
Immune Enhancement by the Tetra-Peptide Hydrogel as a Promising Adjuvant for an H7N9 Vaccine against Highly Pathogenic H7N9 Virus

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
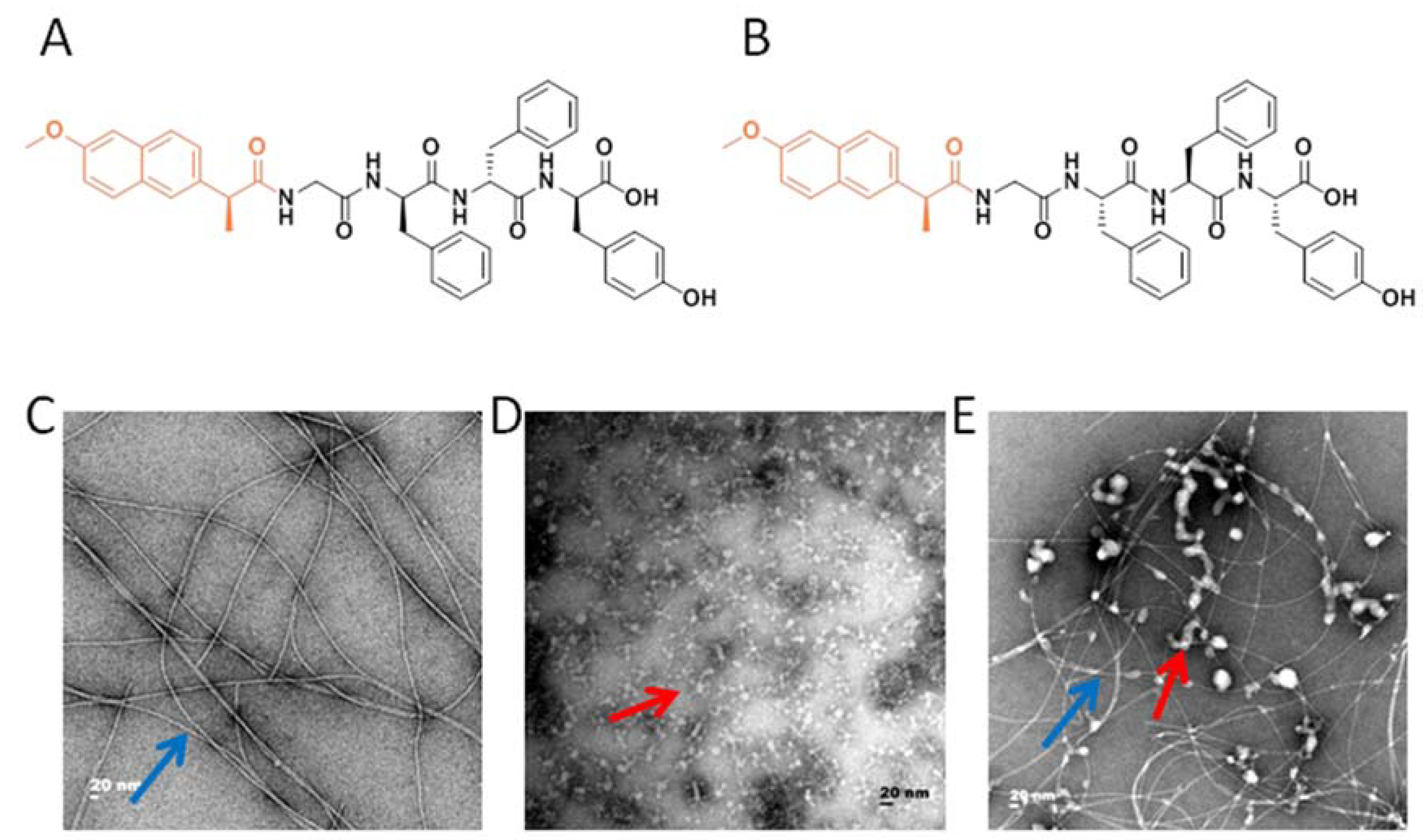
2.1. Peptide Synthesis and Hydrogel Preparation
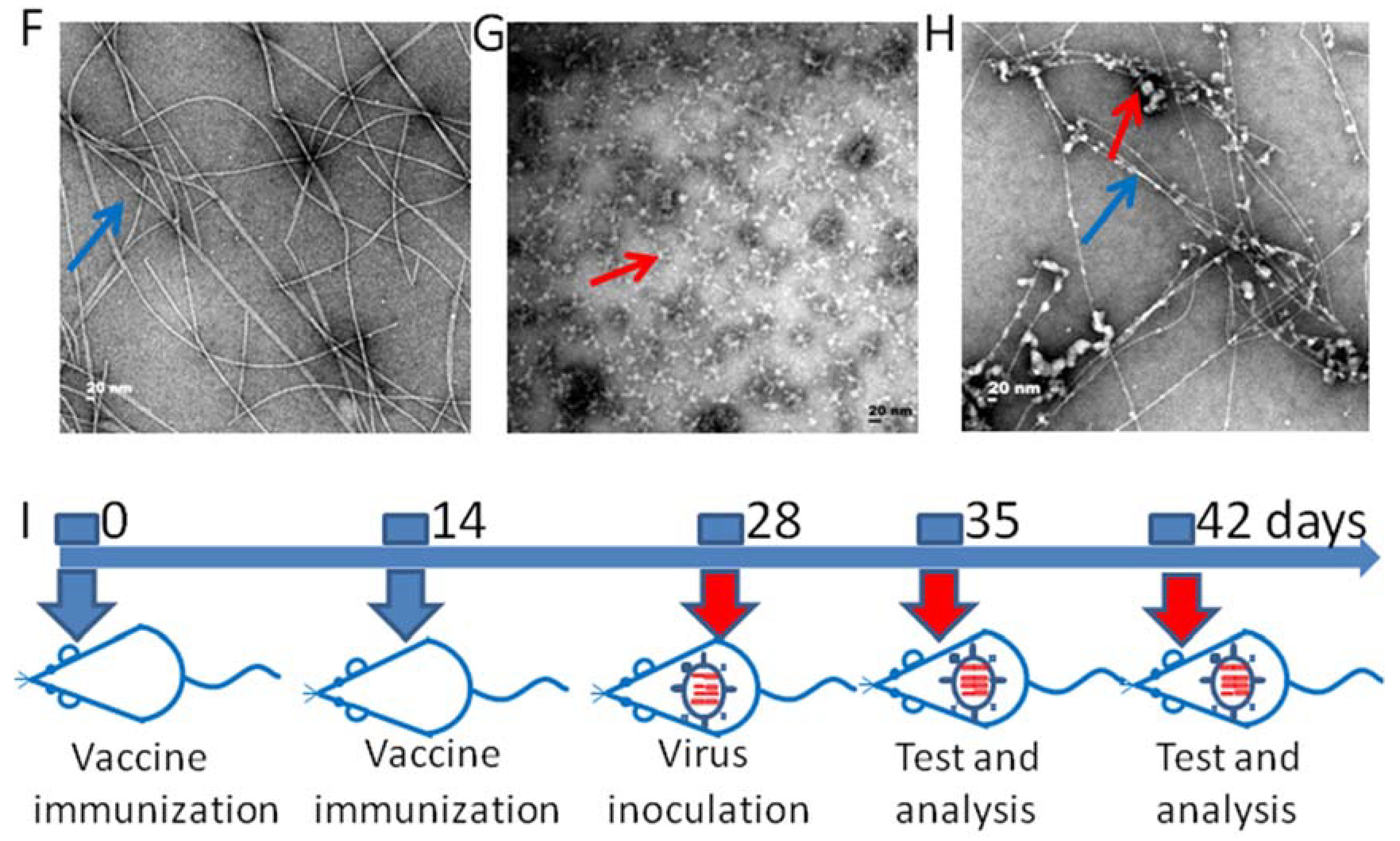
2.2. Transmission Electron Microscopy (TEM)
2.3. Experimental Animal
2.4. Cells and Viruses
2.5. Animal Immunization and Virus Inoculation
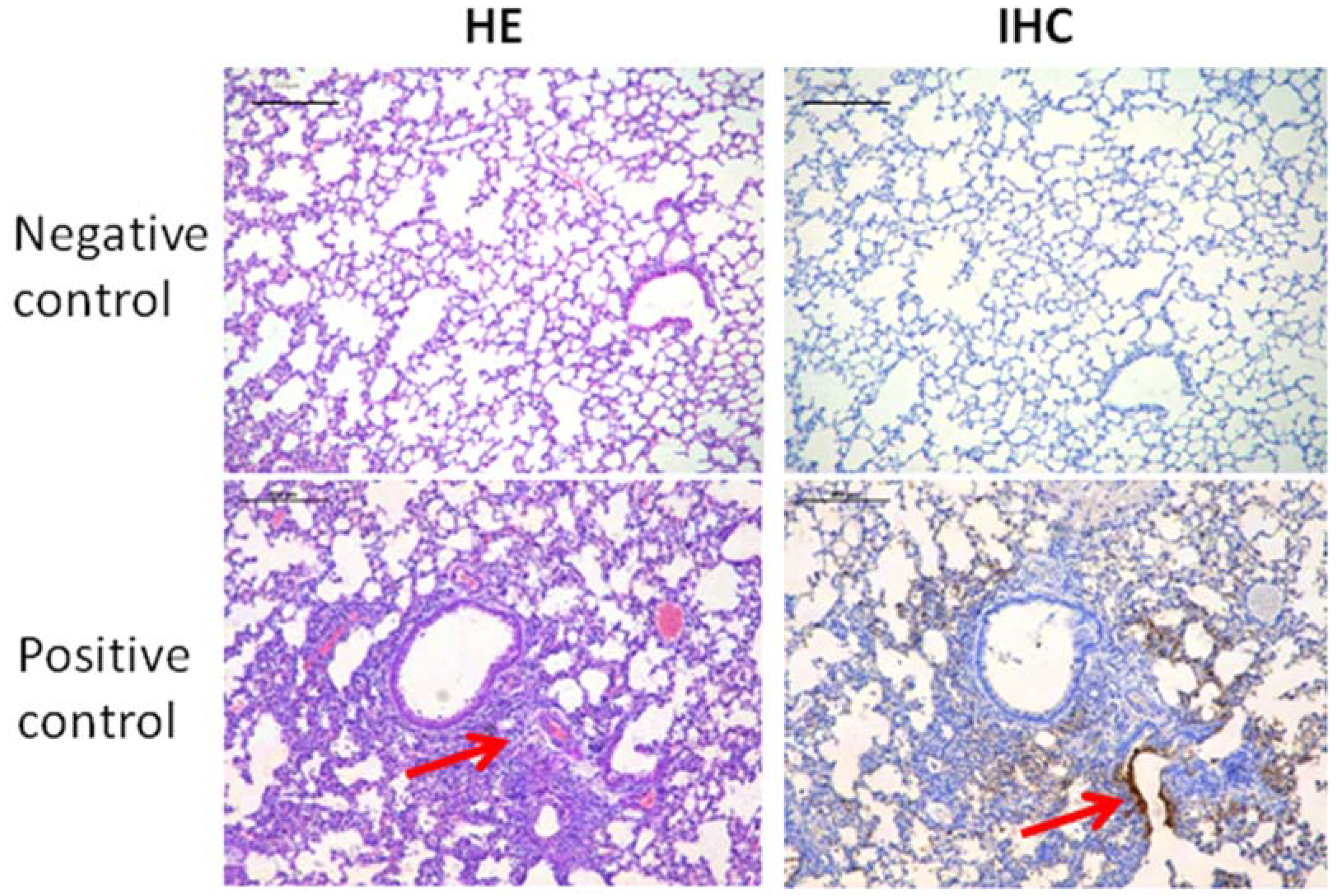
2.6. Histopathological Analysis of Lung Tissues
2.7. Immunoglobulin G Enzyme-Linked Immunosorbent Assay (IgG-ELISA)
2.8. Hemagglutination Inhibition (HI) Titer Assay
2.9. Micro-Neutralization (MN) Assay
2.10. Statistical Analysis
3. Results
3.1. The Preparation of the Hydrogel Adjuvant Vaccine
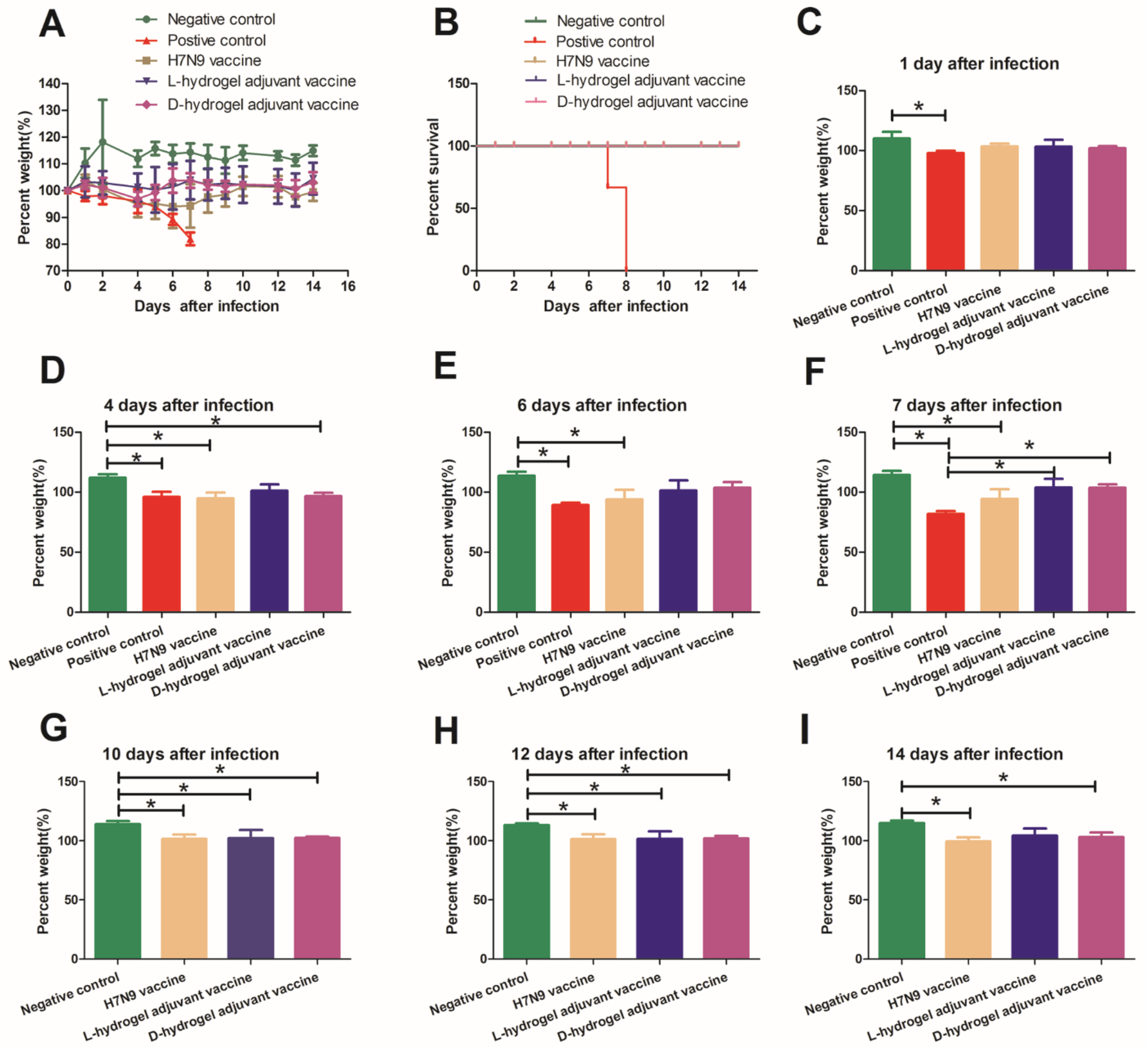
3.2. The Protective Effects of the Hydrogel Adjuvant Vaccine In Vivo
3.3. The Protective Effects of Hydrogel Adjuvant Vaccine against Lung Damage
3.4. The Hydrogel Adjuvant Vaccine Elicited Good Protective Antibody Titers against H7N9 Virus
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grabowski, M.L.; Kosinska, B.; Knap, J.P.; Brydak, L.B. The Lethal Spanish Influenza Pandemic in Poland. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2017, 23, 4880–4884. [Google Scholar] [CrossRef] [PubMed]
- El-Kafrawy, S.A.; Corman, V.M.; Tolah, A.M.; Al Masaudi, S.B.; Hassan, A.M.; Muller, M.A.; Bleicker, T.; Harakeh, S.M.; Alzahrani, A.A.; Alsaaidi, G.A.; et al. Enzootic patterns of Middle East respiratory syndrome coronavirus in imported African and local Arabian dromedary camels: A prospective genomic study. Lancet Planet. Health 2019, 3, e521–e528. [Google Scholar] [CrossRef]
