Abstract
The antioxidant activity of anthocyanins in food is well known. Numerous antioxidant assays have been proposed to measure the capacity of anthocyanins to prevent the oxidation process that naturally occurs. Different solvents, temperatures, and pH levels are applied in each assay, and these factors should be taken into account in order to obtain useful and reproducible results. The concentration and the structure of these compounds are directly related to their antioxidant capacity and their environment. However, the effectiveness of the anthocyanin ingestion against diseases is also influenced by its bioavailability. Novel methodologies that simulate the digestion process have been developed in order to facilitate the current knowledge of anthocyanins bioavailability. Studies highlight the potential synergy effect between parent compounds and their derivatives (metabolites, conjugated products, and microbe-generated metabolites). The aim of this review is to provide an overview of advantages and disadvantages of the most common methods to determine the antioxidant activity of anthocyanins, chemical structure, and concentration of these compounds in different edible fruits, vegetables, and plants; their bioavailability after intake; as well as the main therapeutic effect described in the scientific literature.
1. Introduction
In recent years, the interest in plants and food containing antioxidant properties has increased. The chemical compounds present in vegetables and fruits with these capacities are: vitamins C and E, carotenoids, and flavonoids. The anthocyanins, which are the most important group of flavonoids in plants, are pigments with a flavylium cation (AH+) structure that act as acid. This structure is directly related to its antioxidant activity. Most of the functional properties and the sensory quality of the anthocyanins can be explained by their chemical reactivity. The structures and properties of anthocyanins are dependent on different factors such as pH, temperature, and solvents which should be controlled to carry out antioxidant activity studies of these compounds [1,2,3,4,5,6,7,8,9].
Free radicals, reactive oxygen species (ROS), and/or reactive nitrogen species (RNS) are required for the proper performance of the human body and its organs. These radicals are on-balance by a redox homeostasis in our body. However, the body may be occasionally affected by an oxidative stress resulting from an off-balance state. This stress is important in the development of chronic degenerative diseases including coronary heart disease, cancer, and aging [2]. Anthocyanins have been described as compounds that prevent or inhibit, the oxidation by scavenging free radicals and reducing the oxidative stress. On a regular basis, anthocyanins act as H-atom donator or as single electron transfer. Different methods of analysis based on both mechanisms have been proposed to determine the antioxidant activity of anthocyanins. The antioxidant activity of these compounds depends on their total concentration, structure, and environment. A literature compilation about the concentrations of the most common anthocyanins in different foods is presented in this review in order to have an overview of the different sources of anthocyanins.
The beneficial properties attributed to the dietary ingestion of anthocyanin-rich foods (eye health, cardiovascular diseases, antiobesity, antidiabetic, antimicrobial, anticancer or neuroprotective effect) have been deeply documented in studies carried out with experimental models. These health benefits contrast with the apparent small portion (<1–2%) of these compounds absorbed by our organism [3,4,5]. During the digestion process, anthocyanins undergo to an intense variation in pH that together with the enzymatic and bacterial action can cause the hydrolysis and transformation of anthocyanins into metabolites, conjugated products, or simpler phenolic compounds [7,10,11,12]. The question is: How can anthocyanins be so influential in health? Are anthocyanins the only responsible of their beneficial effects? Last scientific developments highlight the potential synergy effect between parent compounds, metabolites (phases I and II), conjugated products, and microbe-generated metabolites to explain those biological events [4,11,12,13].
Due to their particular physicochemical features, bioavailability of anthocyanins is very difficult to assess. The first studies were performed analyzing blood and urine to determine the anthocyanin concentration levels after the ingestion of foods rich in anthocyanins [14,15,16]. However, the low absorption percentage obtained led to in vitro assays (mostly using cell culture systems) in order to facilitate the knowledge of their biochemical and chemical changes as well as the influence of the digestion steps. Last studies have emphasized the key role of the microbiota in the transformation of anthocyanins, which is not considered in in vitro assays but it is still poorly considered in in vivo and ex vivo studies [14].
This review aims to highlight some aspects regarding the antioxidant activity of anthocyanins and their bioavailability after intake. The first part includes an exposition of the most common antioxidant bioassays used to determine in vitro the antioxidant activity of anthocyanins, being the advantages and disadvantages of each bioassay identified. Afterwards, the effect of the chemical structure and the environment in the ability of the anthocyanins to prevent oxidation is discussed and presented together with information about different sources and range of concentration of these compounds in food. The second part of the manuscript exposes the bioavailability and metabolism of anthocyanins as well as a summary including the main therapeutic effects of anthocyanins on different diseases.
2. Antioxidant Bioassays for Anthocyanins
Numerous antioxidant assays have been proposed to measure the ability of anthocyanins to prevent the oxidation process that naturally occurs. Depending on the source of the anthocyanins and their nature, in most of the cases, an extraction step before carrying out the antioxidant bioassay is needed. The extraction process is a critical step in the determination of the antioxidant activity bioassay, presenting a challenge due to the low stability of anthocyanins after extraction and their tendency to remain bound to the matrix of the sample. Multiple alternatives have been proposed in literature for this procedure [17]. Results show that temperature, pH, solvent system, solvent-to-solid ratio, and number of extractions are factors that play an important role in the extraction efficiency and that should be optimized for each sample [18,19]. Once the anthocyanins are in liquid solution, the antioxidant activity could be determined by different bioassays. In general, two different mechanisms can be used to explain the antioxidant activity of anthocyanins: Hydrogen atom donator (HAT) and single-electron transfer (SET). In HAT mechanism, the free radical R• removes a hydrogen atom from the antioxidant (AH+) converting the free radical to a more stable product. In the SET mechanism, the antioxidant (AH+) donates an electron to the free radical reducing the oxidized intermediates into the stable form [20]. However, the difficulty in distinguishing between HAT and SET reactions is high. In most situations, these two reactions take place simultaneously, and the mechanism of the reaction is determined by the antioxidant structure, solubility, the partition coefficient, and solvent polarity [21]. Different antioxidant bioassays have been commonly used in the literature to determine the antioxidant activity of anthocyanins. A summary of the most common antioxidant bioassays used for anthocyanins is shown in Table 1.

Table 1.
Commonly used methods for measurement in vitro of antioxidant activity
Each one of these methods provides information about the antioxidant activity of the anthocyanins under specific conditions. In order to have more information about the reaction mechanism, a more comprehensive description of the procedure followed in each one of these bioassays are presented together with their main advantages and disadvantages.
2.1. DPPH (Diphenyl-1-Picrylhydrazyl) Assay
DPPH assay is a spectrophotometric method that can be applied for both solid and liquid samples, not being specific for any particular antioxidant. Thus, it can be applied to determine the total antioxidant capacity of the sample. This assay is based on the ability of the free radical (DPPH•) to react with hydrogen donor (AH+). The free radical presents an intense abortion in the UV–vis spectral region at 515 nm, the absorbance at 515 nm decreases when the free radical is reduced.
The antioxidant activity of anthocyanins for DPPH has been assessed by several authors [22,23,24,25,26]. Reliable results can be obtained with this method in a fast and simple manner. Currently, a colorimetric alternative has been proposed to extend the application of this assay where a spectrophotometer is not available [27]. Furthermore, as the free radical is stable its production is not necessary every time that an analysis is carried out. The main disadvantages could be the complexity of the analysis, as it turns more complex when other compounds present in the sample absorb also at 515 nm and many antioxidants are inert to DPPH.
2.2. ORAC Assay (Oxygen Radical Absorbance Capacity)
ORAC assay is a fluorescence method that combines the sample (AH+) with a fluorescent compound, protein phycoerthrin (β-PE) and with a generator of free radicals, 2,2′-azobis(2-amidino-propane) dihydrochloride (AAPH). This assay is based on the loss of the fluorescent compound when it is oxidized by the generated free radicals. When antioxidants (AH+) are involved in the reaction, the fluorescent compound is protected from oxidative degradation. Hence, the fluorescence signal remains. The fluorescence signal is monitored during 1 h at λem = 565 nm and λex = 540 nm respectively. The antioxidant activity of anthocyanins has been assessed by ORAC assay by several authors [26,28,29,30]. This is considered a good method to determine the antioxidant capacity of hydrophilic and hydrophobic samples, as it is easily adaptive to different samples changing the generator of free radicals. One disadvantage is the non-specificity of the fluorescence compounds that can react with the sample losing fluorescence even without the addition of a free radical generator.
2.3. TRAP Assay (Total Peroxyl Radical Trapping Antioxidant Parameter)
TRAP assay is a chemiluminescence method that consists of the following components: (i) thermolabile azo-radical initiator (e.g., 2,2′-azobis(2-amidopropane) hydrochloride (ABAP)), which produces radicals (R•) that react rapidly with O2 to give a peroxyl radicals (ROO•); (ii) oxidizable compounds with chemiluminescence properties to monitor the reaction progress (e.g., Luminol); and (iii) the sample with the antioxidant properties (AH+).
The antioxidant activity of anthocyanins has been assessed by TRAP assay by several authors [31,32,33,34]. This assay is sensitive to all known chain-breaking antioxidants. However, an important disadvantage to highlight is the difficulty comparing the results between laboratories due to the amount of different end points that can be used. It is relatively complex method, time-consuming, and requires a high degree of experience. Furthermore, and such as ORAC assay, the oxidizable compounds are non-specific and other non-radical chain reaction could occur.
2.4. FCT (Ferric Thiocyanate) Assay
The FTC assay is a spectrophotometric method that consists of the oxidation of ferrous chloride to ferric ion by reacting with peroxide. The peroxides are formed during the reaction that takes place when the sample is mixed with ethanol, water, phosphate buffer solution (pH = 7), and linoleic acid. Then, ferrous chloride in hydrochloric acid is added to the reaction and the ferric ion formed is combined with ammonium thiocyanate producing ferric thiocyanate, which is red. The absorbance of the sample is measured at 500 nm until the maximum value is reached.
