
Eimeria maxima Rhomboid-like Protein 5 Provided Partial Protection against Homologous Challenge in Forms of Recombinant Protein and DNA Plasmid in Chickens




Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmids, Parasites and Animals
2.2. Cloning of EmROM5 Gene
2.3. Construction of Recombinant Plasmids pET-32a-ntmEmROM5 and pVAX-EmROM5
2.4. Preparation of NtmEmROM5 Recombinant Protein (rEmROM5), Chicken Anti-E. maxima Serum and Rat Anti-rEmROM5 Serum
2.5. Western Blot Analysis of rEmROM5
2.6. Reverse Transcription PCR and Western Blot Analysis of Transcription and Expression of pVAX-EmROM5 In Vivo
2.7. Determination of Immune Response Induced by EmROM5 in Chickens
2.7.1. Animal Immunization
2.7.2. Determination of EmROM5-Induced Changes in Spleen T Lymphocyte Subpopulations by Flow Cytometry
2.7.3. Determination of EmROM5-Induced Changes in Cytokines by Quantitative Real-Time PCR
2.7.4. Determination of EmROM5-Specific IgG Antibody Level by Indirect ELISA
2.8. Assessment of Protective Efficacy of EmROM5 against Challenge with E. maxima
2.9. Statistical Analysis
3. Results
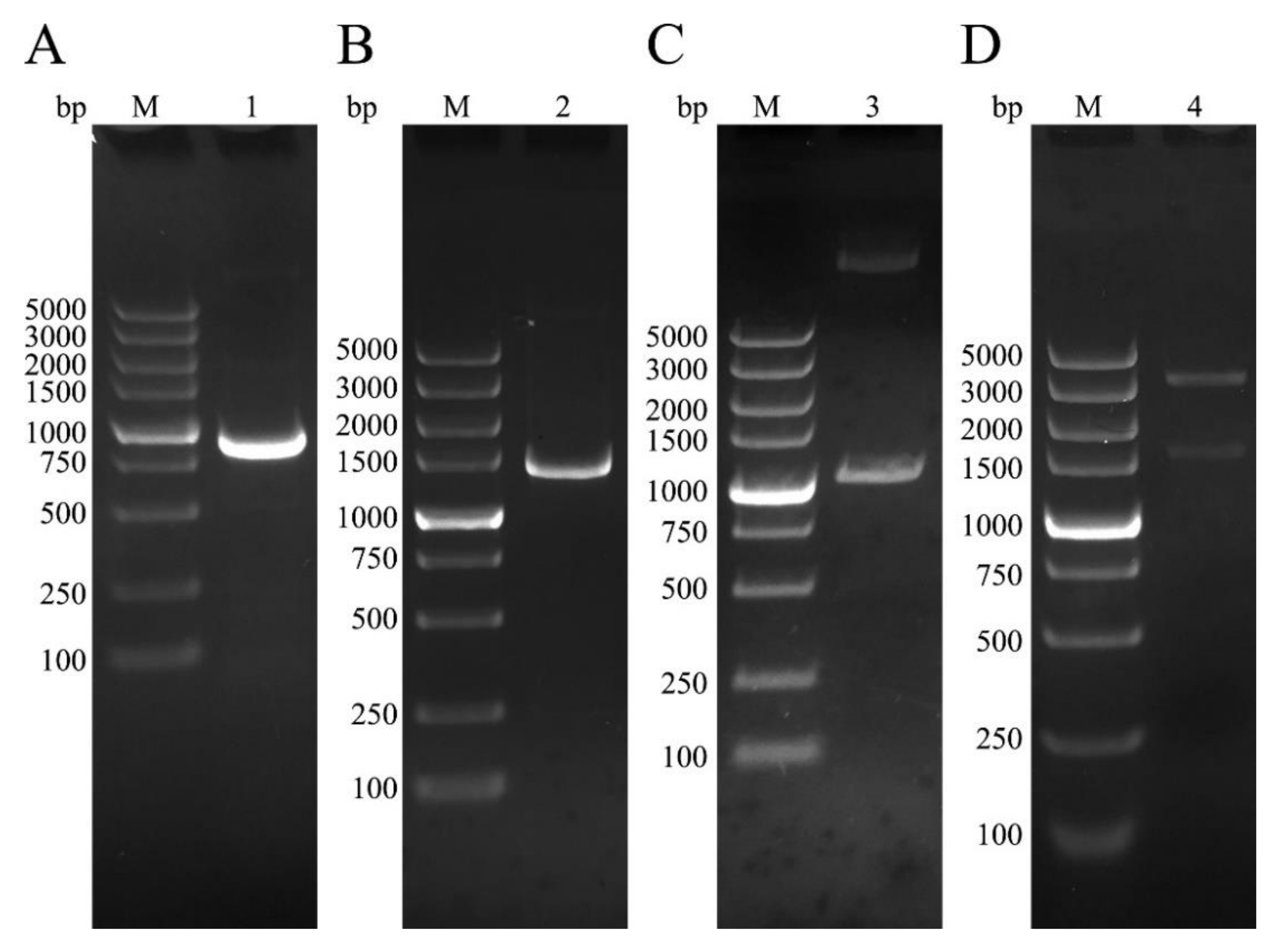
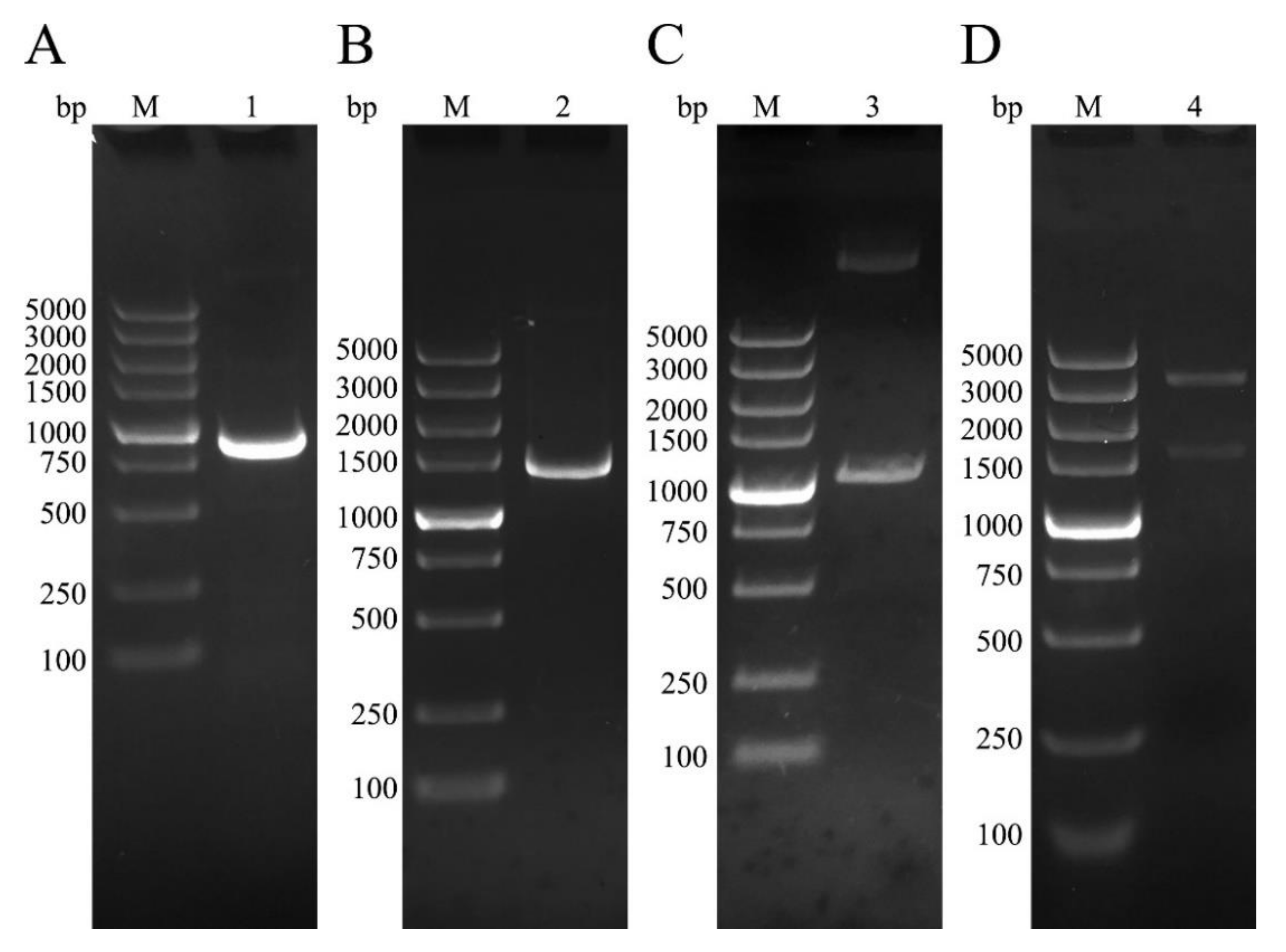
3.1. Cloning of NtmEmROM5 and EmROM5, Construction of pET-32a-ntmEmROM5 and pVAX-EmROM5
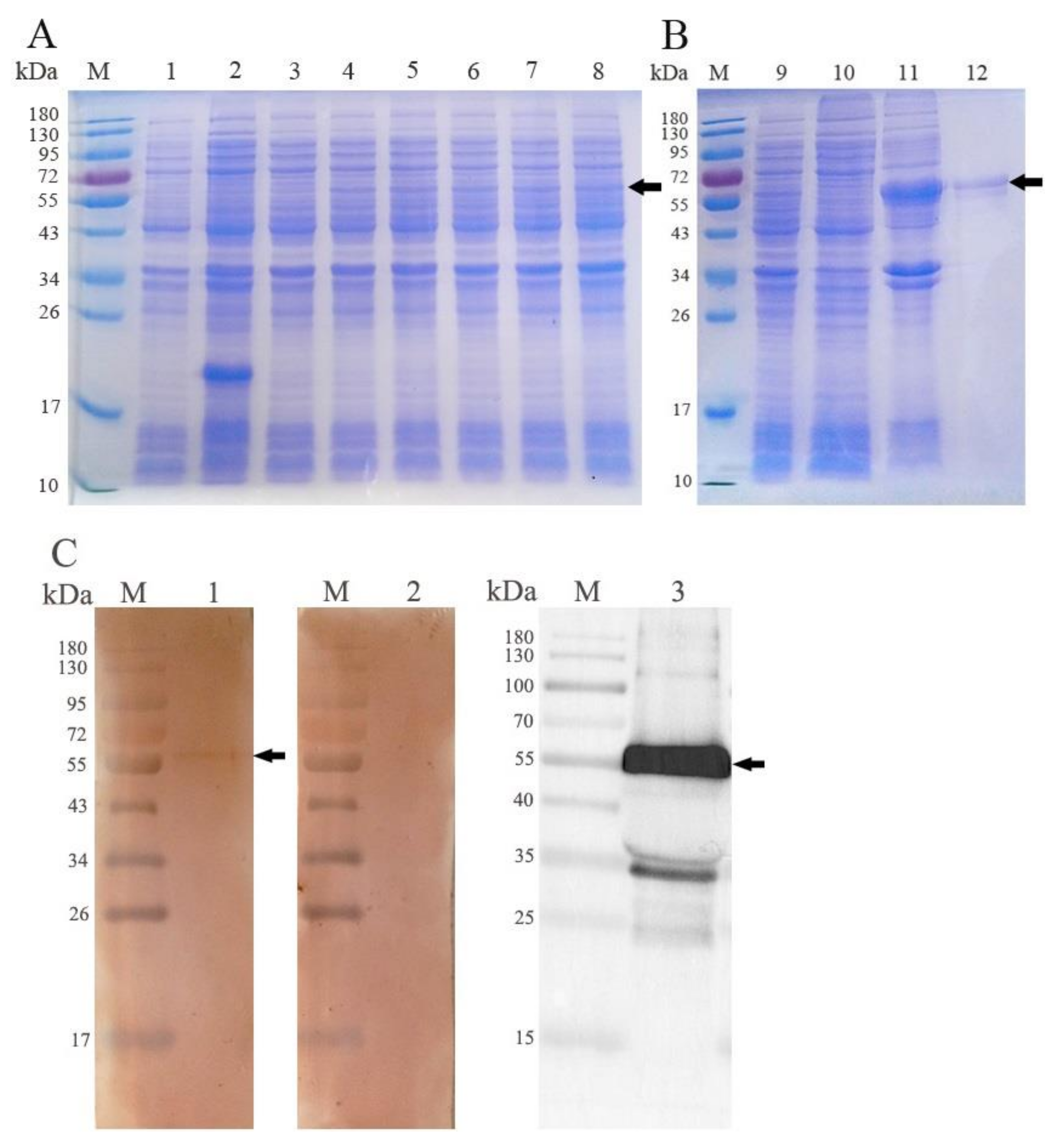
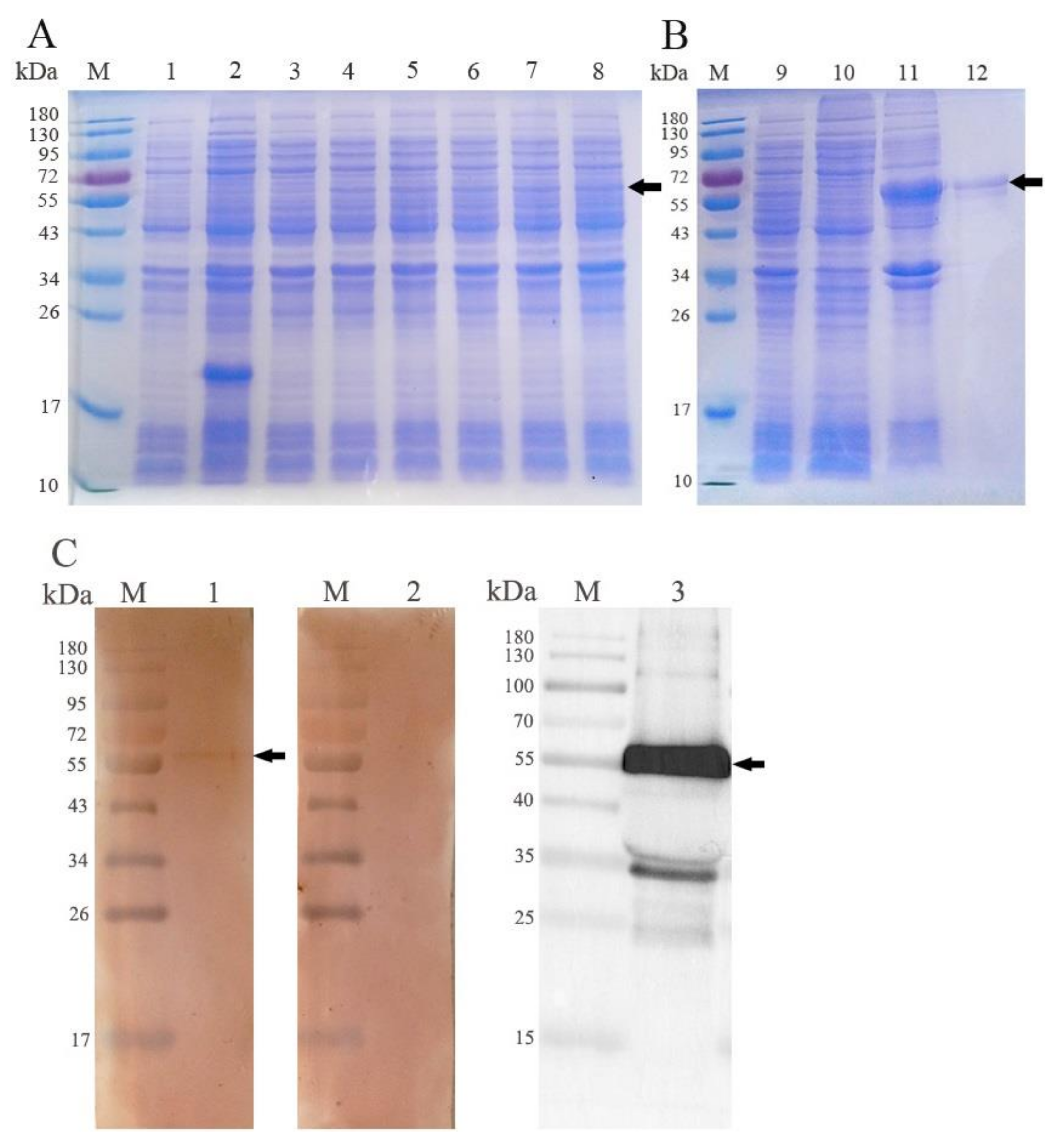
3.2. Expression and Western Blot Analysis of rEmROM5
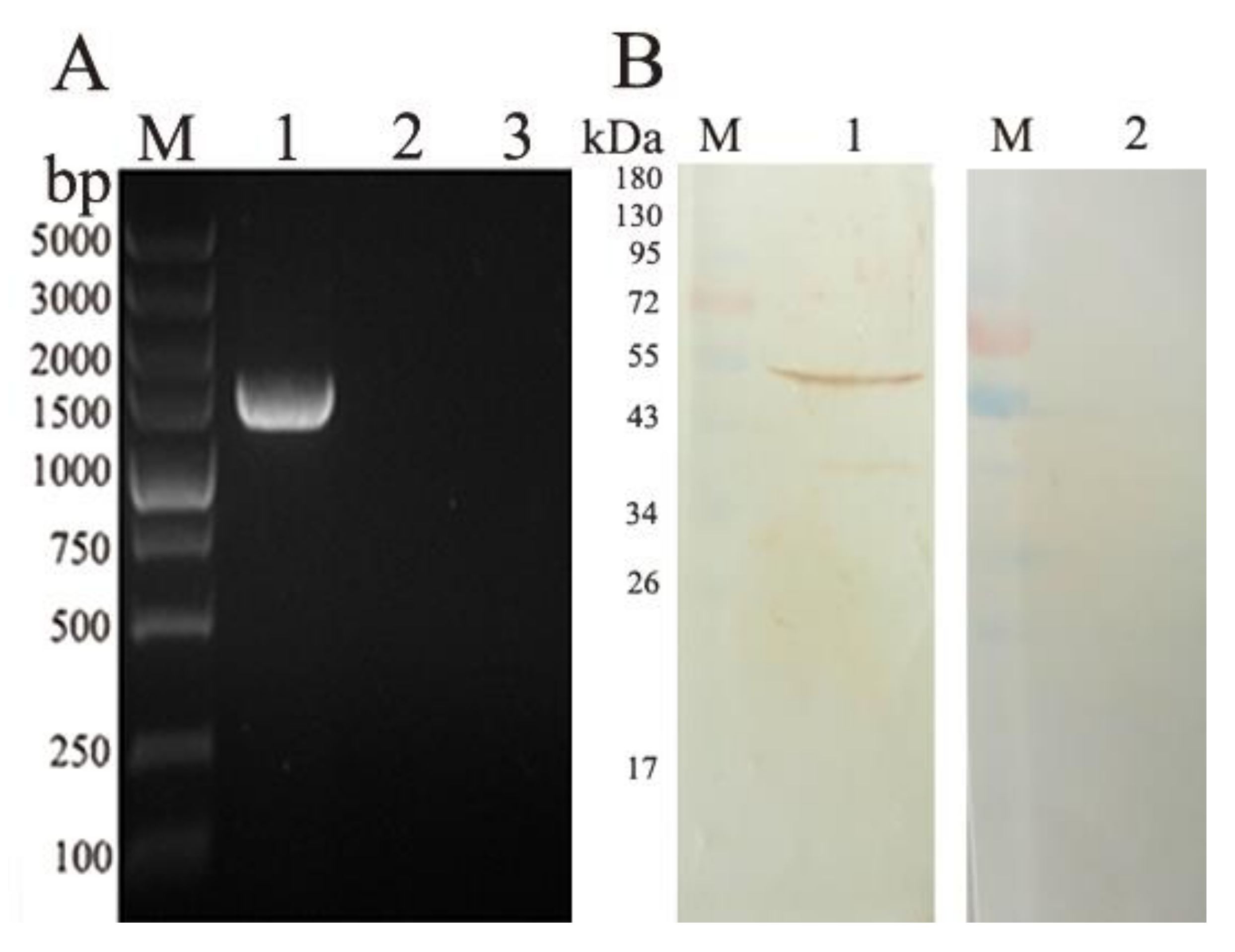
3.3. Transcription and Expression of pVAX-EmROM5 in the Injection Site Muscles of Chickens
3.4. Changes of CD4+/CD3+ and CD8+/CD3+ T Lymphocyte Subpopulation in the EmROM5 Immunized Chickens
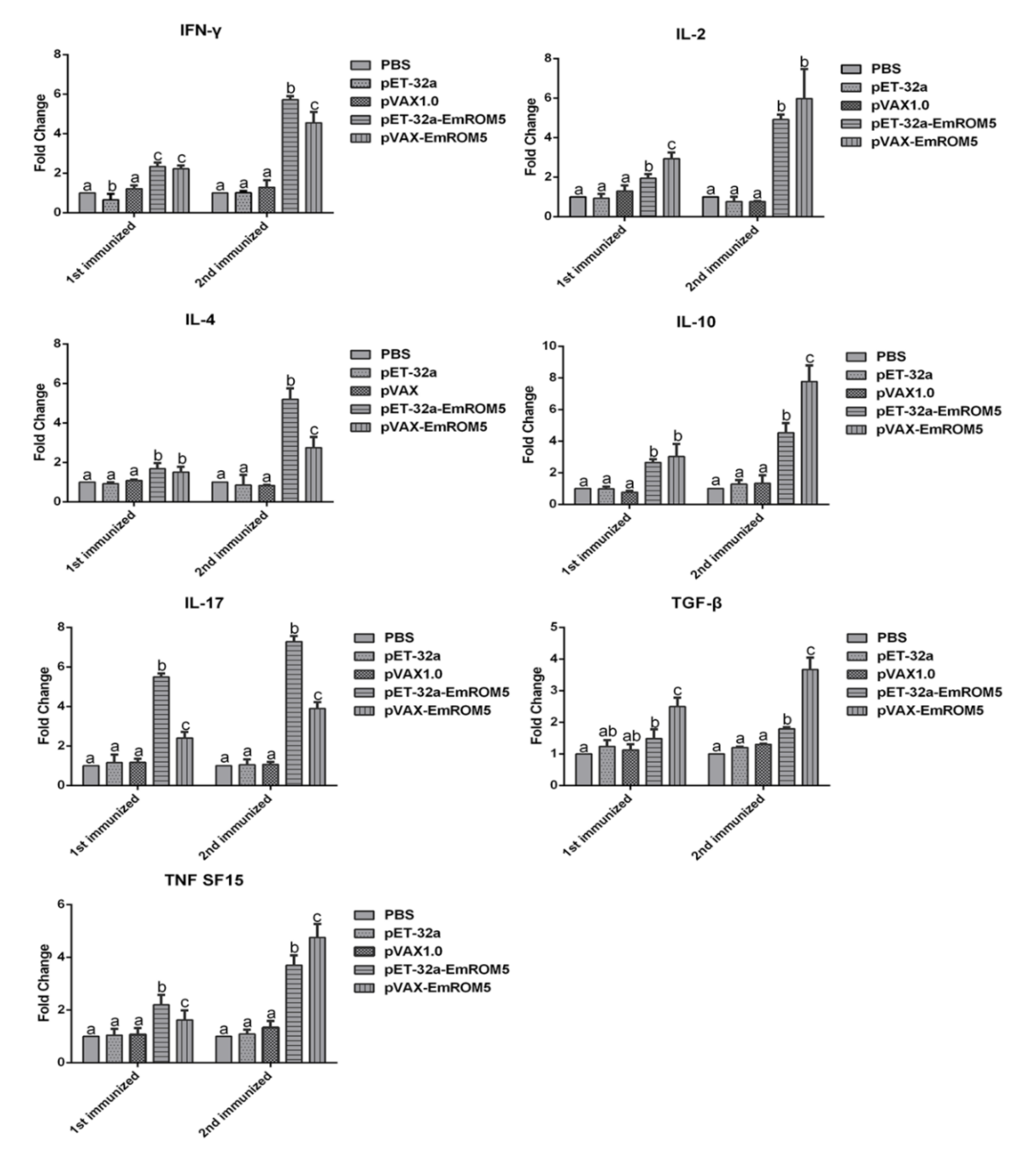
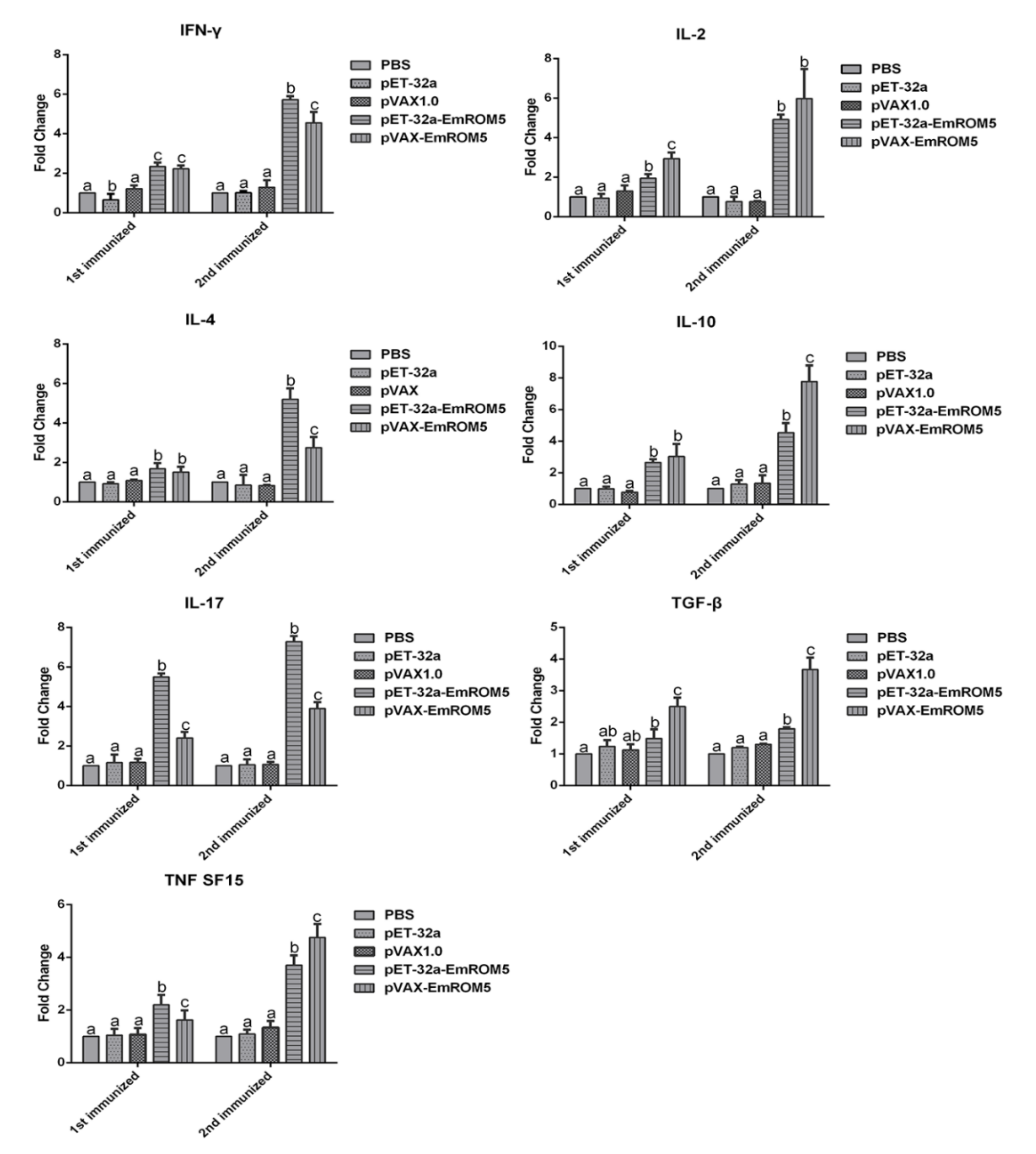
3.5. Changes of Cytokines Transcription in Splenic Lymphocytes in the EmROM5 Immunized Chickens
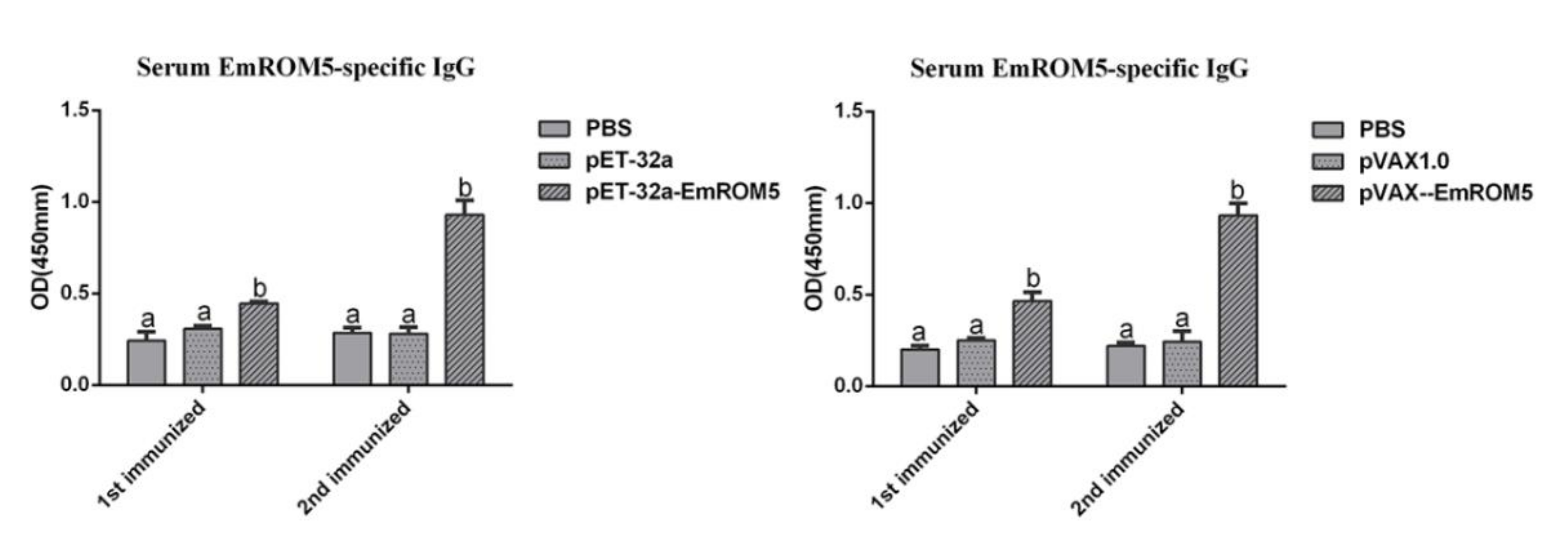
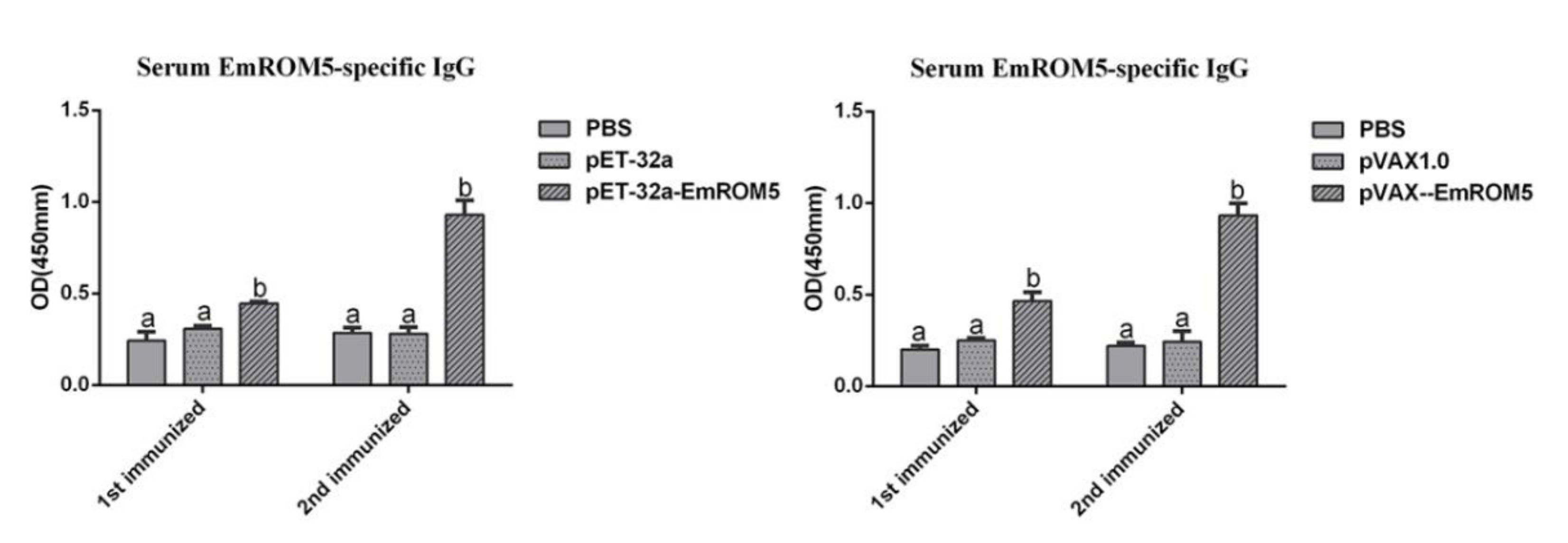
3.6. Specific Antibody IgG Levels in Chicken Serum after Immunization
3.7. Protective Efficacy of EmROM5 against Challenge with E. maxima
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Lillehoj, H.S.; Ruff, M.D.; Bacon, L.D.; Lamont, S.J.; Jeffers, T.K. Genetic control of immunity to Eimeria tenella. Interaction of MHC genes and non-MHC linked genes influences levels of disease susceptibility in chickens. Vet. Immunol. Immunopathol. 1989, 20, 135–148. [Google Scholar] [CrossRef]
- Song, X.K.; Gao, Y.L.; Xu, L.X.; Yan, R.F.; Li, X.R. Partial protection against four species of chicken coccidia induced by multivalent subunit vaccine. Vet. Parasitol. 2015, 212, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Blake, D.P.; Knox, J.; Dehaeck, B.; Huntington, B.; Rathinam, T.; Ravipati, V.; Ayoade, S.; Gilbert, W.; Adebambo, A.O.; Jatau, I.D.; et al. Re-calculating the cost of coccidiosis in chickens. Vet. Res. 2020, 51, 115. [Google Scholar] [CrossRef] [PubMed]