- Zhou, F.; Yu, T.; Du, R.; Fan, G.; Liu, Y.; Liu, Z.; Xiang, J.; Wang, Y.; Song, B.; Gu, X.; et al. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: A retrospective cohort study. Lancet 2020, 395, 1054–1062. [Google Scholar] [CrossRef]
- Wang, L.F.; Anderson, D.E.; Mackenzie, J.S.; Merson, M.H. From Hendra to Wuhan: What has been learned in responding to emerging zoonotic viruses. Lancet 2020, 395, e33–e34. [Google Scholar] [CrossRef]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef]
- Puntasecca, C.J.; King, C.H.; LaBeaud, A.D. Measuring the global burden of chikungunya and Zika viruses: A systematic review. PLoS Negl. Trop. Dis. 2021, 15, e0009055. [Google Scholar] [CrossRef]
- Diseases, G.B.D.; Injuries, C. Global burden of 369 diseases and injuries in 204 countries and territories, 1990-2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet 2020, 396, 1204–1222. [Google Scholar]
- Iuliano, A.D.; Roguski, K.M.; Chang, H.H.; Muscatello, D.J.; Palekar, R.; Tempia, S.; Cohen, C.; Gran, J.M.; Schanzer, D.; Cowling, B.J.; et al. Estimates of global seasonal influenza-associated respiratory mortality: A modelling study. Lancet 2018, 391, 1285–1300. [Google Scholar] [CrossRef]
- Dodd, P.J.; Yuen, C.M.; Jayasooriya, S.M.; van der Zalm, M.M.; Seddon, J.A. Quantifying the global number of tuberculosis survivors: A modelling study. Lancet Infect. Dis. 2021, 21, 984–992. [Google Scholar] [CrossRef]
