Comparative Studies on Different Citrus Cultivars: A Revaluation of Waste Mandarin Components


 ,
, 

Abstract
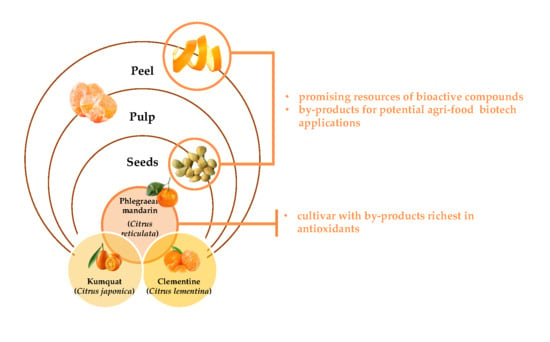
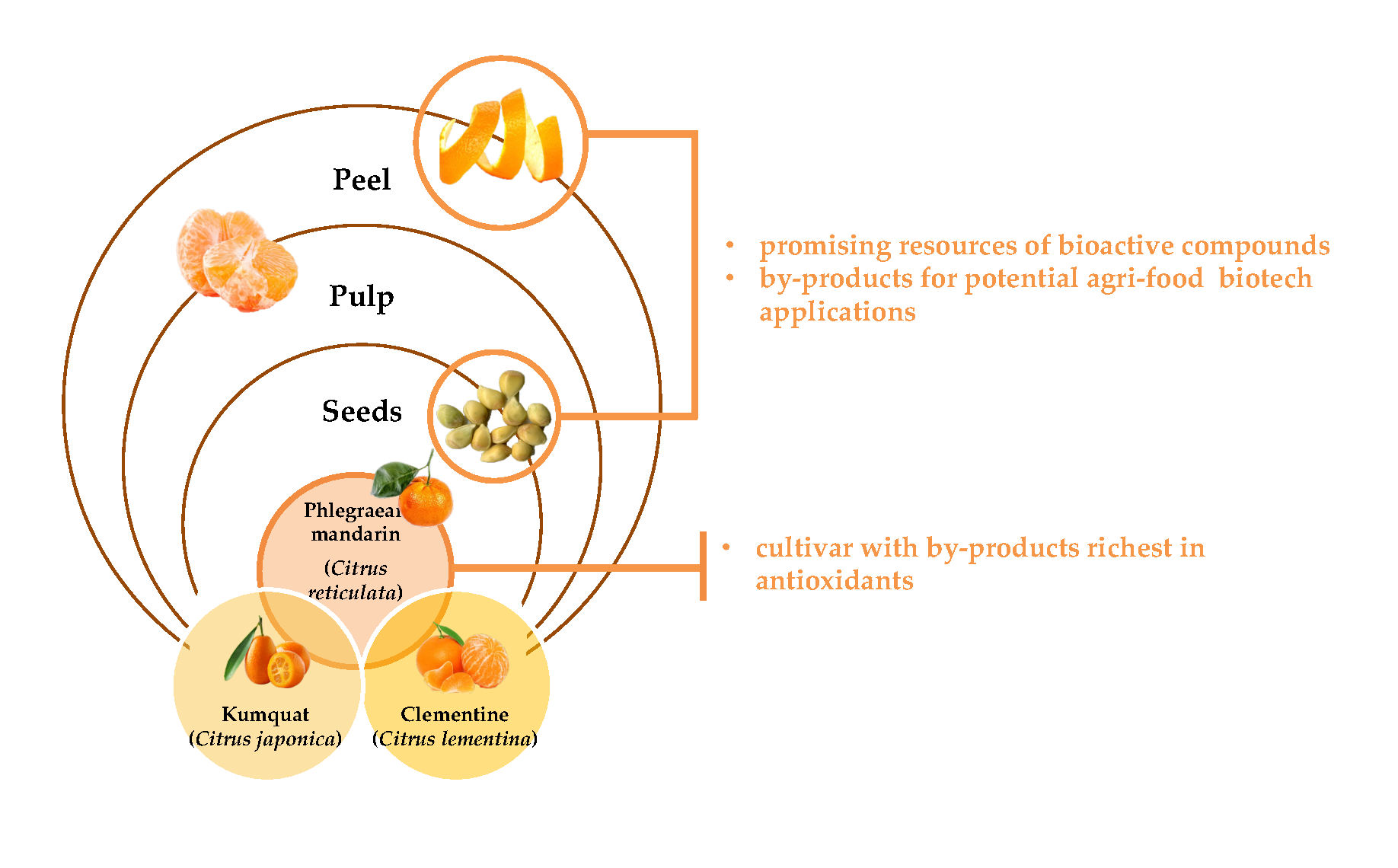
1. Introduction
2. Materials and Methods
2.1. Plant Material and Sample Preparation
2.2. Photosynthetic Pigments Content Determination
2.3. Total Polyphenol Content
2.4. Determination of Antioxidant Capacity and Ascorbic Acid
2.5. Statistical Analysis
3. Results
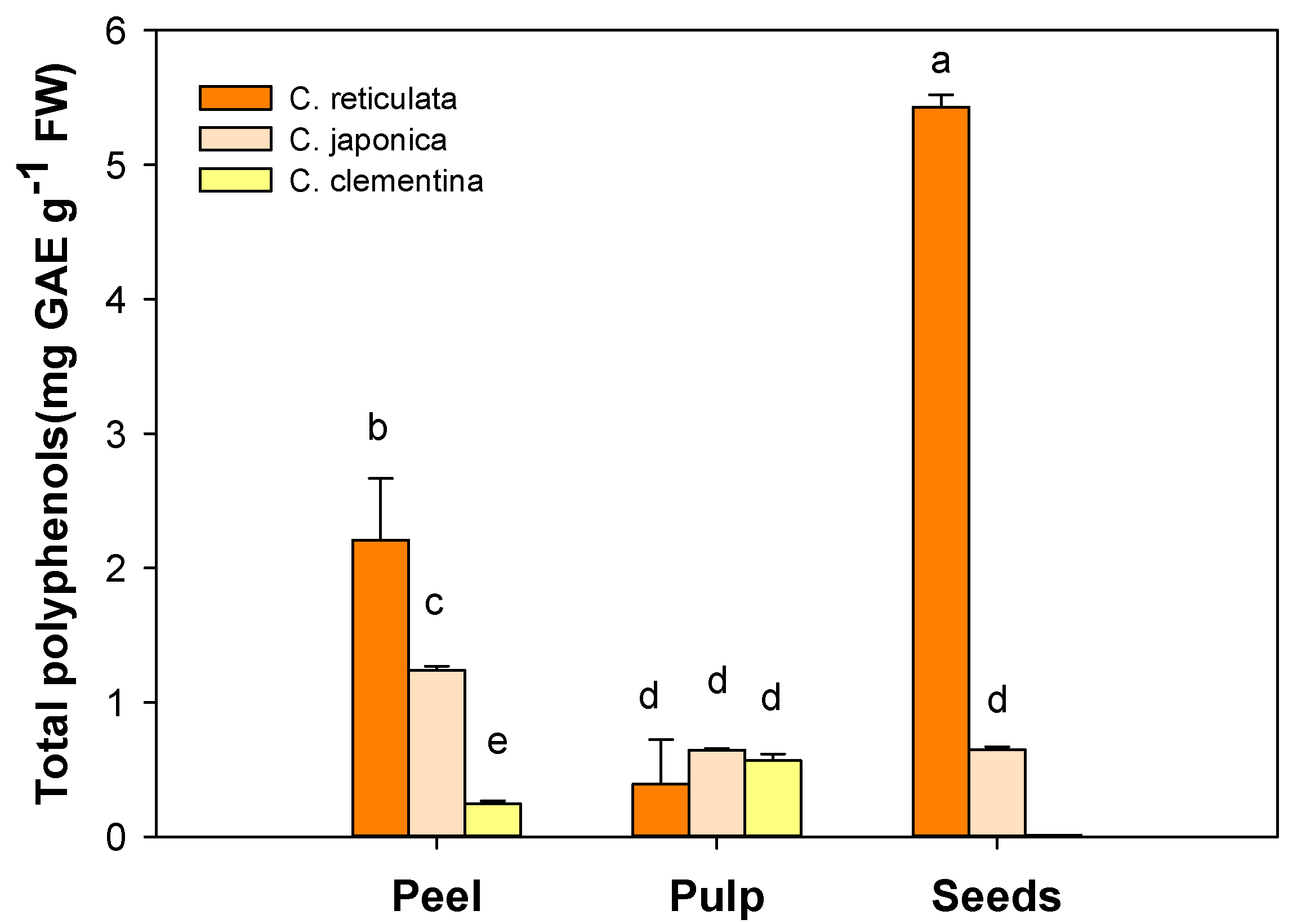
3.1. Total Polyphenol Content
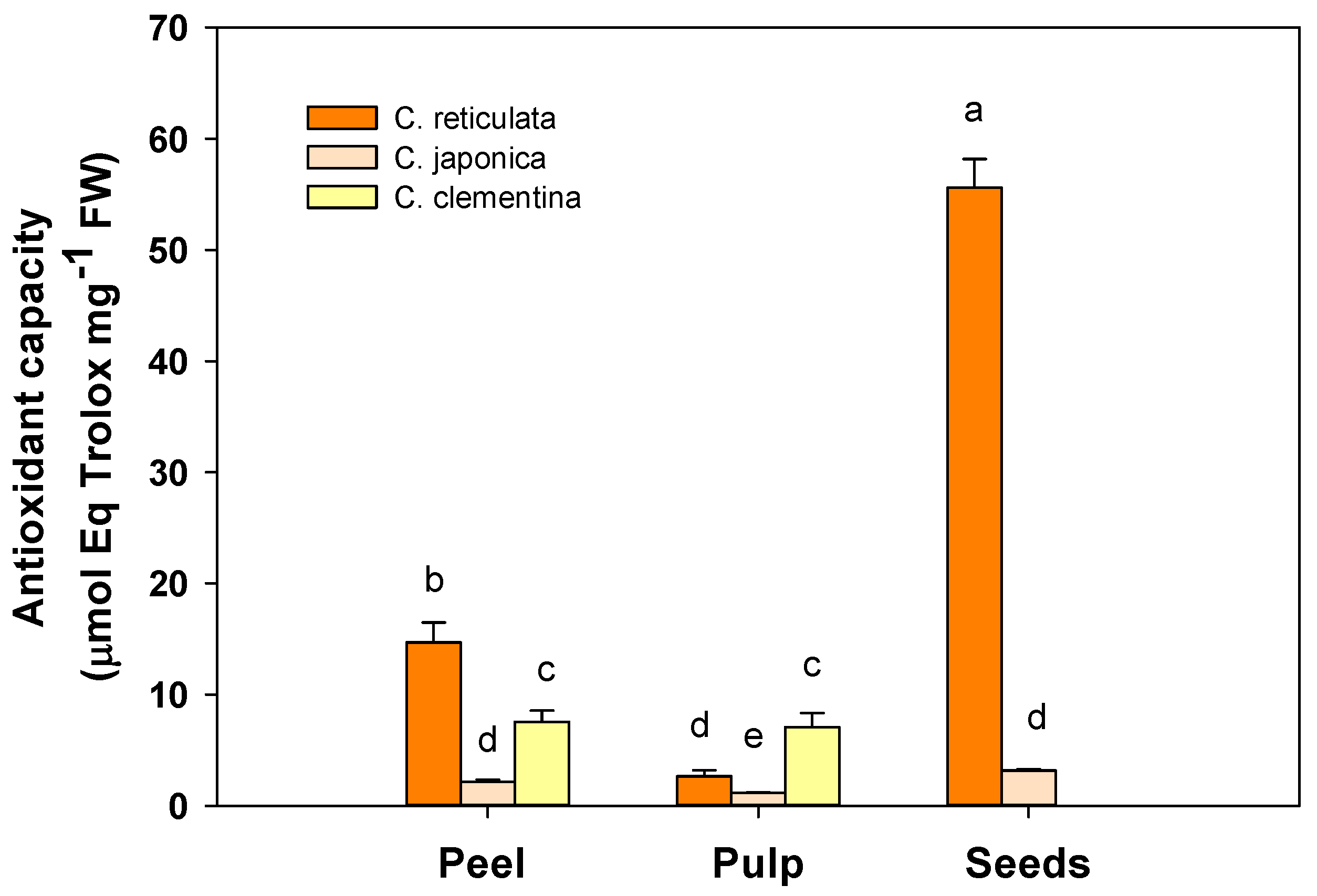
3.2. Total Soluble Antioxidant Capacity
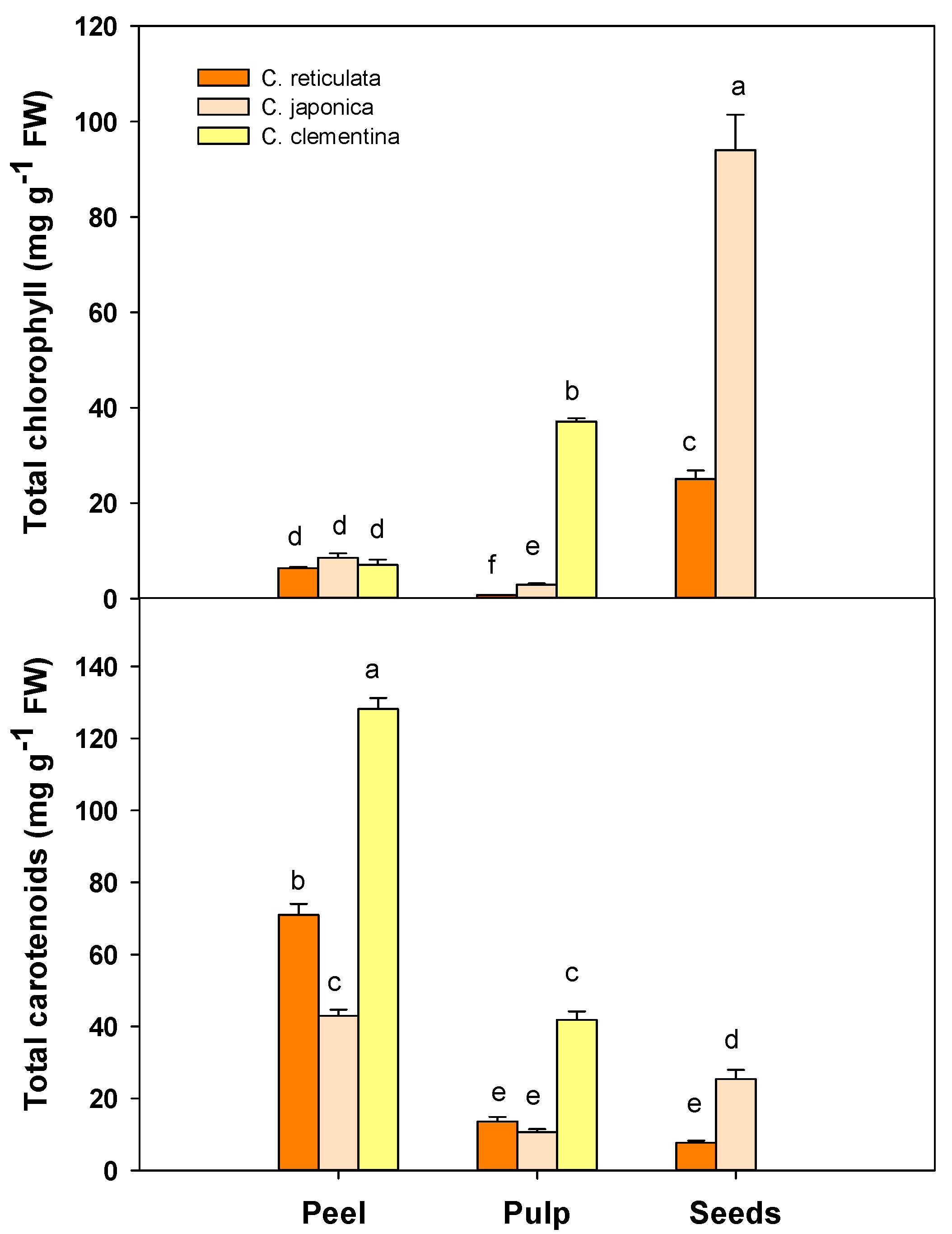
3.3. Total Chlorophyll and Carotenoid Content
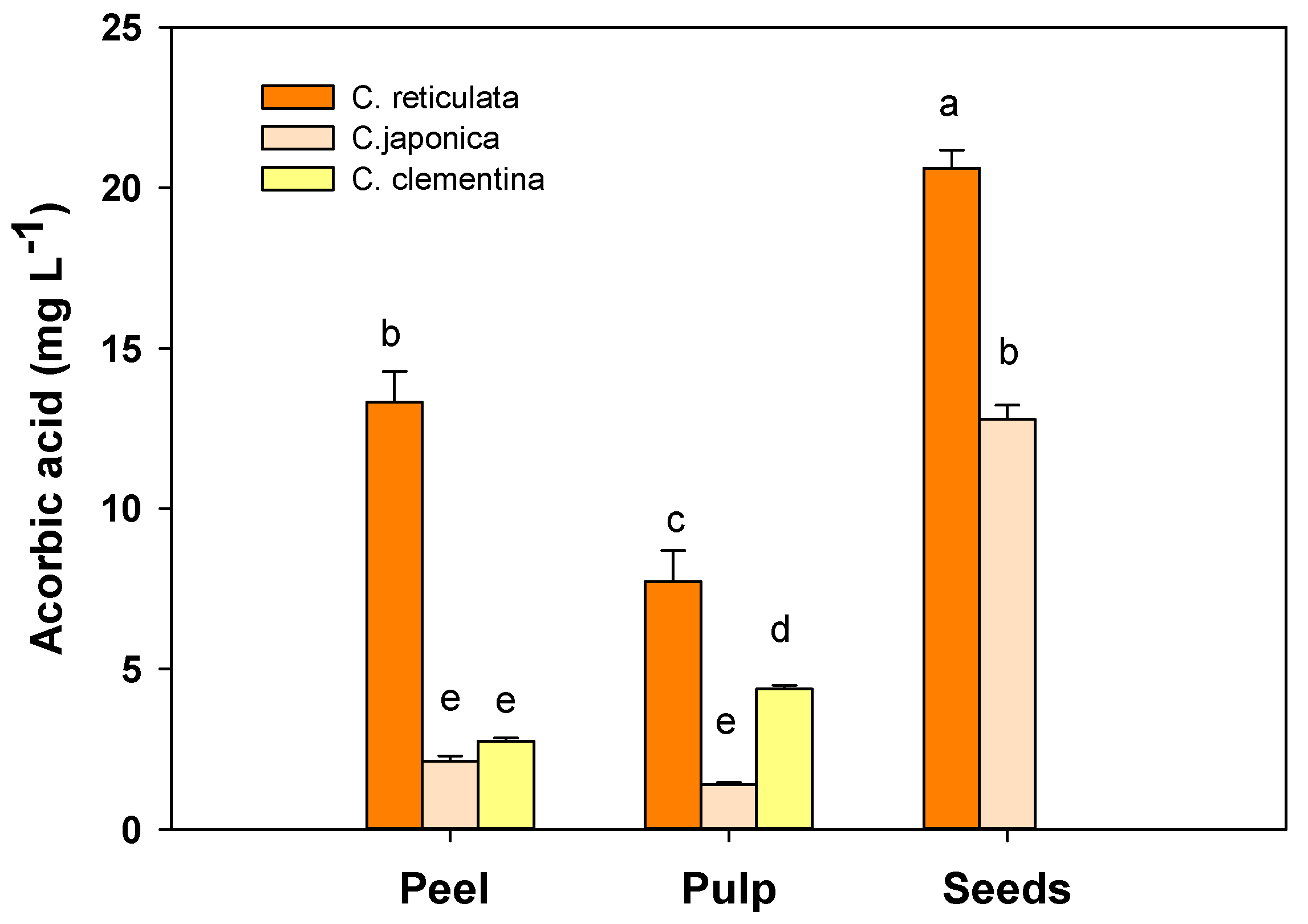
3.4. Ascorbic Acid
3.5. Relationship among Bioactive Compounds in the Seeds
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- De Moraes Barros, H.R.; Aparecida, T.; de Castro Ferreira, P.; Genovese, M.I. Antioxidant capacity and mineral content of pulp and peel from commercial cultivars of citrus from Brazil. Food Chem. 2012, 134, 1892–1898. [Google Scholar] [CrossRef] [PubMed]
- Boeing, H.; Bechthold, A.; Bub, A.; Ellinger, S.; Haller, D.; Kroke, A.; Leschik-Bonnet, E.; Muller, M.J.; Oberritter, H.; Schulze, M.; et al. Critical review: Vegetables and fruit in the prevention of chronic diseases. Eur. J. Nutr. 2012, 51, 637–663. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.D.; Ou, B.; Prior, R.L. The chemistry behind antioxidant capacity assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef] [PubMed]
- Patil, B.S.; Jayaprakasha, G.K.; Murthy, K.N.C.; Vikram, A. Bioactive compounds: Historical perspectives, opportunities, and challenges. J. Agric. Food Chem. 2009, 57, 8142–8160. [Google Scholar] [CrossRef]