- Lan, L.H.; Sun, B.B.; Zuo, B.X.Z.; Chen, X.Q.; Du, A.F. Prevalence and drug resistance of avian Eimeria species in broiler chicken farms of Zhejiang province, China. Poult. Sci. 2017, 96, 2104–2109. [Google Scholar] [CrossRef]
- Williams, R.B.; Bushell, A.C.; Reperant, J.M.; Doy, T.G.; Morgan, J.H.; Shirley, M.W.; Yvore, P.; Carr, M.M.; Fremont, Y. A survey of Eimeria species in commercially-reared chickens in France during 1994. Avian Pathol. 1996, 25, 113–130. [Google Scholar] [CrossRef]
- Jamal, G.; Zivar, S.D.; Mohammadali, B. Prevalence of coccidiosis in broiler chicken farms in Western Iran. J. Vet. Med. 2014, 2014, 980604. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.Y.; Ruan, X.C.; Li, L.; Zeng, M.H. Prevalence of Eimeria species in domestic chickens in Anhui province, China. J. Parasit. Dis. 2017, 41, 1014–1019. [Google Scholar] [CrossRef]
- Sun, X.M.; Pang, W.; Jia, T.; Yan, W.C.; He, G.; Hao, L.L.; Bentué, M.; Suo, X. Prevalence of Eimeria species in broilers with subclinical signs from fifty farms. Avian Dis. 2009, 53, 301–305. [Google Scholar] [CrossRef]
- Godwin, R.M.; Morgan, J.A.T. A molecular survey of Eimeria in chickens across Australia. Vet. Parasitol. 2015, 214, 16–21. [Google Scholar] [CrossRef] [Green Version]
- Moraes, J.C.; França, M.; Sartor, A.A.; Bellato, V.; de Moura, A.B.; de Lourdes Borba Magalhães, M.; de Souza, A.P.; Miletti, L.C. Prevalence of Eimeria spp. in broilers by multiplex PCR in the southern region of Brazil on two hundred and fifty farms. Avian Dis. 2015, 59, 277–281. [Google Scholar] [CrossRef]
- Williams, R.B. Anticoccidial vaccines for broiler chickens: Pathways to success. Avian Pathol. 2002, 31, 317–353. [Google Scholar] [CrossRef] [PubMed]
- Allen, P.C.; Fetterer, R.H. Recent advances in biology and immunobiology of Eimeria species and in diagnosis and control of infection with these coccidian parasites of poultry. Clin. Microbiol. Rev. 2002, 15, 58–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapman, H.D. Resistance to anticoccidial drugs in fowl. Parasitol. Today. 1993, 9, 159–162. [Google Scholar] [CrossRef]
- Ahmad, T.A.; El-Sayed, B.A.; El-Sayed, L.H. Development of immunization trials against Eimeria spp. Trials Vaccinol. 2016, 5, 38–47. [Google Scholar] [CrossRef] [Green Version]
