
Toxoplasma gondii GRA15 DNA Vaccine with a Liposomal Nanocarrier Composed of an SS-Cleavable and pH-Activated Lipid-like Material Induces Protective Immunity against Toxoplasmosis in Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Mice
2.3. Cultures and Purification of T. gondii
2.4. Cloning of the TgGRA15 Gene
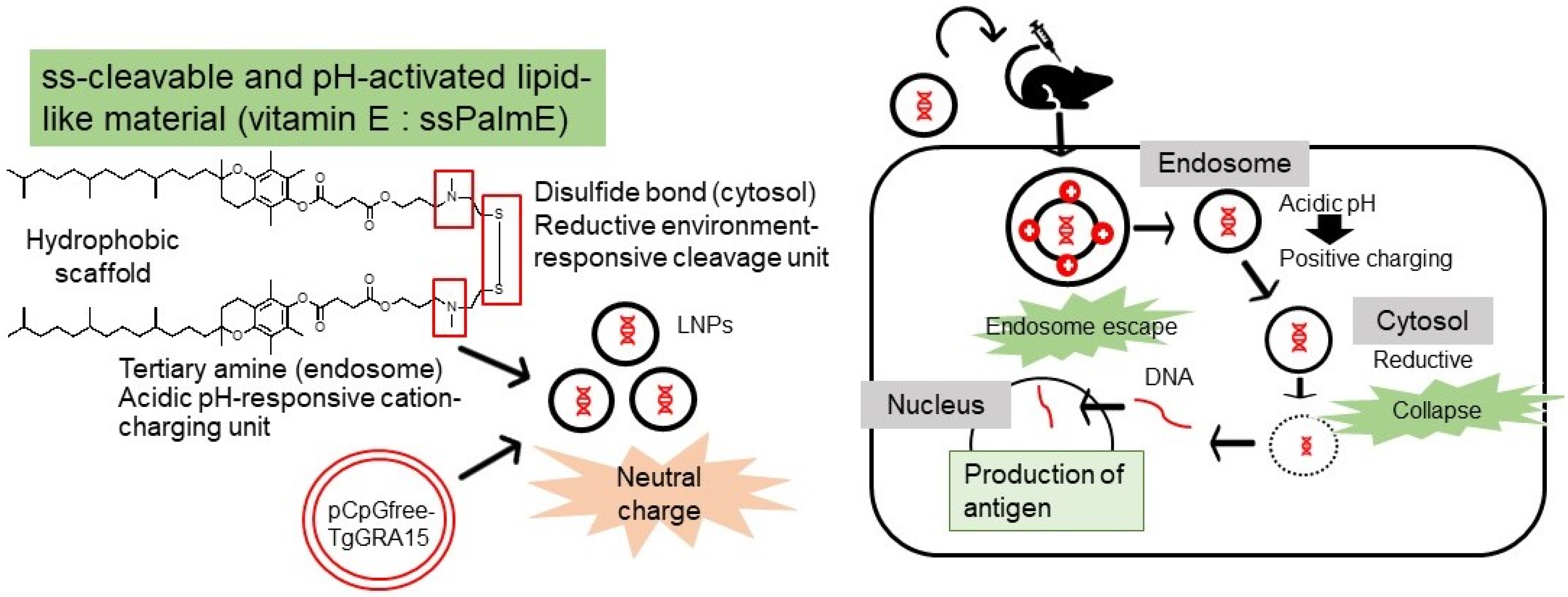
2.5. Preparation of LNP-Encapsulated pCpG-free-TgGRA15
2.6. Immunization and Infection in Mice
2.7. Measurement of TgGRA15-Specific Antibodies
2.8. Measurement of IFN-Gamma (IFN-γ)
2.9. Clinical Score
2.10. Statistical Analysis
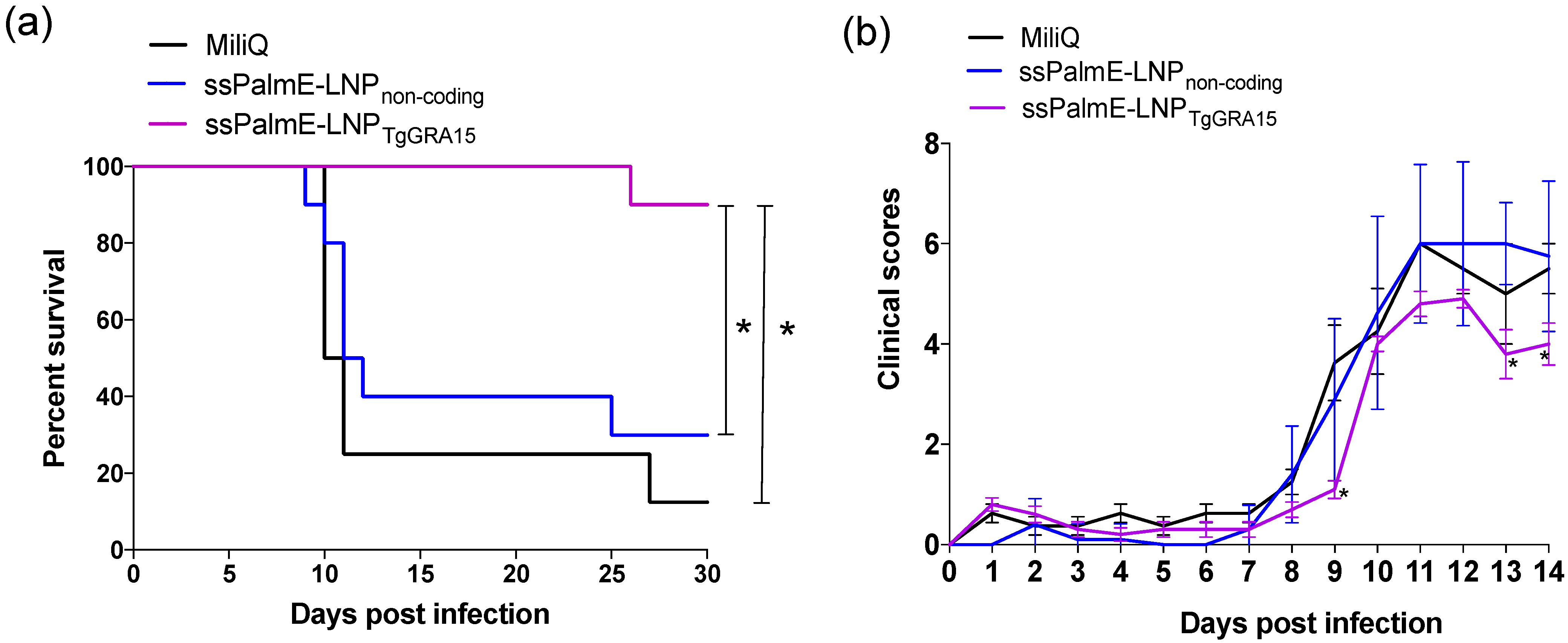
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dubey, J.P. Toxoplasmosis of Animals and Humans, 2nd ed.; CRC Press, Inc.: Boca Raton, NY, USA, 2010; p. 313. [Google Scholar] [CrossRef]
- Montoya, J.G.; Liesenfeld, O. Toxoplasmosis. Lancet 2004, 363, 1965–1976. [Google Scholar] [CrossRef]
- Weiss, L.M.; Dubey, J.P. Toxoplasmosis: A history of clinical observations. Int. J. Parasitol. 2009, 39, 895–901. [Google Scholar] [CrossRef] [PubMed]
- Havelaar, A.H.; Kemmeren, J.M.; Kortbeek, L.M. Disease burden of congenital toxoplasmosis clinical infectious diseases. Clin. Infect Dis. 2007, 44, 1467–1474. [Google Scholar] [CrossRef]
- Buzby, J.C.; Roberts, T. Economic costs and trade impacts of microbial foodborne illness. World Health Stat. Q. 1997, 50, 57–66. [Google Scholar] [PubMed]
- Tenter, A.M.; Heckeroth, A.R.; Weiss, L.M. Toxoplasma gondii: From animals to humans. Int. J. Parasitol. 2000, 30, 12–13. [Google Scholar] [CrossRef]
- Stelzer, S.; Basso, W.; Benavides Silván, J.; Ortega-Mora, L.M.; Maksimov, P.; Gethmann, J.; Conraths, F.J.; Schares, G. Toxoplasma gondii infection and toxoplasmosis in farm animals: Risk factors and economic impact. Food Waterborne Parasitol. 2019, 15, 1–31. [Google Scholar] [CrossRef]
- Jongert, E.; Roberts, C.W.; Gargano, N.; Förster-Wald, E.; Petersen, E. Vaccines against Toxoplasma gondii: Challenges and opportunities. Mem. Inst. Oswaldo Cruz. 2009, 104, 252–266. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.L. Vaccination concepts against Toxoplasma gondii. Expert Rev. Vaccines 2009, 8, 215–225. [Google Scholar] [CrossRef]
- Zukancic, D.; Suys, E.J.A.; Pilkington, E.H.; Algarni, A.; Al-Wassiti, H.; Nghia, P.T. The Importance of poly (ethylene glycol) and lipid structure in targeted gene delivery to lymph nodes by lipid nanoparticles. Pharmaceutics 2020, 12, 1068. [Google Scholar] [CrossRef]