- Masuda, T.; Inaba, Y.; Takeda, Y. Antioxidant mechanism of carnosic acid: Structural identification of two oxidation products. J. Agric. Food Chem. 2001, 49, 5560–5565. [Google Scholar] [CrossRef]
- Arena, E.; Fallico, B.; Maccarone, E. Evaluation of antioxidant capacity of blood orange juices as influenced by constituents, concentration process and storage. Food Chem. 2001, 74, 423–427. [Google Scholar] [CrossRef]
- Saito, S.; Okamoto, Y.; Kawabata, J. Effects of alcoholic solvents on antiradical abilities of protocatechuic acid and its alkyl esters. J. Biosci. Biotechnol. Biochem. 2004, 68, 1221–1227. [Google Scholar] [CrossRef]
- Al-Juhaimi, F.Y.; Ghafoor, K. Bioactive compounds, antioxidant and physico- chemical properties of juice from lemon, mandarin and orange fruits cultivated in Saudi Arabia. Pak. J. Bot. 2013, 45, 1193–1196. [Google Scholar]
- Ebrahimzadeh, M.A.; Hosseinimehr, S.J.; Gayekhloo, M.R. Measuring and comparison of vitamin C content in citrus fruits: Introduction of native variety. Indian J. Chem. B. 2004, 1, 650–652. [Google Scholar]
- Fernandez-Lopez, J.; Zhi, N.; Aleson-Carbonell, L.; Perez-Alvarez, J.A.; Kuri, V. Antioxidant and antibacterial activities of natural extracts: Application in beef meatballs. Meat Sci. 2005, 69, 371–380. [Google Scholar] [CrossRef]
- Jayaprakasha, G.K.; Patil, B.S. In vitro evaluation of the antioxidant activities in fruit extracts from citron and blood orange. Food Chem. 2007, 101, 410–418. [Google Scholar] [CrossRef]
- Barros, L.; Dueñas, M.; Dias, M.I.; Sousa, M.J.; Santos-Buelga, C.; Ferreira, I.C.F.R. Phenolic profiles of in vivo and in vitro grown Coriandrum sativum L. Food Chem. 2012, 132, 841–848. [Google Scholar] [CrossRef]
