Comparison of In Vitro and In Vivo Antioxidant Activities of Six Flavonoids with Similar Structures



Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. In Vitro Antioxidant Activity Assays
2.2.1. DPPH• Radical Scavenging Assay
2.2.2. ABTS+ Radical Scavenging Assay
2.3. Animal Experiments
2.4. Measurement of Oxidative Stress Indicators and Inflammatory Cytokines
2.5. Real-Time Quantitative Polymerase Chain Reaction (RT-PCR) Analysis
2.6. Statistical Analysis
3. Results
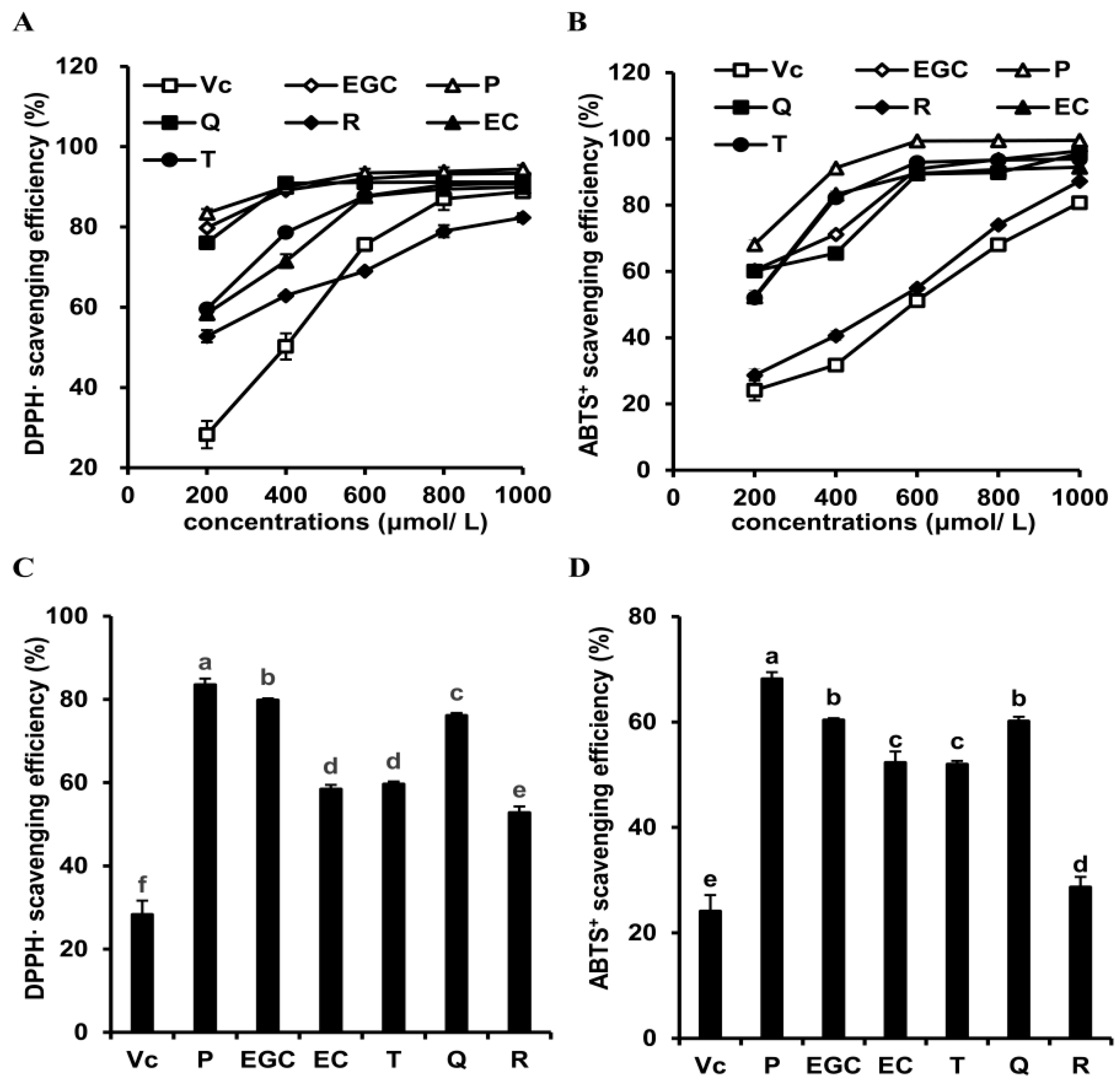
3.1. Effects of Six Flavonoids on In Vitro Antioxidant Activities
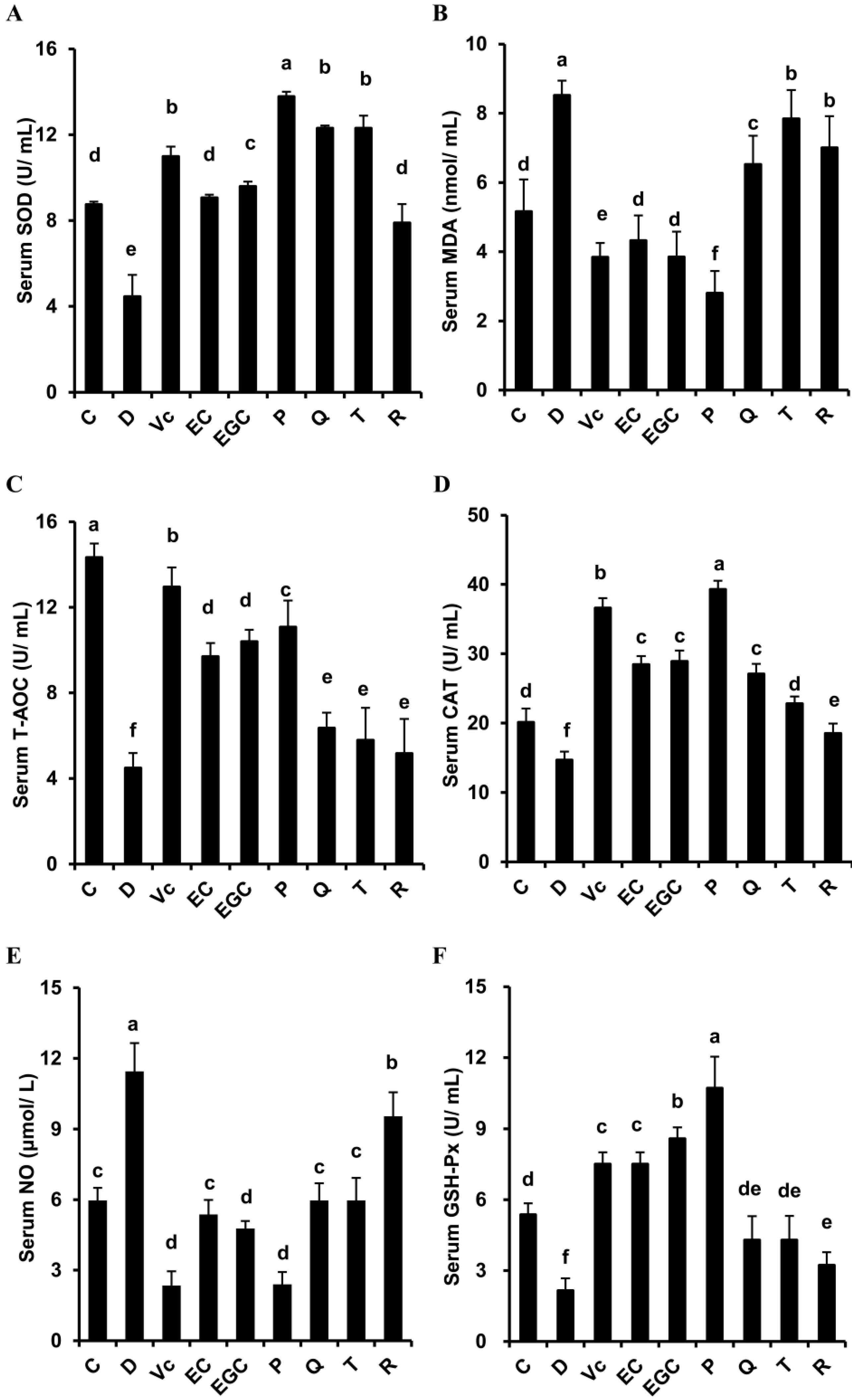
3.2. Effect of Six Flavonoids on Serum Oxidative Stress Indicators In d-Galactose-Treated Mice
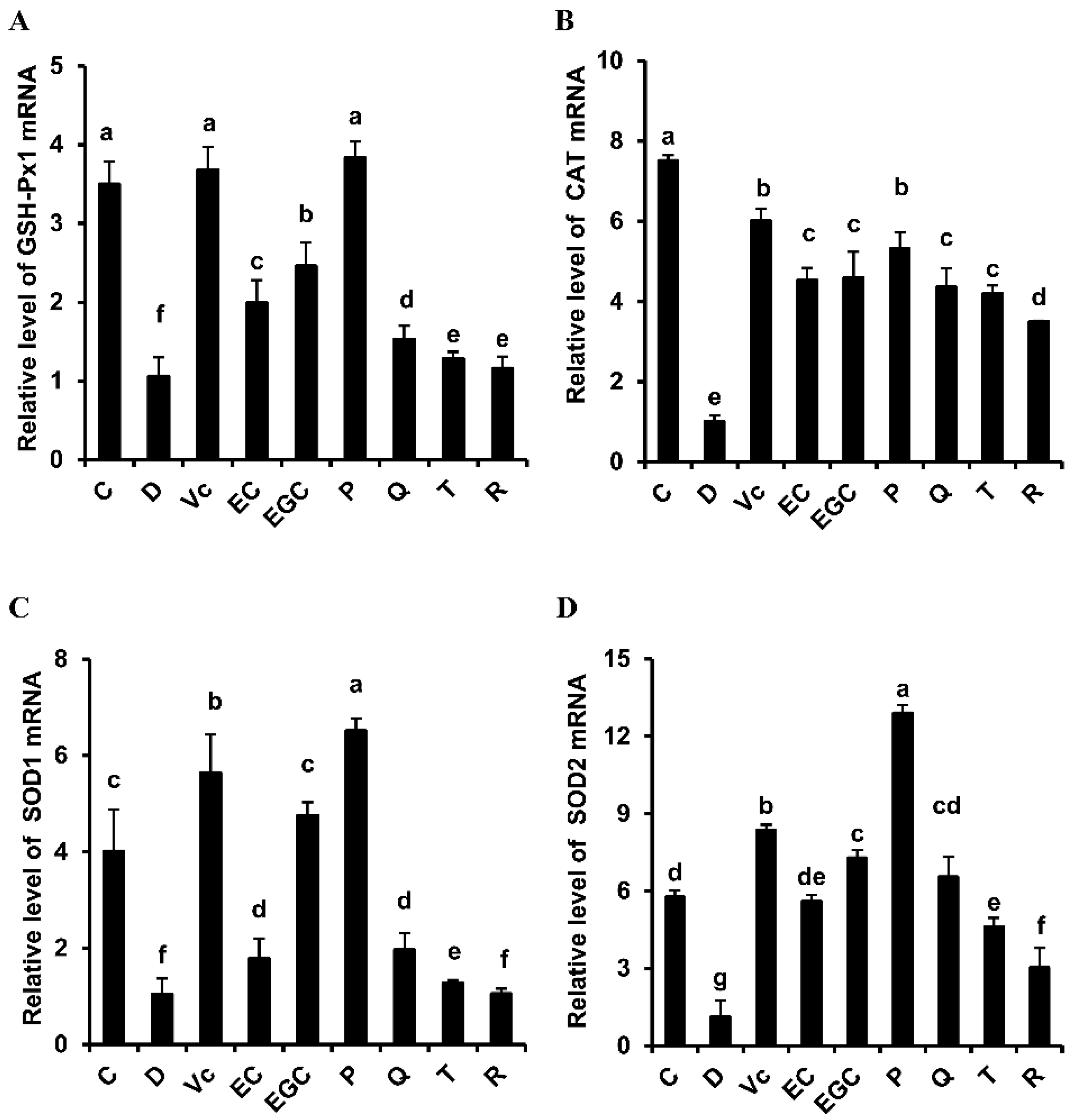
3.3. Effect of Six Flavonoids on Hepatic Oxidative Stress in d-Galactose-Treated Mice
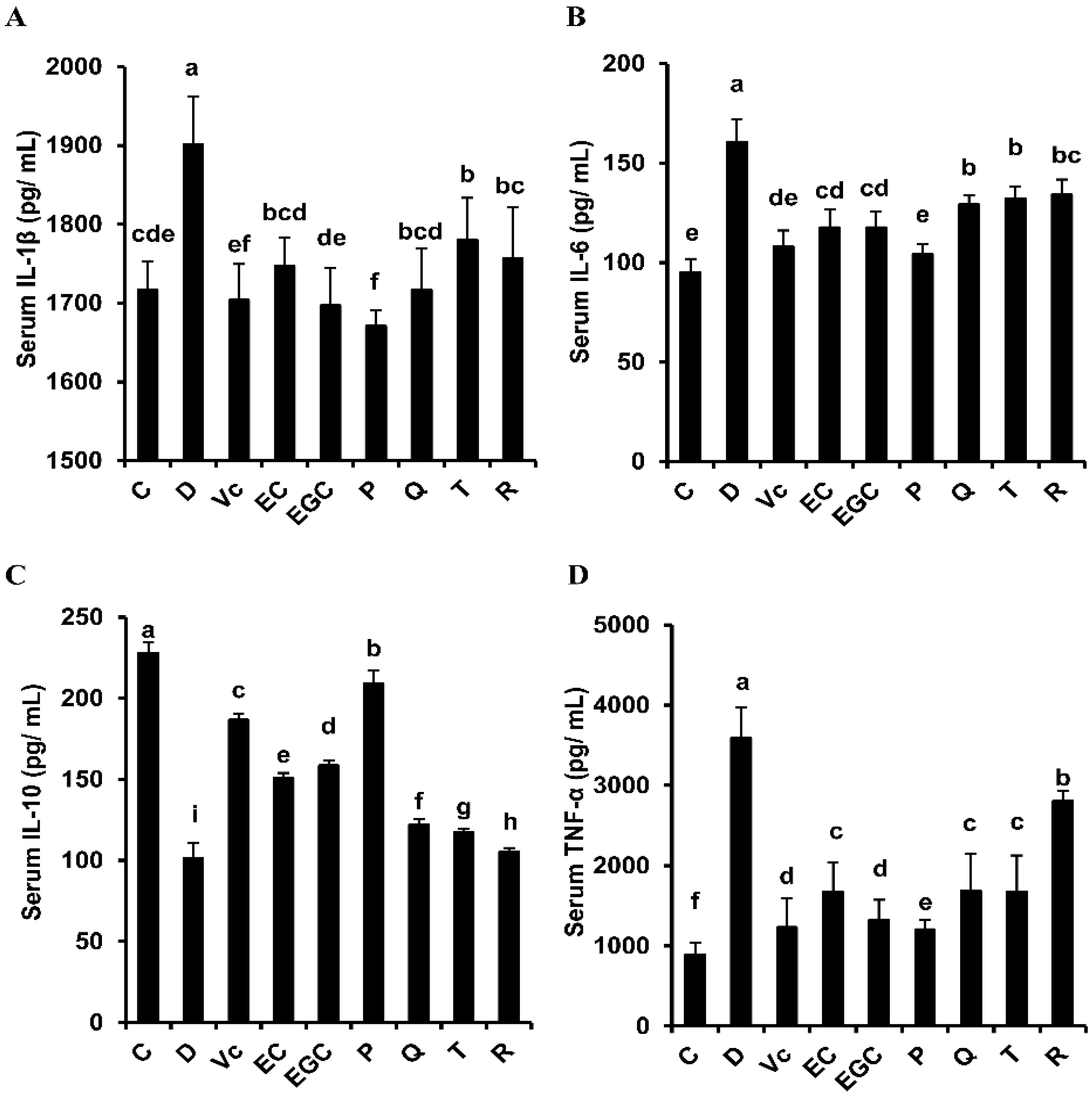
3.4. Effect of Six Flavonoids on Oxidative Stress-Related Inflammatory Response in D-Galactose-Treated Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ji, X.; Leak, R.K.; Chen, F.; Cao, G. Stem cell therapies in age-related neurodegenerative diseases and stroke. Ageing Res. Rev. 2017, 34, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Zhang, Y.; Qi, J.; Liu, R.; Zhang, H.; He, L. Ferulic acid inhibits H2O2-induced oxidative stress and inflammation in rat vascular smooth muscle cells via inhibition of the NADPH oxidase and NF-κB pathway. Int. Immunopharmacol. 2015, 28, 1018–1025. [Google Scholar] [CrossRef] [PubMed]
- Rani, V.; Deep, G.; Singh, R.K.; Palle, K.; Yadav, U.C.S. Oxidative stress and metabolic disorders: Pathogenesis and therapeutic strategies. Life Sci. 2016, 148, 183–193. [Google Scholar] [CrossRef]
- Luo, J.; Mills, K.; le Cessie, S.; Noordam, R.; van Heemst, D. Ageing, age-related diseases and oxidative stress: What to do next? Ageing Res. Rev. 2020, 57, 100982. [Google Scholar] [CrossRef]
- Viña, J.; Borrás, C.; Miquel, J. Theories of ageing. Iubmb Life 2007, 59, 249–254. [Google Scholar] [CrossRef]
- Zeng, L.; Lin, L.; Peng, Y.; Yuan, D.; Zhang, S.; Gong, Z.; Xiao, W. L-Theanine attenuates liver aging by inhibiting advanced glycation end products in d-galactose-induced rats and reversing an imbalance of oxidative stress and inflammation. Exp. Gerontol. 2020, 131, 110823. [Google Scholar] [CrossRef]
- Raman, S.T.; Ganeshan, A.K.P.G.; Chen, C.; Jin, C.; Li, S.; Chen, H.; Gui, Z. In vitro and in vivo antioxidant activity of flavonoid extracted from mulberry fruit (Morus alba L.). Pharmacogn. Mag. 2016, 12, 128–133. [Google Scholar]