This assay is used to measure the amount of peroxide produced during the initial stages of oxidation. In the case of the anthocyanins this method has been applied to determine the antioxidant activity avoiding the peroxidation of polyunsaturated fatty acid [35]. This assay is simple and reproducible. Nevertheless, results are not reliable in the case that compounds within the sample absorb around 500 nm. This drawback is common not only in this method, but in other spectrophotometric assays.
2.5. FRAP (Ferric Reducing Antioxidant Power) Assay
FRAP assay is a colorimetric method that can be employed for the determination of the total antioxidant activity of anthocyanins. It is based on the reduction of complexes of 2,4,6-tripyridyl-s-triazine (TPTZ) with ferric chloride hexahydrate (FeCl3·6H2O) under acidic conditions. The solution turns slightly brownish, forming blue ferrous complexes once the reduction is completed. The absorbance is measured at 593 nm against the blank.
This method has been extensively applied to determine the antioxidant activity of anthocyanins in different matrices: in elderberry [36]; in Roselle extract [37]; in raspberries, blackberries, red currants, gooseberries, and Cornelian cherries [38]; or in carrots, cabbage, cauliflower, potatoes, onions, asparagus and eggplant [39]. However, the results of this method are in the most of the cases compared to the results of other antioxidant assays [40]. Rapid and reproducible results are obtained with this method. However, some limitations related to FRAP assay should be considered. On the one hand, the samples must be aqueous and the pH value is critical. On the other hand, some compounds without antioxidant properties can reduce Fe3+ to Fe2+ inducing an overestimation of the antioxidant activity of the sample.
2.6. CUPRAC (Cupric Ion Reducing Antioxidant Capacity) Assay
CUPRAC assay is a spectrophotometric method similar to FRAP. In this method the anthocyanins react with the CUPRAC reagent (cupric neocuproine) producing the Cu(I)-neocuproine which is a chromophore that absorbs at 450 nm. This method has been successfully applied to various food extracts [7].
The main advantages of this method is related to the positive characteristics of the CUPRAC reagent: availability and easy accessibility, rapidity, stability, low-cost, sensitivity towards thiol-type antioxidants unlike FRAP, and responsiveness to both hydrophilic and lipophilic antioxidants. Nevertheless, the detection is based on the absorption at 450 nm which is non-specific enough and the presence of other compounds present in the sample could be interfered in the results.
2.7. ABTS (2,2′-Azino-bis (3-ehtylbenzothiazoline-6-sulfonic acid) Diamonium Salt) Assay
The most recent ABTS assay method is based on decolorization techniques. It consists of the production of a stable radical, blue/green ABTS chromophore, by the reaction of ABTS with potassium persulfate. This stable radical has a maximum absorbance at 415 nm. A drop in absorbance of this compound occurs when the radical reacts with the antioxidant. This method can determine the antioxidant activity of mixtures of substances, helping to distinguish between additive and synergistic effects. The antioxidant activity is calculated relatively to the reactivity of Trolox standard under similar conditions.
This assay is frequently combined with DPPH assay for the determination of antioxidant activity of anthocyanins [41,42,43]. This method is simple and not a large sample volume and time of analysis are necessary. Nevertheless, a standard solution is required in order to obtain accurate results.
This assay is also described in the literature as TEAC [6-hydroxy-2,5,7,8-tetramethylchroman-2-carboxylic acid (Trolox)] equivalent antioxidant capacity. The same free radical is used both TEAC and ABTS assays. However, different reagents have been proposed for the generation of the green–blue ABTS•+, resulting in different TEAC assays. This radical is produced by the oxidation of 2,2′-azinobis (3-ethylbenzothiazoline-6-sulfonic acid; ABTS). The oxidation can be reached in different ways: (i) in TEAC assay I, metmyoglobin reacted with H2O2 generating the ferrylmyoglobin radical, which then reacted with ABTS. (ii) In TEAC assay II, the ABTS•+ is formed by the reaction with manganese dioxide. (iii) In TEAC assay III, enzymatic reaction using horseradish peroxidase is applied. Other proposals consist of applying electrochemical oxidation or using 2,2′-azobis-2-amidinopropane, dihydrochloride (AAPH) or potassium persulfate (K2S2O8) as oxidants.
These alternatives are interchangeable when the appropriate solvent is selected. However, differences in assay conditions can be found, such as the reaction time or the wavelength used for the detection, for instance, sometimes in order to avoid interferences 734 nm is preferable to 415 nm.
These assays have been used to determine the antioxidant capacity of the anthocyanins, as a consequence some results has been reported in wine [44]; in corn [45]; in pomegranate juice [46] or in blueberries [47].
The advantages of TEAC assay I are the simplicity, reproducibility, and flexibility to determine the antioxidant capacity in hydrophilic and lipophilic foods. However, the pre-addiction of the sample before the radical generation could result in an overestimation of the antioxidant capacity. Other possible disadvantages of this assay may be the fact that ABTS is not found naturally, and that any compound with a redox potential lower than ABTS•+ may react with the radical.
Other alternatives applied to determine the antioxidant activity of anthocyanins involve the use of enzymes [48] or the use of chromatographic techniques. The latter alternative can not only extract the anthocyanins to determine their antioxidant activity [49], but also determining the total anthocyanins concentration (TAC), and the identification and quantification of the presence of each anthocyanin presents in the sample [50,51].
The in vitro assays exposed in this manuscript provide information about the antioxidant activity of the anthocyanins determined by their capacity to neutralize the initiators of the oxidation process (absorbing photons, neutralizing ROS or chelating metals ion) stopping the initiation steps of the autoxidation process. Thus, these assays are based on the reaction of the anthocyanins with: (i) some colored persistent radical such as the free radicals used in DPPH test and TEAC test; or (ii) other oxidizing agents like Fe3+ ions used in FRAP test or Cu2+ ions used in CUPRAC test. This neutralization capacity is measured under specific conditions of temperature, light, or the combination of both. Other assays to measure the antioxidant capacity of anthocyanins are based on a competitive probe reaction where the competitive reaction of radicals with the anthocyanins or with a probe, is monitored by fluorimetric techniques in the case of the ORAC assay, among others. All of these methods offer information on the actual antioxidant activity. They trap free radical and should be considered as chain-breaking antioxidants. This property can be measured properly only with methods based on inhibited autoxidation, while others (such as DPPH, ABTS) provide only a rough estimation. However, different specific studies have been carried out to determine the preventive antioxidant capacity [52]. Other specific assays to gather information about the inhibition of lipid substrate oxidation, where different factors and mechanism of oxidation are studied simultaneously, can be found in the literature [53].
As it has been explained above, the assays presented in this manuscript follow different mechanisms of reaction to measure the antioxidant capacity. Therefore, the information provided by them and the interpretation of their results should be carefully considered. For example, that is the case of the results provided by FRAP and CUPRAC assays versus ORAC assay which consist in a radical-trapping reaction and directly measures the capacity to neutralize initiators.
Despite the fact that the methods mentioned above are the most commonly used to determine the antioxidant capacity of anthocyanins, there is currently a claim for the development of different alternatives in order to provide direct information about the capacity to prevent the autoxidation in the biological system. Diverse alternatives have been proposed to measure the inhibition of autoxidation. That is the case exposed by Matera et al. [54], where the antioxidant activity is measured by studying the inhibited autoxidation in aqueous micelles. For that purpose, linoleic acid and Triton-X100 were mixed under controlled pH together with an initiator and the extracted anthocyanins. The reaction was tracked by monitoring the oxygen consumption with a Clark-type electrode. Another alternative to measure the capacity of anthocyanins to inhibit autoxidation was proposed by Tisuda et al. [55], where the antioxidative activity was measured by using linoleic acid autoxidation in different systems; liposome, rabbit erythrocyte membrane, and rat liver microsomal. This type of study provides direct information about the relation between the results of antioxidants assays and their biological activity in the prevention of the autoxidation. Currently, the European Food Safety Authority (EFSA) has published a guidance for the scientific requirements for health claims related to antioxidants, oxidative damage, and cardiovascular health. In this guidance, the European Health Claims Regulation has given recommendation to determine the biological effect of food supplements with purported antioxidant activity. According to this document, the antioxidant properties of foods are based on scavenging free radicals that are in vitro studied in model systems and it is not established that this capability has a beneficial physiological effect in humans. Thus, the in vivo human studies in plasma using methods—such as TRAP, TEAC, FRAP, and ORAC—carried out to establish the antioxidant capacity of the plasma do not demonstrate that it has a beneficial physiological effect in humans. Consequently, the protection of cells from premature ageing to promote healthy ageing in relation to the antioxidant properties of food are not sufficiently defined because they are established by non-specific criteria. In order to provide information about the prevention of the autoxidation under specific criteria, the EFSA guidance recommend some assays for specific biological molecules. For example, it proposes: (i) the monitoring of F2-isoprostanes to determine the capacity for the protection of the lipids from oxidative damage; (ii) the analyses of 8-hydroxy-2-deoxy-guanosine in blood, tissue and urine to assess oxidative damage to DNA; and (iii) the monitoring of protein carbonyls by ELISA, that can be applied to determine the protection of protein from oxidation damage [56].
All the assays explained in detail in this manuscript provide results that generally express the antioxidant activity as mmol Trolox Equivalent per kg of fresh weight. However, these results do not report exactly about the antioxidative activity of foods. For this purpose, information about the size portion of each food in the diet is also important to evaluate its antioxidant activity [57]. Due to the relevance of this type of results with regard to the antioxidant activity of anthocyanins, the USDA National Nutrient Database for Standard Reference has published a database where the total concentration of anthocyanins of different fruits and vegetables has been determined together with the service size that should be ingested from each fruit or vegetable [58].