- Gorinstein, S.; Belloso, O.M.; Park, Y.; Haruenkit, R.; Lojek, A.; Ciz, M.; Caspi, A.; Libman, I.; Trakhtenberg, S. Comparison of some biochemical characteristics of different citrus fruits. Food Chem. 2001, 74, 309–315. [Google Scholar] [CrossRef]
- Topuz, A.; Topakci, M.; Canakci, M.; Akinci, I.; Ozdemir, F. Physical and nutritional properties of four orange varieties. J. Food Eng. 2005, 66, 519–523. [Google Scholar] [CrossRef]
- Goldemberg, L.; Yaniv, Y.; Porat, R.; Carmi, N. Mandarin fruit quality: A review. J. Sci. Food Agric. 2018, 98, 18–26. [Google Scholar] [CrossRef]
- Putnik, P.; Barba, F.J.; Lorenzo, J.M.; Gabric, D.; Shpigelman, A.; Cravotto, G.; Kovačević, D.B. An integrated approach to mandarin processing: Food safety and nutritional quality, consumer preference, and nutrient bioaccessibility. Compr. Rev. Food Sci. Saf. 2017, 16, 1345–1358. [Google Scholar] [CrossRef]
- BP Statistical Review of World Energy. 2012. Available online: http://www.bp.com/assets/bp_internet/globalbp/globalbp_uk_english/reports_and_publications/statistical_energy_review_2011/STAGING/local_assets/pdf/statistical_review_of_world_energy_full_report_2012.pdf (accessed on 5 September 2012).
- Sharma, K.; Mahato, N.; Cho, M.H.; Lee, Y.R. Converting citrus wastes into value-added products: Economic and environmentally friendly approaches. Nutrition 2017, 34, 29–46. [Google Scholar] [CrossRef]
- Wikandari, R.; Nguyen, H.; Millati, R.; Niklasson, C.; Taherzadeh, M.J. Improvement of biogas production from orange peel waste by leaching of limonene. BioMed Res. Int. 2015, 2015, 494182. [Google Scholar] [CrossRef]
- Ledesma-Escobar, C.A.; de Castro, M.D.L. Towards a comprehensive exploitation of citrus. Trends Food Sci. Technol. 2014, 39, 63–75. [Google Scholar] [CrossRef]