- Wu, P.; Ma, G.; Li, N.; Deng, Q.; Yin, Y.; Huang, R. Investigation of in vitro and in vivo antioxidant activities of flavonoids rich extract from the berries of Rhodomyrtus tomentosa(Ait.) Hassk. Food Chem. 2015, 173, 194–202. [Google Scholar] [CrossRef]
- Wang, L.; Chen, Q.; Zhuang, S.; Wen, Y.; Cheng, W.; Zeng, Z.; Jiang, T.; Tang, C. Effect of Anoectochilus roxburghii flavonoids extract on H2O2-induced oxidative stress in LO2 cells and d-gal induced aging mice model. J. Ethnopharmacol. 2020, 254, 112670. [Google Scholar] [CrossRef]
- Wu, M.; Wang, C.; Mai, C.; Chen, J.; Lai, X.; He, L.; Huang, S.; Zhang, X. Flavonoids from Livistona chinensis fruit ameliorates LPS/D-GalN-induced acute liver injury by inhibiting oxidative stress and inflammation. J. Funct. Foods. 2019, 61, 103460. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, Y.; Nabi, X. Protective effect of Ziziphora clinopodioides flavonoids against H2O2-induced oxidative stress in HUVEC cells. Biomed. Pharmacother. 2019, 117, 109156. [Google Scholar] [CrossRef] [PubMed]
- Narayani, M. Bioethanol Production from Tea Fungal Biomass Grown on Tea Manufacture Waste. Master’s Thesis, National Institute of Technology, Rourkela, India, 2013. [Google Scholar]
- Heim, K.; Tagliaferro, A.; Bobilya, D. Flavonoid antioxidants: Chemistry, metabolism and structure-activity relationships. J. Nutr. Biochem. 2002, 13, 572–584. [Google Scholar] [CrossRef]
- Chen, Y.; Tang, S.; Chen, Y.; Zhang, R.; Zhou, M.; Wang, C.; Feng, N.; Wu, Q. Structure-activity relationship of procyanidins on advanced glycation end products formation and corresponding mechanisms. Food Chem. 2019, 272, 679–687. [Google Scholar] [CrossRef]
- Çelik, H.; Koşar, M. Inhibitory effects of dietary flavonoids on purified hepatic NADH-cytochrome b5 reductase: Structure-activity relationships. Chem. Biol. Interact. 2012, 197, 103–109. [Google Scholar] [CrossRef]
- Sarian, M.N.; Ahmed, Q.U.; So’ad, S.Z.M.; Alhassan, A.M.; Murugesu, S.; Perumal, V.; Mohamad, S.N.A.S.; Khatib, A.; Latip, J. Antioxidant and antidiabetic effects of flavonoids: A structure-activity relationship based study. BioMed Res. Int. 2017, 2017, 8386065. [Google Scholar] [CrossRef]
- Yang, H.; Xiao, L.; Yuan, Y.; Luo, X.; Jiang, M.; Ni, J.; Wang, N. Procyanidin B2 inhibits NLRP3 inflammasome activation in human vascular endothelial cells. Biochem. Pharmacol. 2014, 92, 599–606. [Google Scholar] [CrossRef]