- Santos, J.M.; Graindorge, A.; Soldati-favre, D. New insights into parasite rhomboid proteases. Mol. Biochem. Parasitol. 2012, 182, 27–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brossier, F.; Starnes, G.L.; Beatty, W.L.; Sibley, L.D. Microneme rhomboid protease TgROM1 is required for efficient intracellular growth of Toxoplasma gondii. Eukaryot. Cell 2008, 7, 664–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J.; Gong, P.T.; Jia, H.L.; Li, M.Y.; Zhang, G.C.; Zhang, X.C.; Li, J.H. Eimeria tenella rhomboid 3 has a potential role in microneme protein cleavage. Vet. Parasitol. 2014, 201, 146–149. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.L.; Li, J.H.; Zhang, X.C.; Zhao, Q.; Liu, Q.; Gong, P.T. Eimeria tenella: Construction of a recombinant fowlpox virus expressing rhomboid gene and its protective efficacy against homologous infection. Exp. Parasitol. 2008, 119, 30–36. [Google Scholar] [CrossRef]
- Li, J.H.; Zheng, J.; Gong, P.T.; Zhang, X.C. Efficacy of Eimeria tenella rhomboid-like protein as a subunit vaccine in protective immunity against homologous challenge. Parasitol. Res. 2012, 110, 1139–1145. [Google Scholar] [CrossRef]
- Li, J.H.; Han, Q.Z.; Gong, P.T.; Yang, T.; Ren, B.Y.; Li, S.J.; Zhang, X.C. Toxoplasma gondii rhomboid protein 1 (TgROM1) is a potential vaccine candidate against toxoplasmosis. Vet. Parasitol. 2012, 184, 154–160. [Google Scholar] [CrossRef]
- Zhang, N.Z.; Xu, Y.; Wang, M.; Petersen, E.; Chen, J.; Huang, S.Y.; Zhu, X.Q. Protective efficacy of two novel DNA vaccines expressing Toxoplasma gondii rhomboid 4 and rhomboid 5 proteins against acute and chronic toxoplasmosis in mice. Expert Rev. Vaccines 2015, 14, 1289–1297. [Google Scholar] [CrossRef]
- Tomley, F. Techniques for isolation and characterization of apical organelles from Eimeria tenella sporozoites. Methods 1997, 13, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Song, H.Y.; Yan, R.F.; Xu, L.X.; Song, X.K.; Shah, M.A.; Zhu, H.L.; Li, X.R. Efficacy of DNA vaccines carrying Eimeria acervulina lactate dehydrogenase antigen gene against coccidiosis. Exp. Parasitol. 2010, 126, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, H.; Chapot, R.; Banzet, S.; Koulmann, N.; Birot, O.; Bigard, A.X.; Peinnequin, A. Quantification by real-time PCR of developmental and adult myosin mRNA in rat muscles. Biochem. Biophys. Res. Commun. 2006, 340, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Kenneth, J.L.; Thomas, D.S. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Song, X.K.; Zhao, X.F.; Xu, L.X.; Yan, R.F.; Li, X.R. Immune protection duration and efficacy stability of DNA vaccine encoding Eimeria tenella TA4 and chicken IL-2 against coccidiosis. Res. Vet. Sci. 2017, 111, 31–35. [Google Scholar] [CrossRef]
- Holdsworth, P.A.; Conway, D.P.; McKenzie, M.E.; Dayton, A.D.; Chapman, H.D.; Mathis, G.F.; Skinner, J.T.; Mundt, H.C.; Williams, R.B.; World association for the advancement of veterinary parasitology. World association for the advancement of veterinary parasitology (WAAVP) guidelines for evaluating the efficacy of anticoccidial drugs in chickens and turkeys. Vet. Parasitol. 2004, 121, 189–212. [Google Scholar] [CrossRef]