- Lin, C.S.K.; Pfaltzgraff, L.A.; Herrero-Davila, L.; Mubofu, E.B.; Abderrahim, S.; Clark, J.H.; Koutinas, A.A.; Kopsahelis, N.; Stamatelatou, K.; Dickson, F.; et al. Food waste as a valuable resource for the production of chemicals, materials and fuels. Current situation and global perspective. Energy Environ. Sci. 2013, 6, 426–464. [Google Scholar] [CrossRef]
- Bampidis, V.A.; Robinson, P.H. Citrus by-products as ruminant feeds: A review. Anim. Feed Sci. Technol. 2006, 128, 175–217. [Google Scholar] [CrossRef]
- Calabrò, P.S.; Pontoni, L.; Porqueddu, I.; Greco, R.; Pirozzi, F.; Malpei, F. Effect of the concentration of essential oil on orange peel waste biomethanization: Preliminary batch results. Waste Manag. 2016, 48, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Deng, G.F.; Shen, C.; Xu, X.R.; Kuang, R.D.; Guo, Y.J.; Zeng, L.S.; Gao, L.-L.; Lin, X.; Xie, J.-F.; Xia, E.; et al. Potential of fruit wastes as natural resources of bioactive compounds. Int. J. Mol. Sci. 2012, 13, 8308–8323. [Google Scholar] [CrossRef] [PubMed]
- Manthey, J.A.; Grohmann, K. Phenols in citrus peel by products: Concentrations of hydroxycinnamates and polymethoxylated flavones in citrus peel molasses. J. Agric. Food Chem. 2001, 49, 3268–3273. [Google Scholar] [CrossRef]
- Elkhatim, K.A.S.; Elagib, R.A.A.; Hassan, A.B. Content of phenolic compounds and vitamin C and antioxidant activity in wasted parts of Sudanese citrus fruits. Food Sci. Nutr. 2018, 6, 1214–1219. [Google Scholar] [CrossRef] [PubMed]
- Bocco, A.; Cuvelier, M.; Richard, H.; Berset, C. Antioxidant activity and phenolic composition of citrus peel and seed extracts. J. Agric. Food Chem. 1998, 46, 2123–2129. [Google Scholar] [CrossRef]
- Fattahi, J.; Hamidoghli, Y.; Ghazvini, R.F.; Ghasemnejad, M.; Bakhshi, D. Assessment of fruit quality and antioxidant activity of three citrus species during ripening. South-West. J. Hortic. Biol. Environ. 2011, 2, 113–128. [Google Scholar]