- Wu, Q.; Zaid, M.; Xuan, Z.; Wang, C.; Gu, H.; Shi, M.; Zhu, J.; Hu, Y.; Liu, J. Changes in epidemiological features of vaccine preventable infectious diseases among three eras of national vaccination strategies from 1953 to 2018 in Shanghai, China. Lancet Reg. Health West. Pac. 2021, 7, 100092. [Google Scholar] [CrossRef]
- Damaso, C.R. Revisiting Jenner’s mysteries, the role of the Beaugency lymph in the evolutionary path of ancient smallpox vaccines. Lancet Infect. Dis. 2018, 18, e55–e63. [Google Scholar] [CrossRef]
- Anand, S.; Barnighausen, T. Health workers and vaccination coverage in developing countries: An econometric analysis. Lancet 2007, 369, 1277–1285. [Google Scholar] [CrossRef]
- GBD RVCC. Measuring routine childhood vaccination coverage in 204 countries and territories, 1980–2019: A systematic analysis for the Global Burden of Disease Study 2020, Release 1. Lancet 2021, 398, 503–521. [Google Scholar] [CrossRef]
- The Lancet. Eliminating viral hepatitis: Time to match visions with action. Lancet 2017, 390, 2121. [Google Scholar] [CrossRef]
- Li, Y.; Zhou, Y.; Cheng, Y.; Wu, P.; Zhou, C.; Cui, P.; Song, C.; Liang, L.; Wang, F.; Qiu, Q.; et al. Effectiveness of EV-A71 vaccination in prevention of paediatric hand, foot, and mouth disease associated with EV-A71 virus infection requiring hospitalisation in Henan, China, 2017–2018: A test-negative case-control study. Lancet Child Adolesc. Health 2019, 3, 697–704. [Google Scholar] [CrossRef]
- Li, X.; Mukandavire, C.; Cucunuba, Z.M.; Echeverria Londono, S.; Abbas, K.; Clapham, H.E.; Jit, M.; Johnson, H.L.; Papadopoulos, T.; Vynnycky, E.; et al. Estimating the health impact of vaccination against ten pathogens in 98 low-income and middle-income countries from 2000 to 2030: A modelling study. Lancet 2021, 397, 398–408. [Google Scholar] [CrossRef]