- Yang, D.; Liu, S.; Chen, Y.; Hsu, S.; Chang, Y.; Lin, J. Three pathways assess anti-inflammatory response of epicatechin with lipopolysaccharide-mediated macrophage RAW264.7 Cells. J. Food Biochem. 2015, 39, 334–343. [Google Scholar] [CrossRef]
- Zhu, N.; Huang, T.; Yu, Y.; LaVoie, E.; Yang, C.; Ho, C. Identification of oxidation products of (-)-epigallocatechin gallate and (-)-epigallocatechin with H2O2. J. Agric. Food Chem. 2000, 48, 979–981. [Google Scholar] [CrossRef]
- Zu, Y.; Wu, W.; Zhao, X.; Li, Y.; Wang, W.; Zhong, C.; Zhang, Y.; Zhao, X. Enhancement of solubility, antioxidant ability and bioavailability of taxifolin nanoparticles by liquid antisolvent precipitation technique. Int. J. Pharmaceut. 2014, 471, 366–376. [Google Scholar] [CrossRef]
- Chang, J.; Lai, S.; Chen, W.; Hung, W.; Chow, J.; Hsiao, M.; Lee, W.; Chien, M. Quercetin suppresses the metastatic ability of lung cancer through inhibiting snail-dependent Akt activation and snail-independent ADAM9 expression pathways. BBA Mol. Cell Res. 2017, 1864, 1746–1758. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Ning, S.; Li, Y.; Yu, Y.; Shen, C.; Duan, G. Optimisation of infrared-assisted extraction of rutin from crude flos sophorae immaturus using response surface methodology and HPLC analysis. Phytochem. Anal. 2012, 23, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Wang, X.; Liu, H.; Lv, C.; Lu, J. Structural characterization and antioxidant activity of oligosaccharides from Panax ginseng C. A. Meyer. Int. J. Biol. Macromol. 2020, 150, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.P.; Kaur, A.; Singh, N.; Nim, L.; Shevkani, K.; Kaur, H.; Arora, D.S. In vitro antioxidant and antimicrobial properties of jambolan (Syzygium cumini) fruit polyphenols. LWT Food Sci. Technol. 2015, 65, 1025–1030. [Google Scholar] [CrossRef]
- Huang, R.; Zhang, Y.; Shen, S.; Zhi, Z.; Cheng, H.; Chen, S.; Ye, X. Antioxidant and pancreatic lipase inhibitory effects of flavonoids from different citrus peel extracts: An in vitro study. Food Chem. 2020, 326, 126785. [Google Scholar] [CrossRef]
- Floegel, A.; Kim, D.O.; Chung, S.J.; Koo, S.I.; Chun, O.K. Comparison of ABTS/DPPH assays to measure antioxidant capacity in popular antioxidant-rich US foods. J. Food Compos. Anal. 2011, 24, 1043–1048. [Google Scholar] [CrossRef]
- Ahmadi, S.M.; Farhoosh, R.; Sharif, A.; Rezaie, M. Structure-antioxidant activity relationships of luteolin and catechin. J. Food Sci. 2020, 85, 298–305. [Google Scholar] [CrossRef]