- Johnson, J.; Reid, W.M. Anticoccidial drugs: Lesion scoring techniques in battery and floor-pen experiments with chickens. Exp. Parasitol. 1970, 28, 30–36. [Google Scholar] [CrossRef]
- Rose, M.E.; Mockett, A.P. Antibodies to coccidia: Detection by the enzyme-linked immunosorbent assay (ELISA). Parasite Immunol. 1983, 5, 479–489. [Google Scholar] [CrossRef]
- Talebi, A.; Mulcahy, G. Partial protection against Eimeria acervulina and Eimeria tenella induced by synthetic peptide vaccine. Exp. Parasitol. 2005, 110, 342–348. [Google Scholar] [CrossRef]
- Mcmanus, E.C.; Campbell, W.C.; Cuckler, A.C. Development of resistance to quinoline coccidiostats under field and laboratory conditions. J. Parasitol. 1968, 54, 1190–1193. [Google Scholar] [CrossRef]
- Dalloul, R.A.; Lillehoj, H.S. Poultry coccidiosis: Recent advancements in control measures and vaccine development. Expert Rev. Vaccines 2006, 5, 143–163. [Google Scholar] [CrossRef]
- Ivory, C.; Chadee, K. DNA vaccines: Designing strategies against parasitic infections. Genet. Vaccines Ther. 2004, 2, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blake, D.P.; Tomley, F.M. Securing poultry production from the ever-present Eimeria challenge. Trends Parasitol. 2014, 30, 12–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, X.K.; Ren, Z.; Yan, R.F.; Xu, L.X.; Li, X.R. Induction of protective immunity against Eimeria tenella, Eimeria necatrix, Eimeria maxima and Eimeria acervulina infections using multivalent epitope DNA vaccines. Vaccine 2015, 33, 2764–2770. [Google Scholar] [CrossRef] [PubMed]
- Meunier, M.; Chemaly, M.; Dory, D. DNA vaccination of poultry: The current status in 2015. Vaccine 2016, 34, 202–211. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.W.; Zhang, Z.C.; Li, M.H.; Song, X.K.; Yan, R.F.; Xu, L.X.; Li, X.R. Eimeria maxima microneme protein 2 delivered as DNA vaccine and recombinant protein induces immunity against experimental homogenous challenge. Parasitol. Int. 2015, 64, 408–416. [Google Scholar] [CrossRef]
- Jang, S.I.; Lillehoj, H.S.; Lee, S.H.; Lee, K.W.; Park, M.S.; Cha, S.R.; Lillehoj, E.P.; Subramanian, B.M.; Sriraman, R.; Srinivasan, V.A. Eimeria maxima recombinant Gam82 gametocyte antigen vaccine protects against coccidiosis and augments humoral and cell-mediated immunity. Vaccine 2010, 28, 2980–2985. [Google Scholar] [CrossRef] [PubMed]
- Lamarque, M.; Besteiro, S.; Papoin, J.; Roques, M.; Vulliez-Le Normand, B.; Morlon-Guyot, J.; Dubremetz, J.F.; Fauquenoy, S.; Tomavo, S.; Faber, B.W.; et al. The RON2-AMA1 interaction is a critical step in moving junction-dependent invasion by apicomplexan parasites. PLoS Pathog. 2011, 7, e1001276. [Google Scholar] [CrossRef] [Green Version]