- Wolff, J.A.; Budker, V. The Mechanism of Naked DNA Uptake and Expression. Adv. Genet. 2005, 54, 2–20. [Google Scholar] [CrossRef]
- Rodrigues, M.M.; Boscardin, S.B.; Vasconcelos, J.R.; Hiyane, M.I.; Salay, G.; Soares, I.S. Importance of CD8 T cell-mediated immune response during intracellular parasitic infections and its implications for the development of effective vaccines. An. Da Acad. Bras. De Cienc. 2003, 75, 443–468. [Google Scholar] [CrossRef]
- Lechardeur, D.; Sohn, K.-J.; Haardt, M.; Joshi, P.B.; Monck, M.; Graham, R.W.; Beatty, B.; Squire, J.; O’Brodovich, H.; Lukacs, G.L. Metabolic instability of plasmid DNA in the cytosol: A potential barrier to gene transfer. Gene. Ther. 1999, 6, 482–497. [Google Scholar] [CrossRef]
- Ruponen, M. Extracellular and intracellular barriers in non-viral gene delivery. J. Control Release 2003, 93, 213–217. [Google Scholar] [CrossRef]
- Akita, H.; Ishiba, R.; Hatakeyama, H.; Tanaka, H.; Sato, Y.; Tange, K.; Arai, M.; Kubo, K.; Harashima, H. A Neutral Envelope-Type Nanoparticle Containing pH-esponsive and SS-Cleavable Lipid-Like Material as a Carrier for Plasmid DNA. Adv. Healthc. Mater. 2013, 2, 1120–1125. [Google Scholar] [CrossRef]
- Kawai, M.; Nakamura, T.; Miura, N.; Maeta, M.; Tanaka, H.; Ueda, K.; Higashi, K.; Moribe, K.; Tange, K.; Nakai, Y.; et al. DNA-loaded nano-adjuvant formed with a vitamin E-scaffold intracellular environmentally-responsive lipid-like material for cancer immunotherapy. Nanomed. Nanotechnol. 2018, 14, 2587–2597. [Google Scholar] [CrossRef]
- Maeta, M.; Miura, N.; Tanaka, H.; Nakamura, T.; Kawanishi, R.; Nishikawa, Y.; Asano, K.; Tanaka, M.; Tamagawa, S.; Nakai, Y.; et al. Vitamin E Scaffolds of pH-Responsive Lipid Nanoparticles as DNA Vaccines in Cancer and Protozoan. Mol. Pharm. 2020, 17, 1237–1247. [Google Scholar] [CrossRef]
- Rosowski, E.E.; Lu, D.; Julien, L.; Rodda, L.; Gaiser, R.A.; Jensen, K.D.C.; Saeij, J.P.J. Strain-specific activation of the NF-κB pathway by GRA15, a novel Toxoplasma gondii dense granule protein. J. Exp. Med. 2011, 208, 195–212. [Google Scholar] [CrossRef]
- Gov, L.; Karimzadeh, A.; Ueno, N.; Lodoen, M.B. Human Innate Immunity to Toxoplasma gondii is mediated by host caspase-1 and ASC and Parasite GRA15. mBio 2013, 4, 1–13. [Google Scholar] [CrossRef]