- Chaaban, H.; Ioannou, I.; Chebil, L.; Slimane, M.; Gérardin, C.; Paris, C.; Charbonnel, C.; Chekir, L.; Ghoul, M. Effect of heat processing on thermal stability and antioxidant activity of six flavonoids. J. Food Process Preserv. 2017, 41, e13203. [Google Scholar] [CrossRef]
- Buchner, N.; Krumbein, A.; Rohn, S.; Kroh, L.W. Effect of thermal processing on the flavonols rutin and quercetin. Rapid Commun. Mass Spectrom. 2010, 20, 3229–3235. [Google Scholar] [CrossRef]
- Ioannou, I.; Hafsa, I.; Hamdi, S.; Charbonnel, C.; Ghoul, M. Review of the effects of food processing and formulation on flavonol and anthocyanin behaviour. J. Food Eng. 2012, 111, 208–217. [Google Scholar] [CrossRef]
- Cai, Y.Z.; Sun, M.; Xing, J.; Luo, Q.; Corke, H. Structure-radical scavenging activity relationships of phenolic compounds from traditional Chinese medicinal plants. Life Sci. 2006, 78, 2872–2888. [Google Scholar] [CrossRef] [PubMed]
- Ajay, M.; Cheng, H.M.; Mohd, M.A.; Mustafa, M.R. A comparative study on antioxidant activity of flavonoids: Structure-activity relationships. Malays. J. Sci. 2005, 24, 187–190. [Google Scholar]
- Zhang, Y.; Wang, D.; Yang, L.; Zhou, D.; Zhang, J. Purification and characterization of flavonoids from the leaves of Zanthoxylum bungeanum and correlation between their structure and antioxidant activity. PLoS ONE 2014, 9, e105725. [Google Scholar] [CrossRef] [PubMed]
- Gregoris, E.; Stevanato, R. Correlations between polyphenolic composition and antioxidant activity of Venetian propolis. Food Chem. Toxicol. 2010, 48, 82. [Google Scholar] [CrossRef]
- Feng, J.; Zhang, X.; Li, Y.; Cui, Y.; Chen, Y.H. Pinus massoniana bark extract: Structure-activity relationship and biomedical potentials. Am. J. Chin. Med. 2016, 44, 1559–1577. [Google Scholar] [CrossRef]
- Tian, Y.; Wen, Z.; Lei, L.; Li, F.; Zhao, J.; Zhi, Q.; Li, F.; Yin, R.; Ming, J. Coreopsis tinctoria flowers extract ameliorates d-galactose induced aging in mice via regulation of Sirt1-Nrf2 signaling pathway. J. Funct. Foods. 2019, 60, 103464. [Google Scholar] [CrossRef]
- Govindan, S.; Johnson, E.E.R.; Christopher, J.; Shanmugam, J.; Thirumalairaj, V.; Gopalan, J. Antioxidant and anti-aging activities of polysaccharides from Calocybe indica var. APK2. Exp. Toxicol. Pathol. 2016, 68, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Vives-Bauza, C.; Starkov, A.; Garcia-Arumi, E. Measurements of the antioxidant enzyme activities of superoxide dismutase, catalase, and glutathione peroxidase. Method. Cell Biol. 2007, 80, 379–393. [Google Scholar]