3. Classification and Natural Sources of Anthocyanins
Health and therapeutic effects of anthocyanins are related to their chemical and biochemical reactivity, which are partially explained by their antioxidative activities [59,60]. However, the antioxidative activity of anthocyanins does not necessarily transfer to biological activity because any actions on the body depends both on bioavailability and cellular molecular targets [61]. Furthermore, not all the blue, red, and purple fruits, vegetables, and flowers have the same composition and concentration of anthocyanins and in consequence the same antioxidative activity. The fruits with the highest concentration of anthocyanins are berries, currants, grapes, and some tropical fruit. In the group of edible vegetables, leafy vegetables, grains, roots, and tubers show the highest concentration of anthocyanins as well [62]. Furthermore, the presence of anthocyanins can be detected in different parts of the plant such as stem, leaves, and storage organs.
In nature, these pigments are commonly present as anthocyanin, which is in the form of glycoside, and as anthocyanidin also known as aglycone (one or more saccharide bonded with the aglycone). The base structure of anthocyanins is shown in Figure 1. Nowadays the number of anthocyanins identified in nature is higher than 600 [6]. Among these anthocyanins, glycoside forms of delphinidin, cyanidin, petunidin, peonidin, malvidin, and pelargonidin, are the most abundant [54,55,62,63,64,65]. Table 2 shows the most common anthocyanins that have been identified in different fruits, vegetables, and edible flowers and the numerical code that has been assigned to each anthocyanin to refer them in the manuscript.
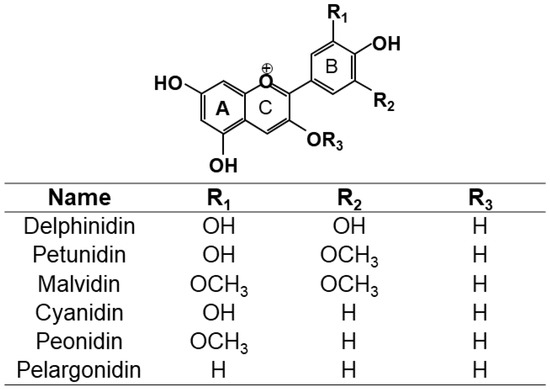
Figure 1.
Structure of anthocyanins R3 = sugar, and anthocyanidins R3 = H.
Table 2.
Anthocyanins identified in different fruits, vegetables, and edible flowers and their codes.
In supplementary information, Tables S1–S6 show the most common anthocyanins identified in different fruits, vegetables, and flowers, grouped according to their chemical structure: delphinidin and its derivatives (Table S1), cyanidin and its derivatives (Table S2), petunidin (Table S3), peonidin (Table S4), malvidin (Table S5) and pelargonidin and its derivatives (Table S6). Each table also indicates the natural source of the anthocyanins, the type of extraction that was applied, the chromatographic method used to identify and quantify them and the antioxidant assays applied to determine the antioxidant capacity.
The main conclusion from Table S1 is that delphinidin 3-glucoside (code 4) is the most common and abundant delphinidin in fruits and edible flowers. However, this group of anthocyanins is not so frequent in vegetables, being delphinine the most common. The results showed in Table S2 pointed out that the most common cyanindin in fruit and edible flower is cyanidin 3-glucoside (code 11), being also abundant cyanidin 3-galactoside (code 10) in fruits. In vegetables, cyanidin (code 8) is the most common and abundant but cyanidin 3-glucoside (code 11) is abundant in some grains as well. Table S3 shows that the following anthocyanins can be identified in fruits: petunidin 3-arabinoside, petunidin 3-galactoside, and petunidin 3-glucoside (code 30, 31, 32) in almost all the studied fruits, with petunidin 3-glucoside (code 32) being the most abundant and the only petunidin identified in the flowers. Petunidin (code 29) is the only anthocyanin of this group identified in the studied vegetables. Table S4 reveals that no peonidin was identified in flowers. In fruits, peonidin 3-glucoside (code 37) was the most abundant but also peonidin 3-galactoside (code 36) was identified in the most of them. In vegetables, the most common anthocyanin of this group was peonidin (code 35). From the group of malvidin (Table S5) it can be concluded that the most abundant in fruits is malvidin 3-galactoside (code 51) followed by malvidin 3-glucoside (code 52). The last stage was also the most abundant in flowers followed by malvidin 3,5-diglucoside (code 53). In vegetables, the malvidin was the only anthocyanin quantified of this group. Table S6 shows that pelargonidin 3-glucoside (code 55) was the most abundant in fruits followed by pelargonidin 3-rutinoside (code 56). The pelargonidin 3,5-diglucoside (code 57) was the most abundant in flowers and the pelargonidin (code 54) was the only anthocyanin of this group identified in the studied vegetables.
The protection of these pigments against oxidation process depends on their structures. Not all of them possess the same activities to scavenge diverse reactive oxygen or nitrogen species. The antioxidant ability of anthocyanins depends on the ring orientation since it will determine the willingness to donate a proton and the capacity to transfer and electron. The number of free hydroxyls around the pyrone ring and their positions also play a key role in the antioxidant activity [2]. The presence of other types of radicals in the main structure has an important role in the antioxidant activity as well. Hence, anthocyanins chalcones and quinoidal bases with a double bond conjugated to the keto group are efficient antioxidants at scavenging free radicals. Also, the glycosylated B-ring structure of anthocyanins contributes to the high antioxidant activity, where orthohydroxylation and methoxylation substantially increase the antioxidant activity. Furthermore, anthocyanidins have higher antioxidant activity in comparison with anthocyanins, which has been reported in the literature. The reason may be the lower stability of the anthocyanidin compared to the anthocyanin due to its structure, what consequently makes anthocyanidin highly reactive [66]. Acylation of anthocyanin with one or more phenolic acids has a significant increase in antioxidant activity [54,67], but glycosylation leads to a reduction in the activity [66,68].
The efficacy of scavenging diverse free radicals differs from one anthocyanin to the other. Pelargonidin-3-glucoside, cyanidin-3-glucoside, and delphinidin-3-glucoside and their standard aglycones have strong antioxidative activity in a liposomal system and reduced formation of malondialdehyde by UVB irradiation [55,69]. Furthermore, the results pointed out the highest inhibitory effect on lipid peroxidation and O2• scavenging activity of delphinidin and delphinidin-3-glucoside followed by cyanidin and pelargonidin [2]. On the contrary, pelargonidin had the highest inhibitory effect on hydroxyl radical scavenging activity [66]. Moreover, a study demonstrates the highest inhibitory effect on copper (II)-induced low-density lipoprotein (LDL) oxidation of cyanidin and cyanidin-3-glucoside compared with other phenolic acids, anthocyanins, and anthocyanin aglycones, whereas delphinidin has intermediate efficacy [70]. Another study analyzed the antioxidant activity of malvidin-3-glucoside and the result showed that the quinoidal-base and pseudo-base of malvidin-3-glucoside significantly inhibited peroxidation of linoleate compared with catechin, malvidin, and resveratrol [71]. However, the oxidation activity assigned to these anthocyanins is dependent on the type of reactive species and in consequence on the type of antioxidant assay carried out for the determination of the antioxidant activity. Thus, FRAP and TEAC assays have reported the significant reduction of the antioxidant activity by the metoxilation in the position 5 or 3 and 5 in petunidin and malvidin monoglucoside respectively [72]. Another factor implicated in the reactivity of the anthocyanins and also in their antioxidative activity is the pH. In the literature different studies have demonstrated the effect of the pH in the antioxidant capacity of the anthocyanins from different sources; Roselle [26], wine [71,72], black rice complexed with cycloamylose [73,74], palm juice [75], and Hibiscus acetosella [76]. The pH is an important factor that should be controlled in order to determine the reactivity of the anthocyanins. The acid nature of the anthocyanin structure is shown in Figure 2. This acid nature is due to the conjugation of the double bonds in the rings of the main structure and the hydroxyl groups at C4′, C5, and C7 respectively. The hydroxyl group at C7 is the strongest acid. The deprotonation can be produced at acid pH~4 yielding a neutral quinonoid base stabilized by tautomerization with the hydroxyl group at C5. The hydroxyl group at C4 is also susceptible to be deprotonated at higher pH~7 yielding the anionic base. If the pH level is still rising to basic pH, higher than 8, the deprotonation is produced in the C5 yielding the dianionic base which can lead to the chalcone anion [1]. Hence, the pH of the solution controls the proportions of protonated and deprotonated hydrated and isomeric form of anthocyanin, affecting its reactivity. Therefore, the pH should be controlled during the extractions process of anthocyanins and also during the antioxidative bioassays because their results are pH dependent [2,26].
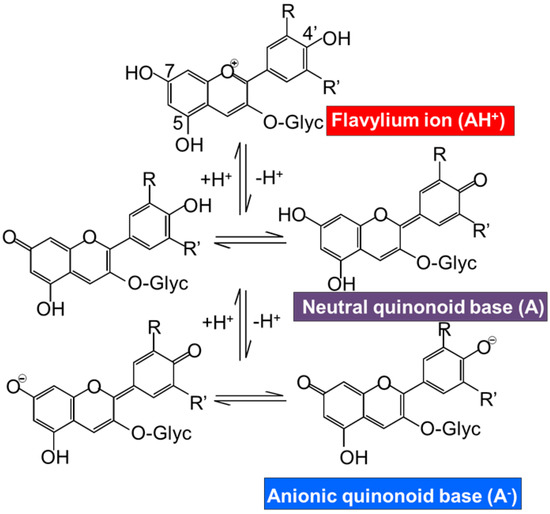
Figure 2.