- Miura, N.; Shaheen, S.M.; Akita, H.; Nakamura, T.; Harashima, H. A KALA-modified lipid nanoparticle containing CpG-free plasmid DNA as a potential DNA vaccine carrier for antigen presentation and as an immune-stimulative adjuvant. Nucleic Acids. Res. 2015, 43, 1317–1331. [Google Scholar] [CrossRef]
- Ihara, F.; Fereig, R.M.; Himori, Y.; Kameyama, K.; Umeda, K.; Tanaka, S.; Ikeda, R.; Yamamoto, M.; Nishikawa, Y. Toxoplasma gondii Dense Granule Proteins 7, 14, and 15 Are involved in modification and control of the immune response mediated via NF-κB pathway. Front. Immunol. 2020, 11, 1–18. [Google Scholar] [CrossRef]
- Terkawi, M.A.; Kameyama, K.; Rasul, N.H.; Xuan, X.; Nishikawa, Y. Development of an immunochromatographic assay based on dense granule protein 7 for serological detection of Toxoplasma gondii infection. Clin. Vaccine Immunol. 2013, 20, 596–601. [Google Scholar] [CrossRef] [PubMed]
- Rogers, D.C.; Fisher, E.M.; Brown, S.D.; Peters, J.; Hunter, A.H.; Martin, J.E. Behavioral and functional analysis of mouse phenotype: SHIRPA, a proposed protocol for comprehensive phenotype assessment. Mamm. Genome 1997, 10, 711–713. [Google Scholar] [CrossRef]
- Fereig, R.M.; Shimoda, N.; Abdelbaky, H.H.; Kuroda, Y.; Nishikawa, Y. Neospora GRA6 possesses immune-stimulating activity and confers efficient protection against Neospora caninum infection in mice. Vet. Parasitol. 2019, 267, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Akita, H.; Ishiba, R.; Tange, K.; Arai, M.; Kubo, K.; Harashima, H. Neutral biodegradable lipid-envelope-type nanoparticle using vitamin A-Scaffold for nuclear targeting of plasmid DNA. Biomaterials 2014, 35, 1755–1761. [Google Scholar] [CrossRef] [PubMed]
- Akita, H.; Ishiba, R.; Togashi, R.; Tange, K.; Nakai, Y.; Hatakeyama, H.; Harashima, H. A neutral lipid envelope-type nanoparticle composed of a pH-activated and vitamin E-scaffold lipid-like material as a platform for a gene carrier targeting renal cell carcinoma. J. Control. Release 2015, 200, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Morel, S.; Didierlaurent, A.; Bourguignon, P.; Delhaye, S.; Baras, B.; Jacob, V.; Planty, C.; Elouahabi, A.; Harvengt, P.; Carlsen, H. Adjuvant System AS03 containing α-tocopherol modulates innate immune response and leads to improved adaptive immunity. Vaccine 2011, 29, 2461–2473. [Google Scholar] [CrossRef] [PubMed]