- Qin, T.; Ma, S.; Miao, X.; Tang, Y.; Huangfu, D.; Wang, J.; Jiang, J.; Xu, N.; Yin, Y.; Chen, S.; et al. Mucosal Vaccination for Influenza Protection Enhanced by Catalytic Immune-Adjuvant. Adv. Sci. 2020, 7, 2000771. [Google Scholar] [CrossRef]
- Wang, Y.; Xie, Y.; Luo, J.; Guo, M.; Hu, X.; Chen, X.; Chen, Z.; Lu, X.; Mao, L.; Zhang, K.; et al. Engineering a self-navigated MnARK nanovaccine for inducing potent protective immunity against novel coronavirus. Nano Today 2021, 38, 101139. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Aldayel, A.M.; Cui, Z. Aluminum hydroxide nanoparticles show a stronger vaccine adjuvant activity than traditional aluminum hydroxide microparticles. J. Control. Release 2014, 173, 148–157. [Google Scholar] [CrossRef]
- Tang, S.; Liu, Z.; Xu, W.; Li, Q.; Han, T.; Pan, D.; Yue, N.; Wu, M.; Liu, Q.; Yuan, W.; et al. Versatile Functionalization of Ferritin Nanoparticles by Intein-Mediated Trans-Splicing for Antigen/Adjuvant Co-delivery. Nano Lett. 2019, 19, 5469–5475. [Google Scholar] [CrossRef]
- Yang, Z.; Liang, G.; Xu, B. Enzymatic control of the self-assembly of small molecules: A new way to generate supramolecular hydrogels. Soft Matter 2007, 3, 515–520. [Google Scholar] [CrossRef]
- Yang, Z.; Liang, G.; Xu, B. Enzymatic hydrogelation of small molecules. Acc. Chem. Res. 2008, 41, 315–326. [Google Scholar] [CrossRef]
- Luo, Z.; Wu, Q.; Yang, C.; Wang, H.; He, T.; Wang, Y.; Wang, Z.; Chen, H.; Li, X.; Gong, C.; et al. A Powerful CD8(+) T-Cell Stimulating D-Tetra-Peptide Hydrogel as a Very Promising Vaccine Adjuvant. Adv. Mater. 2017, 29, 1601776. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, Z.; Yi, X.; Long, J.; Liu, J.; Yang, Z. Anti-degradation of a recombinant complex protein by incorporation in small molecular hydrogels. Chem. Commun. 2011, 47, 955–957. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liang, C.; Shi, F.; He, T.; Gong, C.; Wang, L.; Yang, Z. Cancer vaccines using supramolecular hydrogels of NSAID-modified peptides as adjuvants abolish tumorigenesis. Nanoscale 2017, 9, 14058–14064. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, Y.; Zhang, X.; Hu, Y.; Yi, X.; Ma, L.; Zhou, H.; Long, J.; Liu, Q.; Yang, Z. Supramolecular nanofibers of self-assembling peptides and proteins for protein delivery. Chem. Commun. 2015, 51, 14239–14242. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.-X.; Zhao, L.-Z.; Tang, S.-J.; Weng, T.-H.; Wu, W.-G.; Yao, S.-H.; Wu, H.-B.; Cheng, L.-F.; Wang, J.; Hu, F.-Y.; et al. Novel pathogenic characteristics of highly pathogenic avian influenza virus H7N9: Viraemia and extrapulmonary infection. Emerg. Microbes Infect. 2020, 9, 962–975. [Google Scholar] [CrossRef]
- Wu, X.; Xiao, L.; Li, L. Research progress on human infection with avian influenza H7N9. Front. Med. 2020, 14, 8–20. [Google Scholar] [CrossRef]
- Ou, H.; Yao, W.; Wu, N.; Wang, F.X.C.; Weng, T.; Han, C.; Lu, X.; Yu, D.; Wu, H.; Cheng, L.; et al. Preclinical evaluation of the safety and pathogenicity of a live attenuated recombinant influenza A/H7N9 seed strain and corresponding MF59-adjuvanted split vaccine. Oncotarget 2016, 7, 81012–81025. [Google Scholar] [CrossRef][Green Version]
