Sporozoite Collection and In-Vitro Transformation
2.3. Live Fluorescence Imaging
2.4. Infectivity of Sporozoites and Axenic LS in mice
2.5. Vaccination
2.6. Parasite Challenge
3. Results
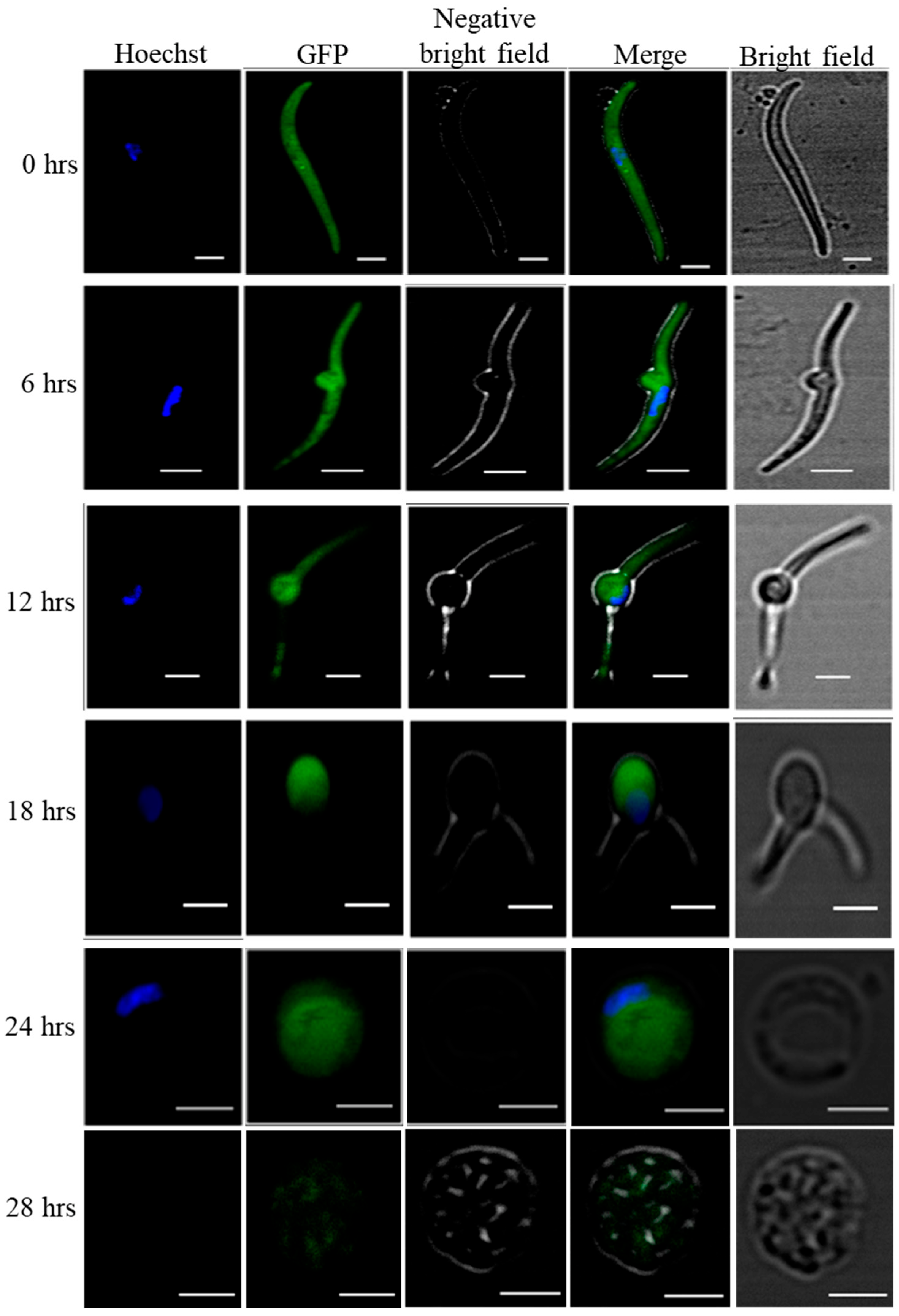
3.1. In Vitro Cultured Sporozoites Transform into Early Liver Stages within 24 hrs of Incubation
3.2. Immunization with Cell-Free Developed Early Liver Stages is Safe and Provides Sterile Protection
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaslow, D.C. Malaria vaccine research & innovation: The intersection of IA2030 and zero malaria. NPJ Vaccines 2020, 5, 109. [Google Scholar] [CrossRef] [PubMed]
- WHO. World Malaria Report 2020. Available online: https://www.who.int/publications/i/item/9789240015791 (accessed on 6 August 2022).