Effect of pH on the structure and color of anthocyanins.
Once the identification of the most common anthocyanins in different foods has been carried out and the influence of the structure and the environment in the capacity of anthocyanins to prevent the oxidation has been discussed, the next step was to determine the bioavailability of these anthocyanins and their implication, enhancing human health.
4. Bioavailability of Anthocyanins
The daily intake of anthocyanins can be estimated via food databases and can range from few to hundreds of milligrams per person due to the methodological differences in the assessment, together with the influence of nutritional, cultural, and social differences of the investigated populations [8]. The pattern followed by anthocyanins after oral dispensation is unique and different from other flavonoids [77]. Anthocyanins have a markedly low bioavailability, only 1–2% of the ingested anthocyanins maintain their parent C6–C3–C6 structure in the organism. Food digestion is a pH-dependent process and, therefore, anthocyanins are subjected to transformations in addition to hydrolyzation by several enzymes in the small intestine [12,78]. A portion of the ingested anthocyanins reaches the large intestine, where they are metabolized into low-molecular-weight catabolites, which can be excreted in the feces within 2–4 h (up to 8 h) or absorbed again. Active transporters through either gastric or intestinal cell barrier play an important role in their transfer and absorption within the liver, kidney, brain, or other organs and tissues, besides the stomach [13,79]. In a recent review on tissue bioavailability in animals, Sandoval-Ramírez et al. [80] concluded that the TAC absorbed was 2.17 × 105 pmol/g in mice kidney, 1.73 × 105 pmol/g in liver, 3.6 × 103 pmol/g in heart, and 1.16 × 105 pmol/g in lung; and 6.08 × 103 pmol/g in pig brain. In the wall of the intestine and then in the liver, anthocyanins and their catabolites undergo phase 2 enzymatic metabolism being also transformed into their glucuronidated, sulphated, and methylated forms [10,12,13,14,78,81,82,83]. The presence of microbial catabolites at many sites of the body, at higher concentration than the native form, has suggested that part of the biological activities attributed to anthocyanins is related to the synergetic effect of their colonic catabolites [13,84]. Anthocyanin metabolites and transformation products have been characterized and quantified by several authors [85,86,87,88,89]. Ferrars et al. [88] identified a wide variety of anthocyanin phenolic metabolites, including 11 novel metabolites, in post-menopausal women after 12 weeks elderberry intake, at concentration levels higher than their anthocyanin native forms. There are many critical factors affecting the fate of anthocyanins and their metabolites in our organism: the ability to cross membranes, pH, digestive enzymes, microbiota, biliary acids, or food matrix. The use of radiolabeled (14C) or stable isotope–labelled (13C) tracer studies provides useful information about in which extent anthocyanins are metabolized to phenolic acid derivatives. In this sense, Czank et al. [90] investigated the fate of anthocyanins in eight male participants after the ingestion of 13C-cyanidin-3-O-glucoside (500 mg). The relative mean bioavailability was 12.38% (5.37% excreted in urine and 6.91% in breath). The authors found maximum serum concentration 42-fold higher for 13C-labeled metabolites than their respective native compound 13C-cyanidin-3-glucoside. Up to 49 metabolites were detected including among others: phase II conjugates of cyanidin-3-glucoside and cyanidin (cyanidin-glucuronide, methyl cyanidin-glucuronide, and methyl cyanidin-3-glucoside-glucuronide); degradation products (protocatechuic acid, phloroglucinaldehyde, and phloroglucinaldehyde); phase II conjugates of protocatechuic acid, phenylacetic acids, phenylpropenoic acids, and hippuric acid.
The mechanisms through which anthocyanins may exert their bioactivity are not fully understood as it is not clear whether their activity is linked to native forms, their derivatives, or both. The distinction of their different biological roles is a very challenging task. Some comparative studies have been conducted on the antioxidant activity of anthocyanin metabolites [91]. Recently, Kim et al. [92] provided basic information of the chemical changes of cyanidin glycosides during in vitro gastrointestinal digestion. Cyanidin-3-O-galactoside was degraded into caffeoylquinic acid, which was not found after in vitro digestion of cyanidin-3-O-glucoside. The bioactivity (DPPH) of the anthocyanin metabolites decreased in the intestinal fraction. However, the bioactivity increased after simulated colonic digestion, possibly because of the newly formed colonic metabolites. Furthermore, anthocyanin metabolites from the chokeberry extract exhibited higher DPPH radical activities than those from the mulberry extract. In another study, α-glucosidase inhibitory activity and ROS scavenging activities of conjugated-pelargonidin-3-O-glucoside samples were potentially increased after gastrointestinal digestion [93].
A scheme on the physicochemical reactions observed during the three main stages of the human digestion process can be observed in Figure 3 [11]. Biotransformation reactions start in the oral cavity through salivary amylase (pH 5.6–7.9). Once in the stomach at pH 1.5–3.5, anthocyanins exist in multiple ionic forms being mainly present as red flavylium cations and quinoidal blue species. Finally, in the intestinal step (pH 6.7–7.4) anthocyanins are present as colorless carbinol (with limited absorption) and occur the biotransformation into low molecular weight molecules such as phenolic acids or catechol (gallic acid, vanillic acid, protocatechuic acid, 4-hydroxybenzoic acid, and syringic acid have been identified as the main degradation products of delphinidin-3-O-glucoside, peonidin-3-O-glucoside, cyanidin-3-O-glucoside, pelargonidin-3-O-glucoside and malvidin-3-O-glucoside, respectively) [13].
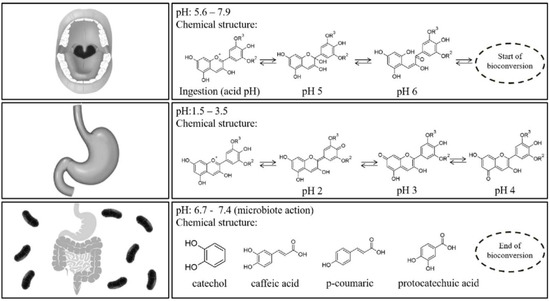
Figure 3.
Schematic representation of the anthocyanins chemical structures influenced by the digestion process steps (R2 and R3 = H or Methyl) (taken from Braga et al. [14], with permission of Elsevier).
For a better understanding of the anthocyanin bioavailability, different in vivo and in vitro models simulating digestion have been proposed [11]. Gowd et al. [94] assessed the phenolic profile of blackberry anthocyanin extract followed by human gut microbiota fermentation at different time intervals (0–48 h). Authors revealed the formation of gut metabolites enhance the high glucose plus palmitic acid induced ROS, mitochondrial membrane collapse, and glutathione depletion in HepG2 cells. Several studies have also reported that after anthocyanin colonic fermentation occurs an increase of beneficial bacteria (Bifidobacterium spp., Actinobacteria, Bacteroidetes, Lactobacillus/Enterococcus spp., Akkermansia) [95,96,97,98,99,100]. Intestinal microbiota possesses β-glucosidase activity, allowing the release of glucose from the aglycone and providing energy to support bacterial growth. A study recently carried out by Zhou et al. [95] suggests that the consumption of blueberry and its extracts could exert prebiotic activity and a modulatory effect on the composition and abundance of human intestinal microbiota. Anthocyanins could enhance human health by modulating gut microorganisms, which are often related to different diseases [95,101]. Nevertheless, it is important to note that anthocyanin derivatives can also reduce some harmful bacteria such as C. histolyticum after colonic fermentation [101,102]. A summary of anthocyanins colonic metabolism metabolites and colon microbiota alteration is shown in Figure 4 [11].
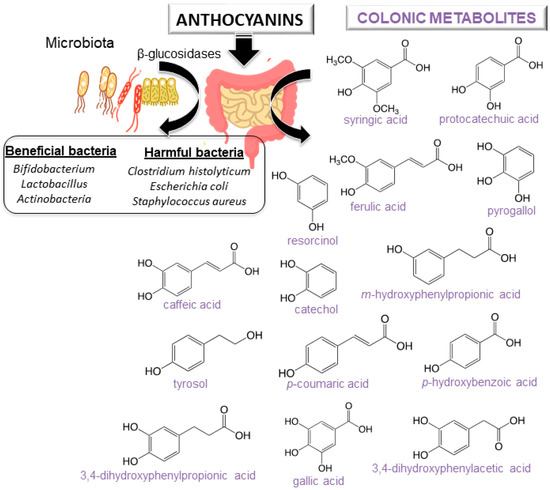
Figure 4.
Scheme of anthocyanins metabolites from colonic metabolism and colon microbiota alteration.
In any case, there are still many doubts on mechanisms involved and which factors have crucial impact on bioavailability [7]. Anthocyanins with efficient effect for one individual may not have the same effect for another [12] and there is still a high variability in the results obtained. Based on the literature and recent reviews [11,14] this variability is due to the lack of homogeneity introduced at three levels: (i) food matrix and food processing; (ii) enzymatic levels (affected by genetic factors and diet, age, and sex); and (iii) microbiota functionality. Reported data considering inter- or intra-individual variability is very scarce and bioavailability methods are not standardized making very difficult to reach firm conclusions. On the one hand, in vitro methods (cell-based assays) fail to consider the role of the individual microbiota present in the human body; while, on the other hand, in in vivo trials (human trials and animal studies) each subject has their own microbiota [14].