- Ou, H.; Yao, H.; Yao, W.; Wu, N.; Wu, X.; Han, C.; Cheng, L.; Chen, K.; Chen, H.; Li, L. Analysis of the immunogenicity and bioactivities of a split influenza A/H7N9 vaccine mixed with MF59 adjuvant in BALB/c mice. Vaccine 2016, 34, 2362–2370. [Google Scholar] [CrossRef]
- Wu, X.-X.; Deng, X.-L.; Yu, D.-S.; Yao, W.; Ou, H.-L.; Weng, T.-H.; Hu, C.-Y.; Hu, F.-Y.; Wu, N.-P.; Yao, H.-P.; et al. The Protective Effects of the A/ZJU01/PR8/2013 Split H7N9 Avian Influenza Vaccine against Highly Pathogenic H7N9 in BALB/c Mice. Cell. Physiol. Biochem. 2018, 46, 633–643. [Google Scholar] [CrossRef]
- Subbarao, K. The success of SARS-CoV-2 vaccines and challenges ahead. Cell Host Microbe 2021, 29, 1111–1123. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zeng, G.; Pan, H.; Li, C.; Hu, Y.; Chu, K.; Han, W.; Chen, Z.; Tang, R.; Yin, W.; et al. Safety, tolerability, and immunogenicity of an inactivated SARS-CoV-2 vaccine in healthy adults aged 18-59 years: A randomised, double-blind, placebo-controlled, phase 1/2 clinical trial. Lancet Infect. Dis. 2021, 21, 181–192. [Google Scholar] [CrossRef]
- Omer, S.B.; Clark, D.R.; Madhi, S.A.; Tapia, M.D.; Nunes, M.C.; Cutland, C.L.; Simões, E.A.F.; Aqil, A.R.; Katz, J.; Tielsch, J.M.; et al. Efficacy, duration of protection, birth outcomes, and infant growth associated with influenza vaccination in pregnancy: A pooled analysis of three randomised controlled trials. Lancet Respir. Med. 2020, 8, 597–608. [Google Scholar] [CrossRef]
- Lim, W.W.; Mak, L.; Leung, G.M.; Cowling, B.J.; Peiris, M. Comparative immunogenicity of mRNA and inactivated vaccines against COVID-19. Lancet Microbe 2021, 2, e423. [Google Scholar] [CrossRef]