- Blasco, B.; Leroy, D.; Fidock, D.A. Antimalarial drug resistance: Linking Plasmodium falciparum parasite biology to the clinic. Nat. Med. 2017, 23, 917–928. [Google Scholar] [CrossRef] [PubMed]
- Duffy, P.E.; Patrick Gorres, J. Malaria vaccines since 2000: Progress, priorities, products. NPJ Vaccines 2020, 5, 48. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Pan, H.; Gu, Y.; Zuo, X.; Ran, N.; Yuan, Y.; Zhang, C.; Wang, F. Prospects for malaria vaccines: Pre-erythrocytic stages, blood stages, and transmission-blocking stages. BioMed Res. Int. 2019, 2019, 9751471. [Google Scholar] [CrossRef] [PubMed]
- Aly, A.S.I.; Vaughan, A.M.; Kappe, S.H.I. Malaria Parasite development in the mosquito and infection of the mammalian host. Annu. Rev. Microbiol. 2009, 63, 195–221. [Google Scholar] [CrossRef]
- Vaughan, A.M.; Aly, A.S.; Kappe, S.H. Malaria Parasite Pre-Erythrocytic Stage Infection: Gliding and Hiding. Available online: ttp://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Citation&list_uids=18779047 (accessed on 6 August 2022).
- Mikolajczak, S.A.; Aly, A.S.; Kappe, S.H. Preerythrocytic malaria vaccine development. Curr. Opin. Infect. Dis. 2007, 20, 461–466. [Google Scholar] [CrossRef]
- Reeder, S.M.; Reuschel, E.L.; Bah, M.A.; Yun, K.; Tursi, N.J.; Kim, K.Y.; Chu, J.; Zaidi, F.I.; Yilmaz, I.; Hart, R.J.; et al. Synthetic DNA vaccines adjuvanted with pIL-33 drive liver-localized t cells and provide protection from plasmodium challenge in a mouse model. Vaccines 2020, 8, 21. [Google Scholar] [CrossRef]
- Rider, P.J.F.; Kamil, M.; Yilmaz, I.; Atmaca, H.N.; Kalkan-Yazici, M.; Ziya Doymaz, M.; Kousoulas, K.G.; Aly, A.S.I. An attenuated HSV-1-derived malaria vaccine expressing liver-stage exported proteins induces sterilizing protection against infectious sporozoite challenge. Vaccines 2022, 10, 300. [Google Scholar] [CrossRef]
- Beeson, J.G.; Kurtovic, L.; Dobaño, C.; Opi, D.H.; Chan, J.A.; Feng, G.; Good, M.F.; Reiling, L.; Boyle, M.J. Challenges and strategies for developing efficacious and long-lasting malaria vaccines. Sci. Transl. Med. 2019, 11. [Google Scholar] [CrossRef]
- Duffy, P.E.; Sahu, T.; Akue, A.; Milman, N.; Anderson, C. Pre-erythrocytic malaria vaccines: Identifying the targets. Expert Rev. Vaccines 2012, 11, 1261–1280. [Google Scholar] [CrossRef]
- Molina-Franky, J.; Cuy-Chaparro, L.; Camargo, A.; Reyes, C.; Gómez, M.; Salamanca, D.R.; Patarroyo, M.A.; Patarroyo, M.E. Plasmodium falciparum pre-erythrocytic stage vaccine development. Malar. J. 2020, 19, 56. [Google Scholar] [CrossRef]
- Kaiser, K.; Camargo, N.; Kappe, S.H.I. Transformation of sporozoites into early exoerythrocytic malaria parasites does not require host cells. J. Exp. Med. 2003, 197, 1045–1050. [Google Scholar] [CrossRef]