It is also important to note that the incorporation of anthocyanins into food and medical products is a challenging task due to their high instability and susceptibility to degradation. In this sense, the use of nano/microencapsulation with natural polymers is one of the best strategies to improve the stability of sensitive substances in in vitro simulated gastrointestinal digestion and colonic fermentation [103,104]. According to a recent review on this topic [104] different techniques have been tested to encapsulate anthocyanins including spray-drying > freeze-drying > gelation > lipid-based particles > electrohydrodynamic processes. The first one is the most economical, simplest, and the most applied method (80–90%) [105,106]. The use of other techniques still remains poorly explored probably due to the hydrophilic nature of anthocyanins, being therefore a promising area of future research [104,107,108,109].
Blackberry anthocyanins encapsulated with β-cyclodextrin [101,110] or gum arabic [111] helped to delay the release of anthocyanins during in vitro simulated gastrointestinal digestion. The stability of anthocyanins can be also influenced by the type of wall material. Recently, Wu et al. [103] evaluated the effect of four different wall materials during in vitro simulated digestion and colonic fermentation. The encapsulation technique enhanced significantly the colonic accessibility and delayed the release of anthocyanins, especially for soy protein. Degradation products of anthocyanins such as syringic acid produced during colonic fermentation by the action of gut microbiota were indicative of their benefits for host health.
5. Therapeutic Effects of Anthocyanins
Available scientific studies prove the beneficial effects of the presence of anthocyanins in fruits and vegetables in the prevention of diseases [60,66,81,112]. Even after the ingestion of high doses of anthocyanin and derivatives no negative effects have been observed [113]. This section covers the main health benefits of anthocyanins in different types of pathologies including eye health, cardiovascular disease, antiobesity, antidiabetic, antimicrobial effects, anticancer activities, and neurodegenerative disorders. A summary of the positive effects of anthocyanins is shown in Table 3 [114,115,116,117,118,119,120,121,122,123,124,125,126,127,128,129,130,131,132,133,134,135,136,137,138,139,140,141,142,143,144,145,146,147,148,149,150,151,152,153,154,155,156,157,158,159,160], and their mechanisms of action in disease prevention are discussed below.
Table 3.
Health benefits of anthocyanins.
Eye health: Since the first report in 1966 about the positive effects of anthocyanins on vision in humans, anthocyanin-rich extracts have been worldwide utilized as a popular supplement for ocular health [161,162]. Oral dispensation of blackcurrant anthocyanins may be a promising supplement for patients with open-angle glaucoma, being also effective for antiglaucoma medication, while anthocyanin-rich bilberry extract has a protective effect on vision during retinal inflammation [115]. It has also been confirmed that cyanidin helps the regeneration of rhodopsin and smooth muscle relaxation in rats [116]. Results have also showed that bilberry extracts were able to suppress the photoxidation of pyridinium disretinoid A2E, an auto-fluorescence pigment that accumulates in retinal epithelial cells with age and can cause light-induced damage to the cell. In a comparative study a significant improvement on nocturnal visual function and an improved contrast sensitivity levels in subjects with myopia versus placebo group was observed [114]. Anthocyanins act also inhibiting transient myopia, reducing eye fatigue or enhancing retinal blood flow with glaucoma [118,121,161,163].
Cardiovascular diseases: It is especially important the role of anthocyanins in preventing myocardial infarction and cardiovascular disease related to mortality. Extracts of anthocyanins have been used to inhibit platelet aggregation being preventive in the initial stage of thrombi; in the treatment of problem with poor micro-circulation resulting from capillary fragility; and also to prevent the LDL oxidation [122,164,165,166]. In a placebo-controlled trial in dyslipidemia patients (40–65 years) the intake of berry-derived anthocyanins improved lipoprotein profile through cholesteryl ester transfer protein inhibition [123]. Authors observed a greater increase in high-density lipoprotein (HDL) cholesterol levels and in the cellular cholesterol efflux to serum as well as a decrease in LDL cholesterol levels in the anthocyanin group in contrast to the placebo group. Similar results were reported by Álvarez Suárez et al. [127] in an in vivo study using healthy volunteers supplemented with strawberries (500 g). Daily consumption improved the lipid profile reducing total cholesterol, LDL cholesterol and triglycerides levels, while HDL cholesterol remained unchanged. This increased antihemolytic defenses and platelet function in the subjects. In another attempt, higher intakes of fruit-based anthocyanins were associated to a lower risk of nonfatal myocardial infarction (14%) and ischemic stroke in a prospective cohort study in men over 24 years [124]. A meta-analysis of 45 randomized controlled trials stated that the consumption of berries and purified anthocyanins (2.2−1230 mg anthocyanins/day) increases significantly HDL-cholesterol and reduces LDL-cholesterol, triglycerides, systolic blood pressure, and diastolic blood pressure as well as the inflammatory markers CRP and TNFα [167]. The analysis also suggested that some individuals are more susceptible to the protective effects of anthocyanin consumption: (i) overweight; (ii) over 50 years; and (iii) those with increased risk of cardiovascular disease. Another meta-analysis of 99 randomized controlled trials showed that the consumption of anthocyanin rich-products decreased significantly both systolic and diastolic blood pressure regardless of the health status of the participants [168].
In in vitro assays, anthocyanins have also shown inhibition of the porcine pancreatic elastase [169], an enzyme that plays a significant function in pathologies such as arteriosclerosis, emphysema, or rheumatoid arthritis, etc., by attacking fibers and collagen. Moreover, acceleration in the cicatrization process due to anthocyanin-rich extract has been demonstrated, showing preventive and curative activity against gastroduodenal ulcers induced in rats [7]. Their influence on the biosynthesis of mucopolysaccharides provably improves the efficacy of the gastric mucous layer, and increases the base substance of the connective tissue and of the capillaries [170].
Antiobesity and Antidiabetic effects: Anthocyanins have shown anti-obesity effects through multiple mechanisms such as inhibiting lipid absorption, regulating lipid metabolism, increasing energy expenditure, suppressing food intake and regulating gut microbiota, which suggests anthocyanins are promising candidates in anti-obesity therapies [171]. Kwon et al. [128] observed that anthocyanins-added diet from black soybean in rats decreases body weight gains, being significantly lowered in the rats fed with a high fat diet plus black soybean anthocyanins compared with the rats fed with high fat diet without black soybean. Anthocyanins also improved the lipid profile and suppressed the high fat diet-induced weight gain in liver intermediately and decreased the weights of epididymal and perirenal fat pads.
In addition, type 2 diabetes is closely related to obesity [66]. Anthocyanins can alleviate complications in type 2 diabetes by inhibiting intestinal glucose absorption, inducing pancreatic insulin secretion, upregulating glucose transporter type 4, and suppressing hepatic gluconeogenesis [172]. After the supplementation of a high-fat diet during 13 weeks with different berries in mice, Heyman et al. [173] observed that those supplemented mice gained lesser body weight and presented lower fasting insulin levels than the control group as well as mediated positive effects on glucose homeostasis. Jankowski et al. [130] described a substantial decrease in the sugar concentration in urine and blood serum after streptozotocin injection in fed rats with grapes. The mechanisms of anthocyanins suggested by the authors were the reduction of the biosynthesis of collagen, lipoproteins, and glycoproteins, as well as the reduction of the activity of elastase and adenosine deaminase (both high in diabetic patients). Treatment with cherries in rats resulted in a significant reduction of blood glucose and urinary microalbumin and an increase of the creatinine secretion level in urea [174]. The pulp, seed and skin from “red chilto” (a red fruit from Argentina) had a hypoglycemic effect and acted increasing glucose absorption, decreasing glucose diffusion rate and promoting glucose transport across the cell membrane [175] in an in vitro simulated gastroduodenal digestion. Consumption of blueberries and apples/pears in humans was also associated to a lower risk of type 2 diabetes [176].
Antimicrobial effects: The antimicrobial activity of anthocyanins against a wide range of microorganisms is also well documented. Possible mechanisms induced cell damage by destroying the cell wall, membrane and intercellular matrix [66,140,177]. Blackberry extracts have antibacterial activity with the highest sensitivity to Aeromonas hydrophilia and Listeria innocua [141]. Cranberry extracts have antibacterial activity towards Enterococcus faecium resistant to vancomycin, Pseudomonas aeruginosa, Staphylococcus aureus, and Escherichia coli [142]. Different types of berry extracts inhibit Gram-negative bacteria but not Gram-positive bacteria [143] probably because Gram-negative bacteria acts as a preventive barrier against hydrophobic compounds but not against hydrophilic compounds [178].
Anticancer activity: Possible mechanisms of the anticancer activity of anthocyanins have been described by many authors: antimutagenic activity; inhibition of oxidative DNA damage and carcinogen activation; induction of phase II enzymes for detoxification; cell cycle arrest; inhibition of cyclooxygenase-2 enzymes; as well as induction of apoptosis and antiangiogenesis [179,180,181,182,183,184].
In breast cancer, anthocyanins cause the inhibition of key modulators that promote its progression and development by acting directly in the DNA fragmentation and promoting the death of MCF-7 cancer cells [185,186]. In addition, the studies indicate that anthocyanins exert extensive in vitro anti-invasive and in vivo anti-metastatic activities. For example, delphinidin can act as a potential antimetastatic agent that suppresses PMA-induced cancer cell invasion through the specific inhibition of NF-κB-dependent MMP-9 gene expression [187,188]. In lung cancer, the treatment of cyanidin-3-glucoside and cyanidin 3-rutinoside, isolated from mulberry, inhibits the migration and invasion of A549 cells and also decreases MMP-2 and uPA and enhances TIMP-2 and PAI. Anthocyanins also inhibit the growth of carcinogenic cells that provoke colon cancer, induce the apoptosis effect, and are even able to act as modulators of the macrophages in the immune response [180]. Forester et al. [189] also reported the positive effect of anthocyanin metabolites decreasing cell viability and causing cell cycle arrest and apoptosis in colon cancer. In oral and cervical cancer, the invasion of SCC-4 cells and HeLa cells were diminished by the treatment of peonidin 3-glucoside and cyanidin-3-glucoside [190].