- Wang, H.; Yang, Z. Short-peptide-based molecular hydrogels: Novel gelation strategies and applications for tissue engineering and drug delivery. Nanoscale 2012, 4, 5259–5267. [Google Scholar] [CrossRef]
- Abudula, T.; Bhatt, K.; Eggermont, L.J.; O’are, N.; Memic, A.; Bencherif, S.A. Supramolecular Self-Assembled Peptide-Based Vaccines: Current State and Future Perspectives. Front. Chem. 2020, 8, 598160. [Google Scholar] [CrossRef]
- Yang, C.; Ren, X.; Ding, D.; Wang, L.; Yang, Z. Enzymatic induction of supramolecular order and bioactivity. Nanoscale 2016, 8, 10768–10773. [Google Scholar] [CrossRef]
- Nakayama, T.; Kumagai, T.; Kashiwagi, Y.; Yoshii, H.; Honjo, K.; Kubota-Koketsu, R.; Okuno, Y.; Suga, S. Cytokine production in whole-blood cultures following immunization with an influenza vaccine. Hum. Vaccines Immunother. 2018, 14, 2990–2998. [Google Scholar] [CrossRef]
- Roth, G.A.; Saouaf, O.M.; Smith, A.A.A.; Gale, E.C.; Hernandez, M.A.; Idoyaga, J.; Appel, E.A. Prolonged Codelivery of Hemagglutinin and a TLR7/8 Agonist in a Supramolecular Polymer-Nanoparticle Hydrogel Enhances Potency and Breadth of Influenza Vaccination. ACS Biomater. Sci. Eng. 2021, 7, 1889–1899. [Google Scholar] [CrossRef]
- Yang, C.; Shi, F.; Li, C.; Wang, Y.; Wang, L.; Yang, Z. Single Dose of Protein Vaccine with Peptide Nanofibers As Adjuvants Elicits Long-Lasting Antibody Titer. ACS Biomater. Sci. Eng. 2018, 4, 2000–2006. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group Name | Vaccine Composition | HA | Adjuvant | Virus Inoculation |
|---|---|---|---|---|
| Negative control | PBS | − | − | − |
| Positive control | PBS | − | − | A/Guangdong/GZ8H002/2017(H7N9) virus |