- Hart, R.J.; Lawres, L.; Fritzen, E.; Mamoun, C.B.; Aly, A.S.I. Plasmodium yoelii vitamin B5 pantothenate transporter candidate is essential for parasite transmission to the mosquito. Sci. Rep. 2014, 4, 5665. [Google Scholar] [CrossRef]
- Aly, A.S.I.; Deveci, G.; Yilmaz, I.; Abraham, A.; Golshan, A.; Hart, R.J. Phenotypic analysis of rodent malaria parasite asexual and sexual blood stages and mosquito stages. J. Vis. Exp. 2019, 147, e55688. [Google Scholar] [CrossRef]
- Silvie, O.; Semblat, J.P.; Franetich, J.F.; Hannoun, L.; Eling, W.; Mazier, D. Effects of irradiation on Plasmodium falciparum sporozoite hepatic development: Implications for the design of pre-erythrocytic malaria vaccines. Parasite Immunol. 2002, 24, 221–223. [Google Scholar] [CrossRef]
- Itsara, L.S.; Zhou, Y.; Do, J.; Grieser, A.M.; Vaughan, A.M.; Ghosh, A.K. The Development of whole sporozoite vaccines for Plasmodium falciparum malaria. Front. Immunol. 2018, 9, 2748. [Google Scholar] [CrossRef]
- Hoffman, S.L.; Goh, L.M.L.; Luke, T.C.; Schneider, I.; Le, T.P.; Doolan, D.L.; Sacci, J.; de la Vega, P.; Dowler, M.; Paul, C.; et al. Protection of humans against malaria by immunization with radiation-attenuated Plasmodium falciparum sporozoites. J. Infect. Dis. 2002, 185, 1155–1164. [Google Scholar] [CrossRef]
- Kublin, J.G.; Mikolajczak, S.A.; Sack, B.K.; Fishbaugher, M.E.; Seilie, A.; Shelton, L.; VonGoedert, T.; Firat, M.; Magee, S.; Fritzen, E.; et al. Complete attenuation of genetically engineered Plasmodium falciparum sporozoites in human subjects. Sci. Transl. Med. 2017, 9. [Google Scholar] [CrossRef]
- Mordmuller, B.; Surat, G.; Lagler, H.; Chakravarty, S.; Ishizuka, A.S.; Lalremruata, A.; Gmeiner, M.; Campo, J.J.; Esen, M.; Ruben, A.J.; et al. Sterile protection against human malaria by chemoattenuated PfSPZ vaccine. Nature 2017, 542, 445–449. [Google Scholar] [CrossRef]
- Mendes, A.M.; Machado, M.; Gonçalves-Rosa, N.; Reuling, I.J.; Foquet, L.; Marques, C.; Salman, A.M.; Yang, A.S.P.; Moser, K.A.; Dwivedi, A.; et al. A Plasmodium berghei sporozoite-based vaccination platform against human malaria. NPJ Vaccines 2018, 3, 33. [Google Scholar] [CrossRef]
- Kreutzfeld, O.; Muller, K.; Matuschewski, K. Engineering of genetically arrested parasites (GAPs) for a precision malaria vaccine. Front. Cell. Infect. Microbiol. 2017, 7, 198. [Google Scholar] [CrossRef] [PubMed]
- Schofield, L.; Villaquiran, J.; Ferreira, A.; Schellekens, H.; Nussenzweig, R.; Nussenzweig, V. γ Interferon, CD8+ T cells and antibodies required for immunity to malaria sporozoites. Nature 1987, 330, 664–666. [Google Scholar] [CrossRef] [PubMed]
- Seder, R.A.; Chang, L.J.; Enama, M.E.; Zephir, K.L.; Sarwar, U.N.; Gordon, I.J.; Holman, L.A.; James, E.R.; Billingsley, P.F.; Gunasekera, A.; et al. Protection against malaria by intravenous immunization with a nonreplicating sporozoite vaccine. Science 2013, 341, 1359–1365. [Google Scholar] [CrossRef] [PubMed]