- DiDonato, J.A.; Mercurio, F.; Karin, M. NF-κB and the link between inflammation and cancer. Immunol. Rev. 2012, 246, 379–400. [Google Scholar] [CrossRef]
- Wang, P.; Li, S.; Zhao, Y.; Zhang, B.; Li, Y.; Liu, S.; Du, H.; Cao, L.; Ou, M.; Ye, X.; et al. The GRA15 protein from Toxoplasma gondii enhances host defense responses by activating the interferon stimulator STING. J. Biol. Chem. 2019, 294, 16494–16508. [Google Scholar] [CrossRef]
- Denis, F.; Ermanno, C. Immune response to Toxoplasma gondii. Ann. Acad. Bras. Cienc. 2004, 40, 71–80. [Google Scholar]
- Gazzinelli, R.T.; Hieny, S.; Wynn, T.A.; Wolf, S.; Sher, A. Interleukin 12 is required for the T-lymphocyte-independent induction of interferon gamma by an intracellular parasite and induces resistance in T-cell-deficient hosts. Proc. Natl. Acad. Sci. USA 1993, 90, 6115–6119. [Google Scholar] [CrossRef]
- Johnson, L.L.; Jessica, H.; Chen, W. Vaccination protects B cell-deficient mice against an oral challenge with mildly virulent Toxoplasma gondii. Vaccine 2004, 22, 4054–4061. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, Z.Y.; Petersen, E.; Huang, S.Y.; Zhou, D.H.; Zhu, X.Q. DNA vaccination with genes encoding Toxoplasma gondii antigens ROP5 and GRA15 induces protective immunity against toxoplasmosis in Kunming mice. Expert Rev. Vaccines 2015, 14, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Petrushina, I.; Tran, M.; Sadzikava, N.; Ghochikyan, A.; Vasilevko, V.; Agadjanyan, M.G.; Cribbs, D.H. Importance of IgG2c isotype in the immune response to β-amyloid in amyloid precursor protein/transgenic mice. Neurosci. Lett. 2003, 338, 5–8. [Google Scholar] [CrossRef]
- Petritus, P.M.; Burns, J.M. Suppression of lethal Plasmodium yoelii malaria following protective immunization requires antibody IL-4, and IFN-γ-dependent responses induced by vaccination and/or challenge infection. J. Immunol. 2008, 180, 444–453. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, F.; Wang, G.; Zhao, Q.; Min, J.; Wang, S.; Cong, H.; Li, Y.; He, S.; Zhou, H. Toxoplasma gondii Immune response and protective efficacy induced by ROP16/GRA7 multicomponent DNA vaccine with a genetic adjuvant B7-2. Hum. Vaccin. Immunother. 2014, 10, 184–191. [Google Scholar] [CrossRef][Green Version]