- Zhang, Z.; Fan, S.; Zheng, Y.; Lu, J.; Wu, D.; Shan, Q.; Hu, B. Purple sweet potato color attenuates oxidative stress and inflammatory response induced by d-galactose in mouse liver. Food Chem. Toxicol. 2009, 47, 496–501. [Google Scholar] [CrossRef]
- Zheng, J.; Xie, Y.; Li, F.; Zhou, Y.; Qi, L.; Liu, L.; Chen, Z. Lactoferrin improves cognitive function and attenuates brain senescence in aged mice. J. Funct. Foods 2020, 65, 103736. [Google Scholar] [CrossRef]
- Li, J.; Nie, S.; Xie, M.; Huang, D.; Wang, Y.; Li, C. Chemical composition and antioxidant activities in immumosuppressed mice of polysaccharides isolated from Mosla chinensis Maxim cv jiangxiangru. Int. Immunopharmacol. 2013, 17, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Mei, X.; Zheng, Y.; Xu, D. Gastroprotective effect of taurine zinc solid dispersions against absolute ethanol-induced gastric lesions is mediated by enhancement of antioxidant activity and endogenous PGE2 production and attenuation of NO production. Eur. J. Pharmacol. 2014, 740, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Qu, Z.; Zhang, J.; Yang, H.; Huo, L.; Gao, J.; Chen, H.; Gao, W. Protective effect of tetrahydropalmatine against d-galactose induced memory impairment in rat. Physiol. Behav. 2016, 154, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Chen, J.; Zhang, Y.; Guan, M.; Li, X.; Zhou, Q.; Song, Q.; Qiu, J. Synthesis and assessment of phenylacrylamide derivatives as potential anti-oxidant and anti-inflammatory agents. Eur. J. Med. Chem. 2019, 180, 62–71. [Google Scholar] [CrossRef]
- Maurent, K.; Vanucci-Bacqué, C.; Baltas, M.; Nègre-Salvayre, A.; Augé, N.; Bedos-Belval, F. Synthesis and biological evaluation of diarylheptanoids as potential antioxidant and anti-inflammatory agents. Eur. J. Med. Chem. 2018, 144, 289–299. [Google Scholar] [CrossRef]
- Rutz, S.; Ouyang, W.; Crellin, N.K.; Valdez, P.A.; Hymowitz, S.G. Regulation and functions of the IL-10 family of cytokines in inflammation and disease. Annu. Rev. Immunol. 2011, 29, 71–109. [Google Scholar]
- Kiecolt-Glaser, J.K.; Preacher, K.J.; Maccallum, R.C.; Atkinson, C.; Glaser, R. Chronic stress and age-related increases in the proinflammatory cytokine IL-6. Proc. Natl. Acad. Sci. USA 2003, 100, 9090–9095. [Google Scholar] [CrossRef]