- Lyke, K.E.; Ishizuka, A.S.; Berry, A.A.; Chakravarty, S.; DeZure, A.; Enama, M.E.; James, E.R.; Billingsley, P.F.; Gunasekera, A.; Manoj, A.; et al. Attenuated PfSPZ vaccine induces strain-transcending T cells and durable protection against heterologous controlled human malaria infection. Proc. Natl. Acad. Sci. USA 2017, 114, 2711–2716. [Google Scholar] [CrossRef]


{kind=link}
| Infectivity of Axenic LS vs. Salivary Gland Sporozoites in BALB/c Mice | Sterile Protection against Malaria Parasite Sporozoite Challenge Following Subcutaneous Immunization with Axenic LS | |||||||
| Group (n) | Infection Dose | Route | Infected/ Injected | Group (n) | Immunization Dose | Immunization Intervals | Route | Protected/ Challenged |
| Group A: Pyp230p(−) Axenic LS (3) | 500,000 | IV | 0/3 | Group 1:Pyp230p(−) Axenic LS (10) | 200,000 | 0, 21, 42, 63 | SC | 10/10 |
| Group B: Pyp230p(−) Axenic LS (3) | 1,000,000 | IV | 0/3 | Group 2: PyWT Axenic LS (10) | 200,000 | 0, 21, 42, 63 | SC | 10/10 |
| Group C: Pyp230p(−) Sporozoites (3) | 500 | IV | 3/3 | Group 3: UninfectedAnopheles stephensisalivary glands (5) | N/A | 0, 21, 42, 63 | SC | 0/5 |
| Group D: PyWT Axenic LS (3) | 500,000 | IV | 0/3 | Group 4: Naive Control (5) | N/A | N/A | N/A | 0/5 |
| Group E: PyWT Axenic LS (3) | 1,000,000 | IV | 0/3 | N/A | N/A | N/A | N/A | N/A |
| Group F: PyWT Sporozoites (3) | 500 | IV | 3/3 | N/A | N/A | N/A | N/A | N/A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamil, M.; Deveci, G.; Kina, U.Y.; Kappe, S.H.I.; Aly, A.S.I. Subcutaneous Immunization with Unaltered Axenic Malaria Parasite Liver Stages Induces Sterile Protection against Infectious Sporozoite Challenge. Vaccines 2022, 10, 1884. https://doi.org/10.3390/vaccines10111884
Kamil M, Deveci G, Kina UY, Kappe SHI, Aly ASI. Subcutaneous Immunization with Unaltered Axenic Malaria Parasite Liver Stages Induces Sterile Protection against Infectious Sporozoite Challenge. Vaccines. 2022; 10(11):1884. https://doi.org/10.3390/vaccines10111884
Chicago/Turabian StyleKamil, Mohd, Gozde Deveci, Umit Y. Kina, Stefan H. I. Kappe, and Ahmed S. I. Aly. 2022. "Subcutaneous Immunization with Unaltered Axenic Malaria Parasite Liver Stages Induces Sterile Protection against Infectious Sporozoite Challenge" Vaccines 10, no. 11: 1884. https://doi.org/10.3390/vaccines10111884
APA StyleKamil, M., Deveci, G., Kina, U. Y., Kappe, S. H. I., & Aly, A. S. I. (2022). Subcutaneous Immunization with Unaltered Axenic Malaria Parasite Liver Stages Induces Sterile Protection against Infectious Sporozoite Challenge. Vaccines, 10(11), 1884. https://doi.org/10.3390/vaccines10111884