- Jia, L.J.; Guo, H.P.; Liu, M.M.; Gao, Y.; Zhang, L.; Li, H.; Xie, S.Z.; Zhang, N.N. Construction of an adenovirus vaccine expressing the cross-reactive antigen AMA1 for Neospora caninum and Toxoplasma gondii and its immune response in an animal model. Iran. J. Parasitol. 2018, 13, 235–243. [Google Scholar]
- Remarque, E.J.; Faber, B.W.; Kocken, C.H.; Thomas, A.W. Apical membrane antigen 1: A malaria vaccine candidate in review. Trends Parasitol. 2008, 24, 74–84. [Google Scholar] [CrossRef]
- Tomley, F.M.; Soldati, D.S. Mix and match modules: Structure and function of microneme proteins in apicomplexan parasites. Trends Parasitol. 2001, 17, 81–88. [Google Scholar] [CrossRef]
- Qi, N.S.; Wang, Y.Y.; Liao, S.Q.; Wu, C.Y.; Lv, M.N.; Li, J.; Tong, Z.X.; Sun, M.F. Partial protective of chickens against Eimeria tenella challenge with recombinant EtMIC-1 antigen. Parasitol. Res. 2013, 112, 2281–2287. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.Y.; Cui, X.X.; Zhao, Q.P.; Zhu, S.H.; Huang, B.; Wang, L.; Zhao, H.Z.; Liu, G.L.; Li, Z.H.; Han, H.Y.; et al. Molecular characterization and protective efficacy of the microneme 2 protein from Eimeria tenella. Parasite 2018, 25, 60. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.C.; Liu, L.R.; Huang, J.W.; Wang, S.; Lu, M.M.; Song, X.K.; Xu, L.X.; Yan, R.F.; Li, X.R. The molecular characterization and immune protection of microneme 2 of Eimeria acervulina. Vet. Parasitol. 2016, 215, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.M.; Liu, J.H.; Tian, D.; Li, W.Y.; Zhou, Z.Y.; Huang, J.M.; Song, X.K.; Xu, L.X.; Yan, R.F.; Li, X.R. The molecular characterization and protective efficacy of microneme 3 of Eimeria mitis in chickens. Vet. Parasitol. 2018, 258, 114–123. [Google Scholar] [CrossRef]
- Dowse, T.J.; Pascall, J.C.; Brown, K.D.; Soldati, D. Apicomplexan rhomboids have a potential role in microneme protein cleavage during host cell invasion. Int. J. Parasitol. 2005, 35, 747–756. [Google Scholar] [CrossRef] [Green Version]
- Ha, Y. Structure and mechanism of intramembrane protease. Semin. Cell Dev. Biol. 2009, 20, 240–250. [Google Scholar] [CrossRef] [Green Version]
- Lillehoj, H.S.; Trout, J.M. Avian gut-associated lymphoid tissues and intestinal immune responses to Eimeria parasites. Clin. Microbiol. Rev. 1996, 9, 349–360. [Google Scholar] [CrossRef]
- Yun, C.H.; Lillehoj, H.S.; Lillehoj, E.P. Intestinal immune responses to coccidiosis. Dev. Comp. Immunol. 2000, 24, 303–324. [Google Scholar] [CrossRef]