- Nielsen, H.V.; Lauemùller, S.; Christiansen, L.; Buus, S.; Fomsgaard, A.; Petersen, E. Complete protection against lethal Toxoplasma gondii infection in mice immunized with a plasmid encoding the SAG1 gene. Infect. Immun. 1999, 67, 6358–6363. [Google Scholar] [CrossRef]
- Angus, C.W.; Klivington-Evans, D.; Dubey, J.P.; Kovacs, J.A. Immunization with a DNA plasmid encoding the SAG1 /P30) protein of Toxoplasma gondii is immunogenic and protective in rodents. J. Infect. Dis. 2000, 181, 317–324. [Google Scholar] [CrossRef]
- Vercammen, M.; Scorza, T.; Huygen, K.; De Braekeleer, J.; Diet, R.; Jacobs, D.; Saman, E.; Verschueren, H. DNA vaccination with genes encoding Toxoplasma gondii antigens GRA1, GRA7, and ROP2 induces partially protective immunity against lethal challenge in mice. Infect. Immun. 2000, 68, 38–45. [Google Scholar] [CrossRef]
- Echeverria, P.C.; de Miguel, N.; Costas, M.; Angel, S.O. Potent antigen-specific immunity to Toxoplasma gondii in adjuvant-free vaccination system using Rop2-Leishmania infantum Hsp83 fusion protein. Vaccine 2006, 24, 4102–4110. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hasan, T.; Kawanishi, R.; Akita, H.; Nishikawa, Y. Toxoplasma gondii GRA15 DNA Vaccine with a Liposomal Nanocarrier Composed of an SS-Cleavable and pH-Activated Lipid-like Material Induces Protective Immunity against Toxoplasmosis in Mice. Vaccines 2022, 10, 21. https://doi.org/10.3390/vaccines10010021
Hasan T, Kawanishi R, Akita H, Nishikawa Y. Toxoplasma gondii GRA15 DNA Vaccine with a Liposomal Nanocarrier Composed of an SS-Cleavable and pH-Activated Lipid-like Material Induces Protective Immunity against Toxoplasmosis in Mice. Vaccines. 2022; 10(1):21. https://doi.org/10.3390/vaccines10010021
Chicago/Turabian StyleHasan, Tanjila, Ryo Kawanishi, Hidetaka Akita, and Yoshifumi Nishikawa. 2022. "Toxoplasma gondii GRA15 DNA Vaccine with a Liposomal Nanocarrier Composed of an SS-Cleavable and pH-Activated Lipid-like Material Induces Protective Immunity against Toxoplasmosis in Mice" Vaccines 10, no. 1: 21. https://doi.org/10.3390/vaccines10010021
APA StyleHasan, T., Kawanishi, R., Akita, H., & Nishikawa, Y. (2022). Toxoplasma gondii GRA15 DNA Vaccine with a Liposomal Nanocarrier Composed of an SS-Cleavable and pH-Activated Lipid-like Material Induces Protective Immunity against Toxoplasmosis in Mice. Vaccines, 10(1), 21. https://doi.org/10.3390/vaccines10010021