- Moulehi, I.; Bourgou, S.; Ourghemmi, I.; Tounsi, M.S. Variety and ripening impact on phenolic composition and antioxidant activity of mandarin (Citrus reticulate Blanco) and bitter orange (Citrus aurantium L.) seeds extracts. Ind. Crop. Prod. 2012, 39, 74–80. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Porzio, L.; Buia, M.C.; Lorenti, M.; Vitale, E.; Amitrano, C.; Arena, C. Ecophysiological response of Jania rubens (Corallinaceae) to ocean acidification. Rend. Lincei-Sci. Fis. 2018, 29, 543–546. [Google Scholar] [CrossRef]
- George, B.; Kaur, C.; Khurdiya, D.S.; Kapoor, H.C. Antioxidants in tomato (Lycopersium esculentum) as a function of genotype. Food Chem. 2004, 84, 45–51. [Google Scholar] [CrossRef]
- Arena, C.; Vitale, L.; Bianchi, A.R.; Mistretta, C.; Vitale, E.; Parisi, C.; Guerriero, G.; Magliulo, V.; De Maio, A. The ageing process affects the antioxidant defences and the poly (ADPribosyl)ation activity in Cistus incanus L. leaves. Antioxidants 2019, 8, 528. [Google Scholar] [CrossRef] [PubMed]
- Agócs, A.; Nagy, V.; Szabó, Z.; Márk, L.; Ohmacht, R.; Deli, J. Comparative study on the carotenoid composition of the peel and the pulp of different citrus species. Innov. Food Sci. Emerg. Technol. 2007, 8, 390–394. [Google Scholar] [CrossRef]
- Kato, M.; Ikoma, Y.; Matsumoto, H.; Sugiura, M.; Hyodo, H.; Yano, M. Accumulation of carotenoids and expression of carotenoid biosynthetic genes during maturation in citrus fruit. Plant Physiol. 2004, 134, 824–837. [Google Scholar] [CrossRef] [PubMed]
- Fanciullino, A.L.; Dhuique-Mayer, C.; Luro, F.; Casanova, J.; Morillon, R.; Ollitrault, P. Carotenoid diversity in cultivated Citrus is highly influenced by genetic factors. J. Agric. Food Chem. 2006, 54, 4397–4406. [Google Scholar] [CrossRef]