- Sun, L.; Zhao, Q.; Xiao, Y.; Liu, X.; Li, Y.; Zhang, J.; Pan, J.; Zhang, Z. Trehalose targets Nrf2 signal to alleviate d-galactose induced aging and improve behavioral ability. Biochem. Bioph. Res. Commun. 2020, 521, 113–119. [Google Scholar] [CrossRef]
- Mohamadi-Zarch, S.; Baluchnejadmojarad, T.; Nourabadi, D.; Khanizadeh, A.M.; Roghani, M. Protective effect of diosgenin on LPS/D-Gal-induced acute liver failure in C57BL/6 mice. Microb. Pathog. 2020, 146, 104243. [Google Scholar] [CrossRef]
- Chen, G.; Fan, M.; Wu, J.; Li, N.; Guo, M. Antioxidant and anti-inflammatory properties of flavonoids from lotus plumule. Food Chem. 2019, 277, 706–712. [Google Scholar] [CrossRef]
- Spagnuolo, C.; Moccia, S.; Russo, G.L. Anti-inflammatory effects of flavonoids in neurodegenerative disorders. Eur. J. Med. Chem. 2018, 153, 105–115. [Google Scholar] [CrossRef] [PubMed]
- GutiErrez-Grijalva, E.P.; Ambriz-Pere, D.L.; Leyva-Lopez, N.; Castillo-Lopez, R.L.; Heiedia, J.B. Review: Dietary phenolic compounds, health benefits and bioaccessibility. Arch. Latinoam. Nutr. 2016, 66, 87–100. [Google Scholar] [PubMed]
- Peng, Y.R.; Meng, Q.L.; Zhou, J.; Chen, B.; Xi, J.J.; Long, P.P.; Zhang, L.; Hou, R.Y. Nanoemulsion delivery system of tea polyphenols enhanced the bioavailability of catechins in rats. Food Chem. 2018, 242, 527–532. [Google Scholar] [CrossRef]
- Penalva, R.; Esparza, I.; Morales-Gracia, J.; Gonzalez-Navarro, C.J.; Larraneta, E.; Irache, J.M. Casein nanoparticles in combination with 2-hydroxypropyl-beta-cyclodextrin improves the oral bioavailability of quercetin. Int. J. Pharmaceut. 2019, 570, 118652. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.J.; Wang, Z.B.; Mi, Y.Y.; Gao, M.J.; Lv, J.N.; Meng, Y.H.; Yang, B.Y.; Kuang, H.X. UHPLC-MS/MS determination, pharmacokinetic, and bioavailability study of taxifolin in rat plasma after oral administration of its nanodispersion. Molecules 2016, 21, 494. [Google Scholar] [CrossRef] [PubMed]
- Murota, K.; Nakamura, Y.; Uehara, M. Flavonoid metabolism: The interaction of metabolites and gut microbiota. Biosci. Biotechnol. Biochem. 2018, 82, 600–610. [Google Scholar] [CrossRef]
- Kim, Y.; Lee, Y.; Choe, J.; Lee, H.; Kim, Y.; Jeoung, D. CD44-epidermal growth factor receptor interaction mediates hyaluronic acid-promoted cell motility by activating protein kinase C signaling involving Akt, Rac1, Phox, reactive oxygen species, focal adhesion kinase, and MMP-2. J. Biol. Chem. 2008, 283, 22513–22528. [Google Scholar] [CrossRef]