It is also important to note that the structures of anthocyanins have a considerable influence on their biological activities [191,192,193]. In this sense, the type of aglycones, sugars, and acylated acids, and the position and degree of glycosylation and acylation seem to be the main factors influencing the anticancer property [191]. Jing et al. [192] compared the anticancer properties of anthocyanin-rich extracts using human colon cancer HT29 cell line. Authors reported the following growth inhibitory activity rates: purple corn > chokeberry and bilberry > purple carrot and grape > radish and elderberry. Those non-acylated monoglycosylated anthocyanins had greater anticancer property than those with pelargonidin, triglycoside, and/or acylation with cinnamic acid.
Neurodegenerative diseases: Anthocyanins are also uniquely suited for the treatment of neurodegenerative diseases such as Alzheimer’s, Parkinson’s, or amyotrophic lateral sclerosis. Their main mechanisms include antioxidant pathways, calcium homeostasis, inflammation, protein homeostasis, and the balance of pro-survival and pro-apoptotic signaling [194,195].
In a primary cell model of Parkinson’s disease, dopaminergic cell death elicited by rotenone was suppressed by extracts prepared from blueberries, grape seed, hibiscus, blackcurrant, and mulberry [154]. Moreover, Strathearn et al. [154] observed that those extracts rich in anthocyanins and proanthocyanidins exhibited greater neuroprotective activity than extracts rich in other polyphenols.
The oral dispensation of anthocyanins (200 mg/kg) in rats was able to regulate cholinergic neurotransmission, to restore Na+, K+-ATPase and Ca2+-ATPase activities, and to prevent memory deficits caused by scopolamine dispensation [156]. Rehman et al. [157] showed the neuroprotective effect of anthocyanins based on an artificial ageing model using D-galactose to induce oxidative stress and inflammatory response. The potential mechanisms of their action included: decreased expression of the receptor for advance glycation end product, reduced level of ROS, and lipid peroxidation. Shih et al. [155] observed that mice fed with anthocyanin-rich mulberry extracts demonstrated significantly less amyloid β protein and showed improvement of learning and memory ability in avoidance response tests. The fed mice also showed a higher antioxidant enzyme activity and less lipid oxidation in both brain and liver, as compared to the control mice. Besides, the treatment with anthocyanin-rich mulberry extract has been proved to decrease the levels of serum aspartate aminotransferase, alanine aminotransferase, triglyceride, and total cholesterol that increase with ageing.
Furthermore, the therapeutic profile of anthocyanins can be improved by encapsulation [158,159,160]. For instance, in Alzheimer’s disease Amin et al. [158] showed that encapsulated nanoparticles loaded with anthocyanins are rapidly taken up by cells enhancing their neuroprotective profile against amyloid beta toxicity above that of anthocyanins alone. Similar activity was also observed in in vivo studies in mice [158,160].
6. Conclusions
In order to establish the antioxidant activity of anthocyanins and how their intake affects the human health, many factors should be taken into account. Firstly, an evaluation of the antioxidant activity from a multiparametric perspective is requires as the total concentration, the structure, the nature of the sample, the pH and the mechanism of the reaction play an important role in their effect. On a regular basis, more than one antioxidant assay—one for each mechanism—should be carried out. Besides identification and quantification of the anthocyanins is also highly recommended to establish an accurate value for the antioxidant activity of a sample. Secondly, it is required to know the bioavailability of these compounds after their intake. Many studies that have demonstrated the benefits of anthocyanin-rich extracts in the prevention of diseases. Nonetheless, it is important to note that their efficacy depends on their bioavailability. Along the digestion process, anthocyanins are metabolized into various conjugates, which then ultimately metabolize into phenolic acid degradation products as well. The accumulated evidence suggests the synergy effect between all possible forms to explain their attributed health-promoting properties. An inter- and intra-individual variability in anthocyanins absorption, metabolism, distribution, and excretion is also evident. Among the main factors that probably affect this variability are: food matrix and processing, enzymatic levels and microbiota functionality. Attention should be paid in different lines: (i) to perform well-designed standardized methods when evaluating bioavailability; (ii) to consider inter or intra-individual variability in anthocyanin metabolism; (iii) to assess the antioxidant activity of anthocyanin metabolites during gastrointestinal digestion; and, since anthocyanin might exert different biological activities, (iv) to draw a more accurate characterization profiles.
Supplementary Materials
The following are available online at https://www.mdpi.com/2076-3921/9/5/451/s1, Table S1: Natural source, type of extraction, chromatographic method and antioxidant assay applied to identify and quantify delphinidin and its derivatives identified with codes, Table S2: Natural source, type of extraction, chromatographic method and antioxidant assay applied to identify and quantify cyanidin and its derivatives identified with codes, Table S3: Natural source, type of extraction, chromatographic method and antioxidant assay applied to identify and quantify petunidin and its derivatives identified with codes, Table S4: Natural source, type of extraction, chromatographic method and antioxidant assay applied to identify and quantify peonidin and its derivatives identified with codes, Table S5: Natural source, type of extraction, chromatographic method and antioxidant assay applied to identify and quantify malvidin and its derivatives identified with codes, Table S6: Natural source, type of extraction, chromatographic method and antioxidant assay applied to identify and quantify pelargonidin and its derivatives identified with codes.
Author Contributions
Investigation, N.T., J.M., and A.G.A.; Project administration, A.G.A.; Supervision, A.G.A.; Writing—original draft, N.T. and J.M.; Writing—review and editing, N.T. and J.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
The authors would like to thank the Vicerrectorado de Investigación de la Universidad de Sevilla for its support. We would also like to thank the reviewers for their helpful comments that have greatly contributed to the improvement of the document. We are also grateful for the very helpful revision by Irene García Sáez.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dangles, O.; Fenger, J.A. The Chemical Reactivity of Anthocyanins and Its Consequences in Food Science and Nutrition. Molecules 2018, 23, 1970. [Google Scholar] [CrossRef] [PubMed]
- Miguel, M.G. Anthocyanins: Antioxidant and/or anti-inflammatory activities. J. Appl. Pharm. Sci. 2011, 1, 7–15. [Google Scholar]
- Kay, C.D.; Pereira-Caro, G.; Ludwig, I.A.; Clifford, M.N.; Crozier, A. Anthocyanins and flavanones are more bioavailable than previously perceived: A review of recent evidence. Annu. Rev. Food Sci. Technol. 2017, 8, 155–180. [Google Scholar] [CrossRef] [PubMed]
- Lila, M.A.; Burton-Freeman, B.; Grace, M.; Kalt, W. Unraveling anthocyanin bioavailability for human health. Annu. Rev. Food Technol. 2016, 7, 375–393. [Google Scholar] [CrossRef] [PubMed]
- Fang, J. Bioavailability of anthocyanins. Drug Metab. Rev. 2014, 46, 508–520. [Google Scholar] [CrossRef]
- Lingua, M.S.; Fabani, M.P.; Wunderlin, D.A.; Baroni, M.V. From grape to wine: Changes in phenolic composition and its influence on antioxidant activity. Food Chem. 2016, 208, 228–238. [Google Scholar] [CrossRef]
- Martín, J.; Kuskoski, E.M.; Navas, M.J.; Asuero, A.G. Antioxidant Capacity of Anthocyanin Pigments. In Flavonoids—From Biosynthesis to Human Health; Justino, J., Ed.; Science, Technology and Medicine Open Access Publisher: Rijeka, Croatia, 2017; Chapter 11; pp. 205–255. [Google Scholar]
- Martín Bueno, J.; Sáez-Plaza, P.; Ramos-Escudero, F.; Jímenez, A.M.; Fett, R.; Asuero, A.G. Analysis and antioxidant capacity of anthocyanin pigments. Part II: Chemical structure, color, and intake of anthocyanins. Crit. Rev. Anal. Chem. 2012, 42, 126–151. [Google Scholar] [CrossRef]