- Lillehoj, H.S.; Lillehoj, E.P. Avian coccidiosis. A review of acquired intestinal immunity and vaccination strategies. Avian Dis. 2000, 44, 408–425. [Google Scholar] [CrossRef] [PubMed]
- Lillehoj, H.S.; Ruff, M.D. Comparison of disease susceptibility and subclass-specific antibody response in SC and FP chickens experimentally inoculated with Eimeria tenella, E. acervulina, or E. maxima. Avian Dis. 1987, 31, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Wallach, M.; Halabi, A.; Pillemer, G.; Sar-Shalom, O.; Mencher, D.; Gilad, M.; Bendheim, U.; Danforth, H.D.; Augustine, P.C. Maternal immunization with gametocyte antigens as a means of providing protective immunity against Eimeria maxima in chickens. Infect. Immun. 1992, 60, 2036–2039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witcombe, D.M.; Smith, N.C. Strategies for anticoccidial prophylaxis. Parasitology 2014, 141, 1379–1389. [Google Scholar] [CrossRef] [PubMed]
- Chapman, H.D. Milestones in avian coccidiosis research: A review. Poult. Sci. 2014, 93, 501–511. [Google Scholar] [CrossRef]
- Lillehoj, H.S.; Min, W.; Dalloul, R.A. Recent progress on the cytokine regulation of intestinal immune responses to Eimeria. Poult. Sci. 2004, 83, 611–623. [Google Scholar] [CrossRef]
- Kim, W.H.; Chaudhari, A.A.; Lillehoj, H.S. Involvement of T cell immunity in avian coccidiosis. Front. Immunol. 2019, 10, 2732. [Google Scholar] [CrossRef] [Green Version]
- Annamalai, T.; Selvaraj, R.K. Effects of in ovo interleukin-4-plasmid injection on anticoccidia immune response in a coccidia infection model of chickens. Poult. Sci. 2012, 91, 1326–1334. [Google Scholar] [CrossRef]
- Min, W.; Kim, W.H.; Lillehoj, E.P.; Lillehoj, H.S. Recent progress in host immunity to avian coccidiosis: IL-17 family cytokines as sentinels of the intestinal mucosa. Dev. Comp. Immunol. 2013, 41, 418–428. [Google Scholar] [CrossRef]
- Zhang, S.; Lillehoj, H.S.; Ruff, M.D. Chicken tumor necrosis-like factor.: 1. in vitro production by macrophages stimulated with Eimeria tenella or bacterial lipopolysaccharide. Poult. Sci. 1995, 74, 1304–1310. [Google Scholar] [CrossRef]
- Jakowlew, S.B.; Mathias, A.; Lillehoj, H.S. Transforming growth factor-beta isoforms in the developing chicken intestine and spleen: Increase in transforming growth factor-beta 4 with coccidia infection. Vet. Immunol. Immunopathol. 1997, 55, 321–339. [Google Scholar] [CrossRef]
- Arendt, M.K.; Sand, J.M.; Marcone, T.M.; Cook, M.E. Interleukin-10 neutralizing antibody for detection of intestinal luminal levels and as a dietary additive in Eimeria challenged broiler chicks. Poult. Sci. 2016, 95, 430–438. [Google Scholar] [CrossRef]