- Vladimirova, O.; Lu, F.; Shawver, L.; Kalman, B. The activation of protein kinase C induces higher production of reactive oxygen species by mononuclear cells in patients with multiple sclerosis than in controls. Inflamm. Res. 1999, 48, 412–416. [Google Scholar] [CrossRef]
- Ferriola, P.C.; Cody, V.; Middleton, E. Protein kinase C inhibition by plant flavonoids. Kinetic mechanisms and structure-activity relationships. Biochem. Pharmacol. 1989, 38, 1617–1624. [Google Scholar] [CrossRef]
- Lotito, S.B.; Frei, B. Relevance of apple polyphenols as antioxidants in human plasma: Contrasting in vitro and in vivo effects. Free. Radic. Biol. Med. 2004, 36, 201–211. [Google Scholar]
- Dong, D.; Quan, E.; Yuan, X.; Xie, Q.; Li, Z.; Baojian, W. Sodium oleate-based nanoemulsion enhances oral absorption of chrysin through inhibition of UGT-mediated metabolism. Mol. Pharmaceut. 2017, 14, 2864–2874. [Google Scholar] [CrossRef] [PubMed]
- Thilakarathna, S.H.; Rupasinghe, H.P.V. Flavonoid bioavailability and attempts for bioavailability enhancement. Nutrients 2013, 5, 3367–3387. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
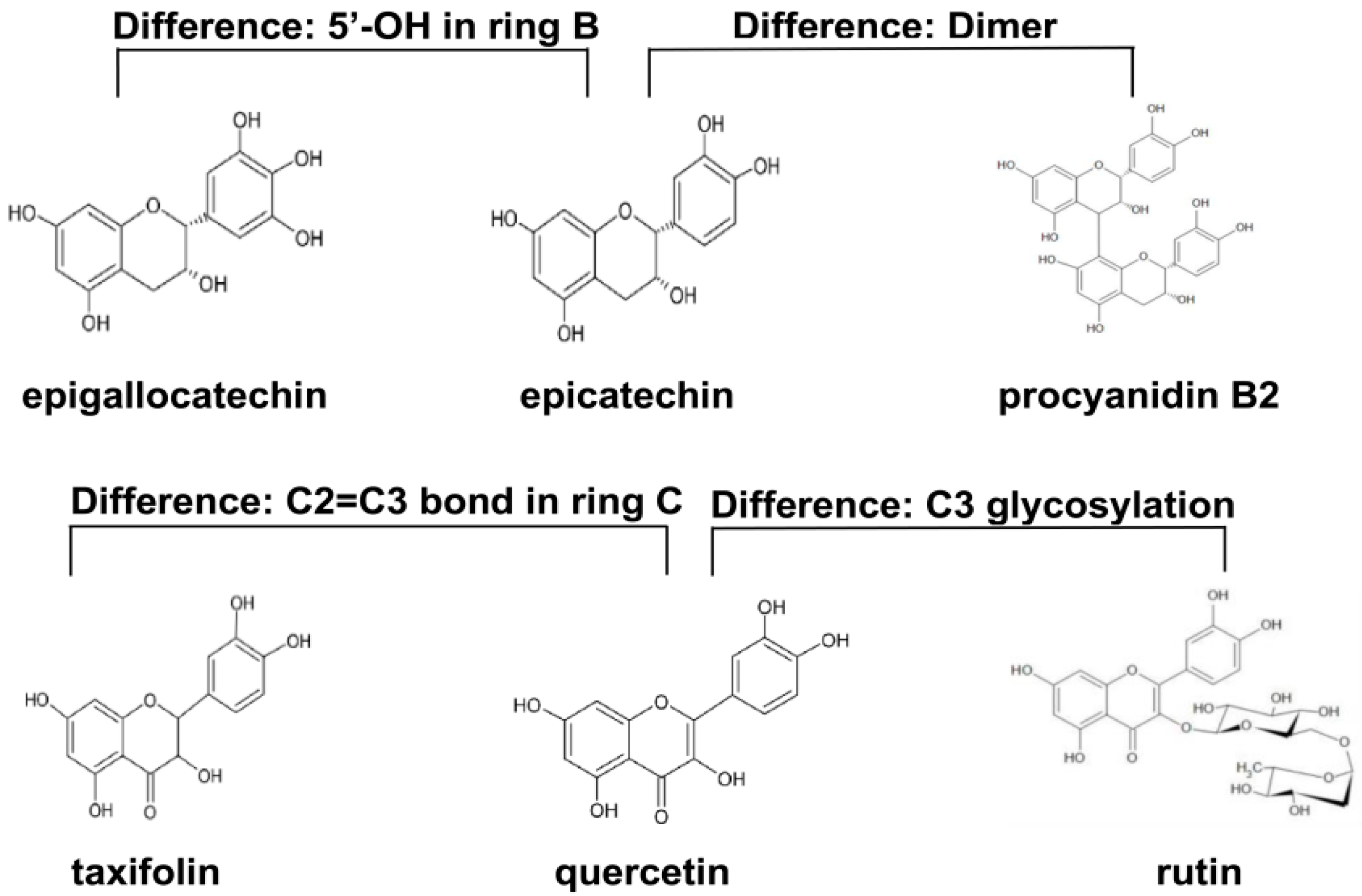
| Flavonoid | The Antioxidant Activities |
|---|---|
| Procyanidin B2 | Procyanidin B2, a widely distributed dimer in natural procyanidins, protects colonic cells against oxidative stress-induced injury [18]. |
| Epicatechin | Epicatechin can exert anti-inflammatory and antioxidant effects, and it can pass through the blood brain barrier to provide neuroprotection [19]. |
| Epigallocatechin | Two most important antioxidants in tea are epigallocatechin gallate and epigallocatechin, and the A ring of epigallocatechin has been demonstrated to be an antioxidant site [20]. |
| Taxifolin | Taxifolin is a free radical scavenger, and its antioxidant capacity is superior to ordinary flavonoids [21]. |
| Quercetin | Quercetin, a ubiquitously distributed flavonoid in plants, has a potent free radical scavenging capacity [22]. |
| Rutin | The antioxidant activity is considered as one of the important pharmacological effects of rutin [23]. |
| Primer Name | Accession Number | Forward Primer (5′ to 3′) | Reverse Primer (5′ to 3′) |
|---|---|---|---|
| GAPDH | NM_001289726.1 | AGGTCGGTGTGAACGGATTTG | GGGGTCGTTGATGGCAACA |
| SOD1 | NM_011434.2 | AACCAGTTGTGTTGTCAGGAC | CCACCATGTTTCTTAGAGTGAGG |
| SOD2 | NM_013671.3 | CAGACCTGCCTTACGACTATGG | CTCGGTGGCGTTGAGATTGTT |
| CAT | NM_009804.2 | GGAGGCGGGAACCCAATAG | GTGTGCCATCTCGTCAGTGAA |
| GSH-Px1 | NM_008160.6 | CCACCGTGTATGCCTTCTCC | AGAGAGACGCGACATTCTCAAT |