- Matsumoto, H.; Ikoma, Y.; Kato, M.; Kuniga, T.; Nakajima, N.; Yoshida, T. Quantification of carotenoids in citrus fruit by LC-MS and comparison of patterns of seasonal changes for carotenoids among citrus varieties. J. Agric. Food Chem. 2007, 55, 2356–2368. [Google Scholar] [CrossRef]
- Conesa, A.; Manera, F.C.; Brotons, J.M.; Fernandez-Zapata, J.C.; Simón, I.; Simón-Grao, S.; Alfosea-Simón, M.; Nicolás, J.J.M.; Valverde, J.M.; Garcìa-Sanchez, F. Changes in the content of chlorophylls and carotenoids in the rind of Fino 49 lemons during maturation and their relationship with parameters from the CIELAB color space. Sci. Hortic. 2019, 243, 253–260. [Google Scholar] [CrossRef]
- Casodoro, G.; Rascio, N. Cotyledonal chloroplasts in the hypogeal seeds of clementine. Planta 1987, 170, 300–307. [Google Scholar] [CrossRef]
- Marcelis, L.F.M.; Hofman-Eijer, L.R.B. The contribution of fruit photosynthesis to the carbon requirement of cucumber fruits as affected by irradiance, temperature and ontogeny. Physiol. Plant 1995, 93, 476–483. [Google Scholar] [CrossRef]
- Howitt, C.A.; Pogson, B.J. Carotenoid accumulation and function in seeds and non-green tissues. Plant Cell Environ. 2006, 29, 435–445. [Google Scholar] [CrossRef]
- Pinzino, C.; Capocchi, A.; Galleschi, L.; Saviozzi, F.; Nanni, B.; Zandomeneghi, M. Aging, free radicals, and antioxidants in wheat seeds. J. Agric. Food Chem. 1999, 47, 1333–1339. [Google Scholar] [CrossRef] [PubMed]
- Calucci, L.; Capocchi, A.; Galleschi, L.; Ghiringhelli, S.; Pinzino, C.; Saviozzi, F.; Zandomeneghi, M. Antioxidants, free radicals, storage proteins, puroindolines, and proteolytic activities in bread wheat (Triticum aestivum) seeds during accelerated aging. J. Agric. Food Chem. 2004, 52, 4274–4281. [Google Scholar] [CrossRef] [PubMed]