- Min, W.; Lillehoj, H.S.; Burnside, J.; Weining, K.C.; Staeheli, P.; Zhu, J.J. Adjuvant effects of IL-1β, IL-2, IL-8, IL-15, IFN-α, IFN-γ, TGF-β4 and lymphotactin on DNA vaccination against Eimeria acervulina. Vaccine 2001, 20, 267–274. [Google Scholar] [CrossRef]
- Lillehoj, H.S.; Ding, X.C.; Quiroz, M.A.; Bevensee, E.; Lillehoj, E.P. Resistance to intestinal coccidiosis following DNA immunization with the cloned 3-1E Eimeria gene plus IL-2, IL-15, and IFN-gamma. Avian Dis. 2005, 49, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Song, X.K.; Huang, X.M.; Yan, R.F.; Xu, L.X.; Li, X.R. Efficacy of chimeric DNA vaccines encoding Eimeria tenella 5401 and chicken IFN-γ or IL-2 against coccidiosis in chickens. Exp. Parasitol. 2015, 156, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Wallach, M.; Smith, N.C.; Petracca, M.; Miller, C.M.; Eckert, J.; Braun, R. Eimeria maxima gametocyte antigens: Potential use in a subunit maternal vaccine against coccidiosis in chickens. Vaccine 1995, 13, 347–354. [Google Scholar] [CrossRef]
- Jang, S.I.; Lillehoj, H.S.; Lee, S.H.; Lee, K.W.; Lillehoj, E.P.; Bertrand, F.; Dupuis, L.; Deville, S. Montanide IMS 1313 N VG PR nanoparticle adjuvant enhances antigen-specific immune responses to profilin following mucosal vaccination against Eimeria acervulina. Vet. Parasitol. 2011, 182, 163–170. [Google Scholar] [CrossRef]
- Song, X.K.; Xu, L.X.; Yan, R.F.; Huang, X.M.; Shah, M.A.; Li, X.R. The optimal immunization procedure of DNA vaccine pcDNA-TA4-IL-2 of Eimeria tenella and its cross-immunity to Eimeria necatrix and Eimeria acervulina. Vet. Parasitol. 2009, 159, 30–36. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Size (bp 3) |
|---|---|---|
| ntmEmROM5 1 | Forward: 5’-CCGGAATTCATGTCTTCCCCCATTG-3’ | 963 |
| Reverse: 5’-CCCTCGAGATGCAAAAAGGAGGCCCAAAAGAC-3’ | ||
| EmROM5 2 | Forward: 5’-CCGGAATTCATGTCTTCCCCCATTG-3’ | 1461 |
| Reverse: 5’-AAATATGCGGCCGCTCAAGTAAACTT-3’ |
| RNA Target | Primer Sequence | Accession No. | Amplification Efficiency (%) | Correlation Coefficient (r2) |
|---|---|---|---|---|
| GAPDH | GGTGGTGCTAAGCGTGTTAT | K01458 | 100.74% | 0.9917 |
| ACCTCTGTCATCTCTCCACA | ||||
| IL-2 | TAACTGGGACACTGCCATGA | AF000631 | 102.44% | 0.9921 |
| GATGATAGAGATGCTCCATAAGCTG | ||||
| IL-4 | ACCCAGGGCATCCAGAAG | AJ621735 | 99.09% | 0.9936 |
| CAGTGCCGGCAAGAAGTT | ||||
| IL-10 | GGAGCTGAGGGTGAAGTTTGA | AJ621614 | 99.19% | 0.9923 |
| GAAGCGCAGCATCTCTGACA | ||||
| IL-17 | ACCTTCCCATGTGCAGAAAT | EF570583 | 100.24% | 0.994 |
| GAGAACTGCCTTGCCTAACA | ||||
| IFN-γ | AGCTGACGGTGGACCTATTATT | Y07922 | 103.07% | 0.9868 |
| GGCTTTGCGCTGGATTC | ||||
| TGF-β | CGGGACGGATGAGAAGAAC | M31160 | 102.79% | 0.9815 |
| CGGCCCACGTAGTAAATGAT | ||||
| TNF SF15 | GCTTGGCCTTTACCAAGAAC | NM001024578 | 100.57% | 0.993 |
| GGAAAGTGACCTGAGCATAGA |