| H7N9 vaccine | H7N9 vaccine | 2.5 μg/mouse | − | A/Guangdong/GZ8H002/2017(H7N9) virus |
| L-hydrogel adjuvant vaccine | L-Tetra-Peptide Hydrogel and H7N9 vaccine | 2.5 μg/mouse | 100 μL/mouse | A/Guangdong/GZ8H002/2017(H7N9) virus |
| D-hydrogel adjuvant vaccine | D-Tetra-Peptide Hydrogel and H7N9 vaccine | 2.5 μg/mouse | 100 μL/mouse | A/Guangdong/GZ8H002/2017(H7N9) virus |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, X.; Tang, S.; Wang, Z.; Ma, X.; Zhang, L.; Zhang, F.; Xiao, L.; Zhao, S.; Li, Q.; Wang, Y.; et al. Immune Enhancement by the Tetra-Peptide Hydrogel as a Promising Adjuvant for an H7N9 Vaccine against Highly Pathogenic H7N9 Virus. Vaccines 2022, 10, 130. https://doi.org/10.3390/vaccines10010130
Wu X, Tang S, Wang Z, Ma X, Zhang L, Zhang F, Xiao L, Zhao S, Li Q, Wang Y, et al. Immune Enhancement by the Tetra-Peptide Hydrogel as a Promising Adjuvant for an H7N9 Vaccine against Highly Pathogenic H7N9 Virus. Vaccines. 2022; 10(1):130. https://doi.org/10.3390/vaccines10010130
Chicago/Turabian StyleWu, Xiaoxin, Songjia Tang, Zhehua Wang, Xiaoyun Ma, Lingjian Zhang, Fen Zhang, Lanlan Xiao, Shuai Zhao, Qian Li, Ying Wang, and et al. 2022. "Immune Enhancement by the Tetra-Peptide Hydrogel as a Promising Adjuvant for an H7N9 Vaccine against Highly Pathogenic H7N9 Virus" Vaccines 10, no. 1: 130. https://doi.org/10.3390/vaccines10010130
APA StyleWu, X., Tang, S., Wang, Z., Ma, X., Zhang, L., Zhang, F., Xiao, L., Zhao, S., Li, Q., Wang, Y., Wang, Q., & Chen, K. (2022). Immune Enhancement by the Tetra-Peptide Hydrogel as a Promising Adjuvant for an H7N9 Vaccine against Highly Pathogenic H7N9 Virus. Vaccines, 10(1), 130. https://doi.org/10.3390/vaccines10010130