- Navas, M.J.; Jiménez-Moreno, A.M.; Martín Bueno, J.; Sáez-Plaza, P.; Asuero, A.G. Analysis and antioxidant capacity of anthocyanin pigments. Part IV: Extraction of anthocyanins. Crit. Rev. Anal. Chem. 2012, 42, 313–342. [Google Scholar] [CrossRef]
- Gamel, T.H.; Wright, A.J.; Tucker, A.J.; Pickard, M.; Rabalski, I.; Podgorski, M.; Di Ilio, N.; O’Brien, C.; Abdel-Aal, E.M. Absorption and metabolites of anthocyanins and phenolic acids after consumption of purple wheat crackers and bars by healthy adults. J. Cereal Sci. 2019, 86, 60–68. [Google Scholar] [CrossRef]
- Cavalcante Braga, A.R.; Murador, D.C.; Mendes de Souza Mesquita, L.; Vera de Rosso, V. Bioavailability of anthocyanins: Gaps in knowledge, challenges and future research. J. Food Compos. Anal. 2018, 68, 31–40. [Google Scholar] [CrossRef]
- Fernandes, I.; Faria, A.; Calhau, C.; de Freitas, V.; Mateus, N. Bioavailability of anthocyanins and derivatives. J. Funct. Foods 2014, 7, 54–66. [Google Scholar] [CrossRef]
- Fernandes, I.; Faria, A.; de Freitas, V.; Calhau, C.; Mateus, N. Multiple-approach studies to assess anthocyanin bioavailability. Phytochem. Rev. 2015, 14, 899–919. [Google Scholar] [CrossRef]
- Eker, M.E.; Aaby, K.; Budic-Leto, I.; Rimac Brncˇic, S.; El, S.N.; Karakaya, S.; Simsek, S.; Manach, C.; Wiczkowski, W.; de Pascual-Teresa, S. A Review of Factors Affecting Anthocyanin Bioavailability: Possible Implications for the Inter-Individual Variability. Foods 2020, 9, 2. [Google Scholar] [CrossRef] [PubMed]
- Mülleder, U.; Murkovic, M.; Pfannhauser, W. Urinary excretion of cyanidin glycosides. J. Biochem. Biophys. Methods 2002, 53, 61–66. [Google Scholar]
- Bub, A.; Watzl, B.; Heeb, D.; Rechkemmer, G.; Briviba, K. Malvidin-3-glucoside bioavailability in humans after ingestion of red wine, dealcoholized red wine and red grape juice. Eur. J. Nutr. 2001, 40, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.T.; Netzel, M.E.; O’Hare, T.J. Optimization of extraction procedure and development of LC–DAD–MS methodology for anthocyanin analysis in anthocyanin-pigmented corn kernels. Food Chem. 2020, 319, 126515. [Google Scholar] [CrossRef] [PubMed]
- Castañeda-Ovando, A.; Pacheco-Hernandez, M.L.; Paez-Hernandez, M.E.; Rodriguez, J.A.; Galan-Vidal, C.A. Chemical studies of anthocyanins: A review. Food Chem. 2009, 113, 859–871. [Google Scholar] [CrossRef]
- Fernandez-Aulis, F.; Hernandez-Vazquez, L.; Aguilar-Osorio, G.; Arrieta-Baez, D.; Navarro-Ocan, A. Extraction and Identification of Anthocyanins in Corn Cob and Corn Husk from Cacahuacintle Maize. J. Food Sci. 2019, 84, 954–962. [Google Scholar] [CrossRef]
- Siti Azima, A.M.; Noriham, A.; Manshoor, N. Anthocyanin content in relation to the antioxidant activity and colour properties of Garcinia mangostana peel, Syzigium cumini and Clitoria ternatea extracts. Int. Food Res. J. 2014, 21, 2369–2375. [Google Scholar]
- Liang, N.; Kitts, D.D. Antioxidant Property of Coffee Components: Assessment of Methods that Define Mechanisms of Action. Molecules 2014, 19, 19180–19208. [Google Scholar] [CrossRef]
- Shalaby, E.A.; Shanab, S.M.M. Antioxidant compounds, assays of determination and mode of action. Afr. J. Pharm. Pharmacol. 2013, 7, 528–539. [Google Scholar] [CrossRef]
- Goupy, P.; Bautista-Ortin, A.-B.; Fulcrand, H.; Dangles, O. Antioxidant activity of wine pigments derived from anthocyanins: Hydrogen transfer reactions to the DPPH radical and inhibition of the heme-induced peroxidation of linoleic acid. J. Agric. Food Chem. 2009, 57, 5762–5770. [Google Scholar] [CrossRef] [PubMed]
- Molyneux, P. The use of the stable free radical diphenylpicrylhydrazyl (DPPH) for estimating antioxidant activity. Songklanakarin. J. Sci. Technol. 2004, 26, 211–219. [Google Scholar]
- Sudheeran, P.K.; Feygenberg, O.; Maurer, D.; Alkan, N. Improved Cold Tolerance of Mango Fruit with Enhanced Anthocyanin and Flavonoid Contents. Molecules 2018, 23, 1832. [Google Scholar] [CrossRef]
- Wu, H.Y.; Yang, K.M.; Chiang, P.Y. Roselle Anthocyanins: Antioxidant Properties and Stability to Heat and pH. Molecules 2018, 23, 1357. [Google Scholar] [CrossRef]
- Akar, Z.; Kucuk, M.; Dogan, H. A new colorimetric DPPH scavenging activity method with no need for a spectrophotometer applied on synthetic and natural antioxidants and medicinal herbs. J. Enzym. Inhib. Med. Chem. 2017, 32, 640–647. [Google Scholar] [CrossRef]
- Ehlenfeldt, M.K.; Prior, R.L. Oxygen radical absorbance capacity (ORAC) and phenolic and anthocyanin concentrations in fruit and leaf tissues of highbush blueberry. J. Agric. Food Chem. 2001, 49, 2222–2227. [Google Scholar] [CrossRef]
- Bellido, G.G.; Beta, T. Anthocyanin Composition and Oxygen Radical Scavenging Capacity (ORAC) of Milled and Pearled Purple, Black, and Common Barley. J. Agric. Food Chem. 2009, 57, 1022–1028. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, S.Y.; Wang, C.Y.; Zheng, W. Changes in strawberry phenolics, anthocyanins, and antioxidant capacity in response to high oxygen treatments. LWT 2007, 40, 49–57. [Google Scholar] [CrossRef]
- Denev, P.; Ciz, M.; Ambrozova, G.; Lojek, A.; Yanakieva, I.; Kratchanova, M. Solid-phase extraction of berries’ anthocyanins and evaluation of their antioxidative properties. Food Chem. 2010, 123, 1055–1061. [Google Scholar] [CrossRef]
- Rossetto, M.; Vanzani, P.; Lunelli, M.; Scarpa, M.; Mattivi, F.; Rigo, A. Peroxyl radical trapping activity of anthocyanins and generation of free radical intermediates. Free Radic. Res. 2007, 41, 854–859. [Google Scholar] [CrossRef] [PubMed]
- Dresch, R.R.; Kreinecker-Dresch, M.T.; Biegelmeyer, R.; Fretes-Argenta, D.; Fagundes da Rocha, R.; Ferreira-Teixeira, H.; Fonseca-Moreira, J.C.; Henriques, A.T. Potential use of secondary products of the agri-food industry for topical formulations and comparative analysis of antioxidant activity of grape leaf polyphenols. Nat. Prod. Res. 2018, 32, 486–492. [Google Scholar] [CrossRef] [PubMed]
- Philpott, M.; Lim, C.C.; Ferguson, L.R. Dietary Protection Against Free Radicals: A Case for Multiple Testing to Establish Structure-activity Relationships for Antioxidant Potential of Anthocyanic Plant Species. Int. J. Mol. Sci. 2009, 10, 1081–1103. [Google Scholar] [CrossRef] [PubMed]
- Su, M.S.; Chien, P.J. Antioxidant activity, anthocyanins, and phenolics of rabbiteye blueberry (Vaccinium ashei) fluid products as affected by fermentation. Food Chem. 2007, 104, 182–187. [Google Scholar] [CrossRef]
- Özgen, M.; Scheerens, J.C.; Neil Reese, R.; Miller, R.A. Total phenolic, anthocyanin contents and antioxidant capacity of selected elderberry (Sambucus canadensis L.) accessions. Pharm. Mag. 2010, 6, 198–203. [Google Scholar] [CrossRef]
- Tsai, P.J.; Intosh, J.M.; Pearce, P.; Camden, B.; Jordan, B.R. Anthocyanin and antioxidant capacity in Roselle (Hibiscus Sabdariffa L.) extract. Food Res. Int. 2002, 35, 351–356. [Google Scholar] [CrossRef]
- Pantelidis, G.E.; Vasilakakis, M.; Manganaris, G.A.; Diamantidis, G. Antioxidant capacity, phenol, anthocyanin and ascorbic acid contents in raspberries, blackberries, red currants, gooseberries and Cornelian cherries. Food Chem. 2007, 102, 777–783. [Google Scholar] [CrossRef]
- Li, H.; Deng, Z.; Zhu, H.; Hu, C.; Liu, R.; Young, J.C.; Tsao, R. Highly pigmented vegetables: Anthocyanin compositions and their role in antioxidant activities. Food Res. Int. 2012, 46, 250–259. [Google Scholar] [CrossRef]
- Moon, J.K.; Shibamoto, T. Antioxidant Assays for Plant and Food Components. J. Agric. Food Chem. 2009, 57, 1655–1666. [Google Scholar] [CrossRef]
- Ge, Q.; Ma, X. Composition and antioxidant activity of anthocyanins isolated from Yunnan edible rose (An ning). Food Sci. Hum. Wellness 2013, 2, 68–74. [Google Scholar] [CrossRef]
- Frond, A.D.; Iuhas, C.I.; Stirbu, I.; Leopold, L.; Socaci, S.; Andreea, S.; Ayvaz, H.; Andreea, S.; Mihai, S.; Diaconeasa, Z.; et al. Phytochemical Characterization of Five Edible Purple-Reddish Vegetables: Anthocyanins, Flavonoids, and Phenolic Acid Derivatives. Molecules 2019, 24, 1536. [Google Scholar] [CrossRef] [PubMed]
- Pasko, P.; Barton, H.; Zagrodzki, P.; Gorinstein, S.; Fołta, M.; Zachwieja, Z. Anthocyanins, total polyphenols and antioxidant activity in amaranth andquinoa seeds and sprouts during their growth. Food Chem. 2009, 115, 994–998. [Google Scholar] [CrossRef]