| Trials | Groups | Average Body Weight Gain (g) | Relative Body Weight Gain (%) | Mean Lesion Score | Average OPG (×105) | ACI |
|---|---|---|---|---|---|---|
| 1 | Unchallenged control | 56.91 ± 10.24 a | 100 a | 0 ± 0 a | 0 ± 0 a | 200 |
| Challenged control | 27.21 ± 8.52 c | 47.81 c | 2.84 ± 0.88 c | 2.25 ± 0.94 c | 79.41 | |
| pET-32a tag protein control | 29.46 ± 11.25 c | 51.77 c | 2.66 ± 0.93 c | 2.15 ± 0.97 c | 85.17 | |
| rEmROM5 | 49.36 ± 11.35 b | 86.73 b | 1.46 ± 0.52 b | 0.56 ± 0.48 b | 171.13 | |
| 2 | Unchallenged control | 79.32 ± 9.59 a | 100 a | 0 ± 0 a | 0 ± 0 a | 200 |
| Challenged control | 39.28 ± 9.72 c | 49.53 c | 2.83 ± 0.72 c | 2.81 ± 0.13 c | 81.23 | |
| pVAX1.0 control | 38.19 ± 15.39 c | 48.15 c | 2.75 ± 0.62 c | 2.80 ± 0.16 c | 80.65 | |
| pVAX–EmROM5 | 65.95 ± 4.96 b | 83.14 b | 1.25 ± 0.75 b | 0.67 ± 0.19 b | 169.64 |
| Marker | Groups | 1st Immunization | 2nd Immunization |
|---|---|---|---|
| CD4+/CD3+ | PBS buffer | 18.27 ± 0.21 a | 19.47 ± 3.21 a |
| pET-32a tag protein | 20.90 ± 1.64 a | 23.28 ± 1.62 a | |
| rEmROM5 | 27.75 ± 1.35 bc | 30.07 ± 0.57 b | |
| PBS buffer | 10.18 ± 0.87 a | 14.13 ± 1.50 a | |
| pVAX1.0 | 11.50 ± 2.05 a | 16.45 ± 2.05 a | |
| pVAX-EmROM5 | 22.57 ± 1.85 b | 27.51 ± 4.95 b | |
| CD8+/CD3+ | PBS buffer | 21.45 ± 0.72 a | 22.80 ± 5.20 a |
| pET-32a tag protein | 22.37 ± 1.53 a | 25.93 ± 3.39 a | |
| rEmROM5 | 35.20 ± 5.20 b | 43.59 ± 6.76 b | |
| PBS buffer | 13.00 ± 1.65 a | 13.78 ± 1.61 a | |
| pVAX1.0 | 15.60 ± 1.13 b | 17.24 ± 0.42 a | |
| pVAX-EmROM5 | 22.37 ± 0.17 c | 31.40 ± 4.48 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, D.; Liu, X.; Li, X.; Xu, L.; Yan, R.; Song, X. Eimeria maxima Rhomboid-like Protein 5 Provided Partial Protection against Homologous Challenge in Forms of Recombinant Protein and DNA Plasmid in Chickens. Vaccines 2022, 10, 32. https://doi.org/10.3390/vaccines10010032
Tian D, Liu X, Li X, Xu L, Yan R, Song X. Eimeria maxima Rhomboid-like Protein 5 Provided Partial Protection against Homologous Challenge in Forms of Recombinant Protein and DNA Plasmid in Chickens. Vaccines. 2022; 10(1):32. https://doi.org/10.3390/vaccines10010032
Chicago/Turabian StyleTian, Di, Xiaoqian Liu, Xiangrui Li, Lixin Xu, Ruofeng Yan, and Xiaokai Song. 2022. "Eimeria maxima Rhomboid-like Protein 5 Provided Partial Protection against Homologous Challenge in Forms of Recombinant Protein and DNA Plasmid in Chickens" Vaccines 10, no. 1: 32. https://doi.org/10.3390/vaccines10010032
APA StyleTian, D., Liu, X., Li, X., Xu, L., Yan, R., & Song, X. (2022). Eimeria maxima Rhomboid-like Protein 5 Provided Partial Protection against Homologous Challenge in Forms of Recombinant Protein and DNA Plasmid in Chickens. Vaccines, 10(1), 32. https://doi.org/10.3390/vaccines10010032