| Index | Dimer | 5′-OH in Ring B | C2=C3 Bond in Ring C | C3 Glycosylation |
|---|---|---|---|---|
| DPPH• scavenging efficiency | P (+) > EC (-) | EGC (+) > EC (-) | Q (+) > T (-) | Q (-) > R (+) |
| ABTS+ scavenging efficiency | P (+) > EC (-) | EGC (+) > EC (-) | Q (+) > T (-) | Q (-) > R (+) |
| Serum SOD | P (+) > EC (-) | EGC (+) > EC (-) | Q (+) ≈ T (-) | Q (-) > R (+) |
| Serum T-AOC | P (+) > EC (-) | EGC (+) ≈ EC (-) | Q (+) ≈ T (-) | Q (-) ≈ R (+) |
| Serum CAT | P (+) > EC (-) | EGC (+) ≈ EC (-) | Q (+) > T (-) | Q (-) > R (+) |
| Serum GSH-Px | P (+) > EC (-) | EGC (+) > EC (-) | Q (+) ≈ T (-) | Q (-) ≈ R (+) |
| Serum NO | P (+) < EC (-) | EGC (+) < EC (-) | Q (+) ≈ T (-) | Q (-) < R (+) |
| Serum MDA | P (+) < EC (-) | EGC (+) ≈ EC (-) | Q (+) < T (-) | Q (-) < R (+) |
| Hepatic GSH-Px1 mRNA | P (+) > EC (-) | EGC (+) > EC (-) | Q (+) > T (-) | Q (-) > R (+) |
| Hepatic CAT mRNA | P (+) > EC (-) | EGC (+) ≈ EC (-) | Q (+) ≈ T (-) | Q (-) > R (+) |
| Hepatic SOD1 mRNA | P (+) > EC (-) | EGC (+) > EC (-) | Q (+) > T (-) | Q (-) > R (+) |
| Hepatic SOD2 mRNA | P (+) > EC (-) | EGC (+) > EC (-) | Q (+) > T (-) | Q (-) > R (+) |
| Serum IL-10 | P (+) > EC (-) | EGC (+) > EC (-) | Q (+) > T (-) | Q (-) > R (+) |
| Serum IL-1β | P (+) < EC (-) | EGC (+) ≈ EC (-) | Q (+) ≈ T (-) | Q (-) ≈ R (+) |
| Serum IL-6 | P (+) < EC (-) | EGC (+) ≈ EC (-) | Q (+) ≈ T (-) | Q (-) ≈ R (+) |
| Serum TNF-α | P (+) < EC (-) | EGC (+) < EC (-) | Q (+) ≈ T (-) | Q (-) < R (+) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, Y.; Song, J.; Zhang, M.; Wang, H.; Zhang, Y.; Suo, H. Comparison of In Vitro and In Vivo Antioxidant Activities of Six Flavonoids with Similar Structures. Antioxidants 2020, 9, 732. https://doi.org/10.3390/antiox9080732
Zeng Y, Song J, Zhang M, Wang H, Zhang Y, Suo H. Comparison of In Vitro and In Vivo Antioxidant Activities of Six Flavonoids with Similar Structures. Antioxidants. 2020; 9(8):732. https://doi.org/10.3390/antiox9080732
Chicago/Turabian StyleZeng, Yixiu, Jiajia Song, Meimei Zhang, Hongwei Wang, Yu Zhang, and Huayi Suo. 2020. "Comparison of In Vitro and In Vivo Antioxidant Activities of Six Flavonoids with Similar Structures" Antioxidants 9, no. 8: 732. https://doi.org/10.3390/antiox9080732
APA StyleZeng, Y., Song, J., Zhang, M., Wang, H., Zhang, Y., & Suo, H. (2020). Comparison of In Vitro and In Vivo Antioxidant Activities of Six Flavonoids with Similar Structures. Antioxidants, 9(8), 732. https://doi.org/10.3390/antiox9080732