- Maluf, M.P.; Saab, I.N.; Wurtzel, E.T.; Sachs, M.M. The viviparous 12 maize mutant is deficient in abscisic acid, carotenoids, and chlorophyll synthesis. J. Exp. Bot. 1997, 48, 1259–1268. [Google Scholar] [CrossRef]
- Fenner, M.; Thompson, K. The Ecology of Seeds; Cambridge University Press: Cambridge, UK, 2005; p. 260. [Google Scholar] [CrossRef]
- Bewley, J.D.; Bradford, K.; Hilhorst, H.; Nonogaki, H. Seeds. Physiology of Development, Germination and Dormancy, 3rd ed.; Springer: New York, NY, USA, 2013; p. 392. [Google Scholar]
- Zakynthinos, G.; Varzakas, T. Carotenoids: From plants to food industry. Curr. Res. Nutr. Food Sci. 2016, 4, 38–51. [Google Scholar] [CrossRef]
- Ortuno, A.; Baidez, A.; Gomez, P.; Arcas, M.C.; Porras, I.; Garcia-Lidon, A.; Del Rio, J.A. Citrus paradisi and Citrus sinensis flavonoids: Their influence in the defence mechanism against Penicillium digitatum. Food Chem. 2006, 98, 351–358. [Google Scholar] [CrossRef]
- Jayaprakasha, G.K.; Murthy, K.N.C.; Patil, B.S. Antioxidant activities of polyphenol containing extracts from Citrus. Funct. Food Health Dis. 2008, 24, 264–276. [Google Scholar]
- Manganaris, G.A.; Goulas, V.; Vicente, A.R.; Terry, L.A. Berry antioxidants: Small fruits providing large benefits. J. Sci. Food Agric. 2014, 94, 825–833. [Google Scholar] [CrossRef]
- Silva, F.G.D.; O’Callagahan, Y.; O’Brien, N.M.; Netto, F.M. Antioxidant capacity of flaxseed products: The effect of in vitro digestion. Plant Food Hum. Nutr. 2013, 68, 24–30. [Google Scholar] [CrossRef]
- Al-Juhaimi, F.A. Citrus fruits by-products ad sources of bioactive compounds with antioxidant potential. Pak. J. Bot. 2014, 46, 1459–1462. [Google Scholar]
- Jimenez-Moreno, N.; Esparza, I.; Bimbela, F.; Gandìa, L.M.; Ancín-Azpilicueta, C. Valorization of selected fruit and vegetable wastes as bioactive compounds: Opportunities and challenges. Crit. Rev. Environ. Sci. Technol. 2019, 1–48. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Variables | TP | AC | Chl (a + b) | Car (x + c) | AsA |
|---|---|---|---|---|---|---|
| C. reticulata | TP | 1 | 0.373 | 0.094 | 0.096 | −0.09 |
| AC | 1 | 0.893 * | 0.897 * | 0. 265 | ||
| Chl (a + b) | 1 | 0.998 ** | 0.544 | |||
| Car (x + c) | 1 | 0.548 | ||||
| AsA | 1 | |||||
| C. japonica | TP | 1 | 0.643 | 0.496 | 0.469 | −0.414 |
| AC | 1 | −0.268 | −0.239 | −0.893 * | ||
| Chl (a + b) | 1 | 0.837 * | 0.546 | |||
| Car (x + c) | 1 | 0.314 | ||||
| AsA | 1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costanzo, G.; Iesce, M.R.; Naviglio, D.; Ciaravolo, M.; Vitale, E.; Arena, C. Comparative Studies on Different Citrus Cultivars: A Revaluation of Waste Mandarin Components. Antioxidants 2020, 9, 517. https://doi.org/10.3390/antiox9060517
Costanzo G, Iesce MR, Naviglio D, Ciaravolo M, Vitale E, Arena C. Comparative Studies on Different Citrus Cultivars: A Revaluation of Waste Mandarin Components. Antioxidants. 2020; 9(6):517. https://doi.org/10.3390/antiox9060517
Chicago/Turabian StyleCostanzo, Giulia, Maria Rosaria Iesce, Daniele Naviglio, Martina Ciaravolo, Ermenegilda Vitale, and Carmen Arena. 2020. "Comparative Studies on Different Citrus Cultivars: A Revaluation of Waste Mandarin Components" Antioxidants 9, no. 6: 517. https://doi.org/10.3390/antiox9060517
APA StyleCostanzo, G., Iesce, M. R., Naviglio, D., Ciaravolo, M., Vitale, E., & Arena, C. (2020). Comparative Studies on Different Citrus Cultivars: A Revaluation of Waste Mandarin Components. Antioxidants, 9(6), 517. https://doi.org/10.3390/antiox9060517