- Villaño, D.; Fernández-Pachón, M.S.; Troncoso, A.M.; García-Parrilla, M.C. The Antioxidant Activity of Wines Determined by the ABTS (+) Method: Influence of Sample Dilution and Time. Talanta 2004, 64, 501–509. [Google Scholar]
- Hu, Q.P.; Xu, J.G. Profiles of carotenoids, anthocyanins, phenolics, and antioxidant activity of selected color waxy corn grains during maturation. J. Agric. Food Chem. 2011, 59, 2026–2033. [Google Scholar] [CrossRef]
- Pala, C.U.; Toklucu, A.K. Effect of UV-C light on anthocyanin content and other quality parameters of pomegranate juice. J. Food Compos. Anal. 2011, 24, 790–795. [Google Scholar] [CrossRef]
- Bunea, A.; Rugina, D.O.; Pintea, A.M.; Conta, Z.; Bunea, C.I.; Socaciu, C. Comparative Polyphenolic Content and Antioxidant Activities of Some Wild and Cultivated Blueberries from Romania. Not. Bot. Horti Agrobo. 2011, 39, 70–76. [Google Scholar] [CrossRef]
- Moniruzzaman, M.; Khalil, M.I.; Sulaiman, S.A.; Gan, S.H. Advances in the analytical methods for determining the antioxidant properties of honey: A review. Afr. J. Tradit. Complement. Altern. Med. 2012, 9, 36–42. [Google Scholar] [CrossRef]
- Degenhardt, A.; Knapp, H.; Winterhalter, P. Separation and Purification of Anthocyanins by High-Speed Countercurrent Chromatography and Screening for Antioxidant Activity. J. Agric. Food Chem. 2000, 48, 338–343. [Google Scholar] [CrossRef]
- Rubinskiene, M.; Jasutiene, I.; Venskutonis, P.R.; Viskelis, P. HPLC Determination of the Composition and Stability of Blackcurrant Anthocyanins. J. Chromatogr. Sci. 2005, 43, 478–482. [Google Scholar] [CrossRef]
- Welch, C.R.; Wub, Q.; Simon, J.E. Recent Advances in Anthocyanin Analysis and Characterization. Curr. Anal. Chem. 2008, 4, 75–101. [Google Scholar] [CrossRef]
- Amorati, R.; Valgimigli, L. Methods to Measure the Antioxidant Activity of Phytochemicals and Plant Extracts. J. Agric. Food Chem. 2018, 66, 3324–3329. [Google Scholar] [CrossRef] [PubMed]
- Félix, R.; Valentão, P.; Andrade, P.B.; Félix, C.; Novais, S.C.; Lemos, M.F.L. Evaluating the in Vitro Potential of Natural Extracts to Protect Lipids from Oxidative Damage. Antioxidants 2020, 9, 231. [Google Scholar] [CrossRef] [PubMed]
- Matera, R.; Gabbanini, S.; Berretti, S.; Amorati, R.; De Nicola, G.R.; Iori, R.; Valgimigli, L. Acylated anthocyanins from sprouts of Raphanus sativus cv. Sango: Isolation, structure elucidation and antioxidant activity. Food Chem. 2015, 166, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, T.; Watanabe, M.; Ohshima, K.; Norinobu, S.; Choi, S.W.; Kawakishi, S.; Osawa, T. Antioxidative Activity of the Anthocyanin Pigments Cyanidin 3-o-β-D-Glucoside and Cyanidin. J. Agri. Food Chem. 1994, 42, 2407–2410. [Google Scholar] [CrossRef]
- EFSA. Guidance for the scientific requirements for health claims related to antioxidants, oxidative damage and cardiovascular health (Revision 1). EFSA J. 2018, 16, 5136. [Google Scholar]
- Prior, R.L.; Go, L.; Wu, X.; Jacob, R.A.; Sotoudeh, G.; Kader, A.A.; Cook, R.A. Plasma antioxidant capacity changes following a meal as a measure of the ability of a food to alter in vivo antioxidant status. J. Am. Coll. Nutr. 2007, 26, 170–181. [Google Scholar] [CrossRef]
- Comert, E.D.; Mogol, B.A.; Gokmen, V. Relationship between color and antioxidant capacity of fruits and vegetables. Curr. Res. Food Sci. 2020, 2, 1–10. [Google Scholar] [CrossRef]
- Bakuradze, T.; Tausend, A.; Galan, J.; Groh, I.A.M.; Berry, D.; Tur, J.A.; Marko, D.; Richling, E. Antioxidative activity and health benefits of anthocyanin-rich fruit juice in healthy volunteers. Free Radic. Res. 2019, 53, 1045–1055. [Google Scholar] [CrossRef]
- Khoo, H.E.; Lim, S.M.; Azlan, A. Evidence-based therapeutic effects of anthocyanins from foods. Pak. J. Nutr. 2019, 18, 1–11. [Google Scholar]
- Williamson, G. The role of polyphenols in modern nutrition. Nutr. Bull. 2017, 42, 226–235. [Google Scholar] [CrossRef]
- Chaiyasut, C.; Sivamaruthi, B.S.; Pengkumsri, N.; Sirilun, S.; Peerajan, S.; Chaiyasut, K.; Kesika, P. Anthocyanin profile and its antioxidant activity of widely used fruits, vegetables, and flowers in Thailand Asian. J. Pharm. Clin. Res. 2016, 9, 218–224. [Google Scholar]
- Ogawa, K.; Sakakibara, H.; Iwata, R.; Ishii, T.; Sato, T.; Goda, T.; Shimoi, K.; Kumazawa, S. Anthocyanin Composition and Antioxidant Activity of the Crowberry (Empetrum nigrum) and Other Berries. J. Agric. Food Chem. 2008, 56, 4457–4462. [Google Scholar] [CrossRef] [PubMed]
- Zykin, P.A.; Andreeva, E.A.; Lykholay, A.N.; Tsvetkova, N.V.; Voylokov, A.V. Anthocyanin Composition and Content in Rye Plants with Different Grain Color. Molecules 2018, 23, 948. [Google Scholar] [CrossRef] [PubMed]
- De Morais, J.S.; Sant’Ana, A.S.; Dantas, A.M.; Silva, B.S.; Lima, M.S.; Borges, G.C.; Magnani, M. Antioxidant activity and bioaccessibility of phenolic compounds in white, red, blue, purple, yellow and orange edible flowers through a simulated intestinal barrier. Food Res. Int. 2020, 131, 109046. [Google Scholar] [CrossRef] [PubMed]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef]
- Tamura, H.; Yamagami, A. Antioxidative activity of monoacylated anthocyanins isolated from Muscat Bailey A grape. J. Agric. Food Chem. 1994, 42, 1612–1615. [Google Scholar] [CrossRef]
- Stintzing, F.C.; Stintzing, A.S.; Carle, R.; Frei, B.; Wrolstad, R.E. Color and antioxidant properties of cyanidin-based anthocyanin pigments. J. Agric. Food Chem. 2002, 50, 6172–6181. [Google Scholar] [CrossRef]
- Tsuda, T.; Shiga, K.; Ohshima, K.; Kawakishi, S.; Osawa, T. Inhibition of lipid peroxidation and the active oxygen radical scavenging effect of anthocyanin pigments isolated from Phaseolus vulgaris L. Biochem. Pharmacol. 1996, 52, 1033–1110. [Google Scholar] [CrossRef]
- Brown, J.E.; Kelly, M.F. Inhibition of lipid peroxidation by anthocyanins, anthocyanidins and their phenolic degradation products. Eur. J. Lipid Sci. Technol. 2007, 109, 66–71. [Google Scholar] [CrossRef]
- Lapidot, T.; Harel, S.; Akiri, B.; Granit, R.; Kanner, J. pH-dependent forms of red wine anthocyanins as antioxidants. J. Agric. Food Chem. 1999, 47, 67–70. [Google Scholar] [CrossRef]
- Muselík, J.; García-Alonso, M.; Martín-López, M.P.; Žemlička, M.; Rivas-Gonzalo, J.C. Measurement of antioxidant activity of wine catechins, procyanidins, anthocyanins and pyranoanthocyanins. Int. J. Mol. Sci. 2007, 8, 797–809. [Google Scholar] [CrossRef]
- Jung, Y.K.; Joo, K.S.; Rho, S.J.; Kim, Y.R. pH-dependent antioxidant stability of black rice anthocyanin complexed with cycloamylose. LWT Food Sci. Technol. 2020, 129, 109474. [Google Scholar] [CrossRef]
- Sui, X.; Dong, X.; Zhou, W. Combined effect of pH and high temperature on the stability and antioxidant capacity of two anthocyanins in aqueous solution. Food Chem. 2014, 163, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Chakraborty, R.; Raychaudhuri, U. Determination of pH-dependent antioxidant activity of palm (Borassus flabellifer) polyphenol compounds by photoluminol and DPPH methods: A comparison of redox reaction sensitivity. 3 Biotech 2015, 5, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Março, P.H.; Poppi, R.J.; Scarminio, I.S.; Tauler, R. Investigation of the pH effect and UV radiation on kinetic degradation of anthocyanin mixtures extracted from Hibiscus acetosella. Food Chem. 2011, 125, 1020–1102. [Google Scholar]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Remesy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005, 81, 230–242. [Google Scholar] [CrossRef]
- Felgines, C.; Talavéra, S.; Texier, O.; Gil-Izquierdo, A.; Lamaison, J.-L.; Remesy, C. Blackberry anthocyanins are mainly recovered from urine as methylated and glucuronidated conjugates in humans. J. Agric. Food Chem. 2005, 53, 7721–7727. [Google Scholar] [CrossRef]
- Passamonti, S.; Vanzo, A.; Vrhovsek, U.; Terdoslavich, M.; Cocolo, A.; Decorti, G.; Mattivi, F. Hepatic uptake of grape anthocyanins and the role of bilitranslocase. Food Res. Int. 2005, 38, 953–960. [Google Scholar] [CrossRef]
- Sandoval-Ramírez, B.A.; Catalanń, U.; Fernanńdez-Castillejo, S.; Rubió, L.; Maciá, A.; Sola, R. Anthocyanin tissue bioavailability in animals: Possible implications for human health. A systematic review. J. Agric. Food Chem. 2018, 66, 11531–11543. [Google Scholar] [CrossRef]
- Lucioli, S. Anthocyanins: Mechanism of action and therapeutic efficacy. In Medicinal Plants as Antioxidant Agents: Understanding Their Mechanism of Action and Therapeutic Efficacy; Capasso, A., Ed.; Research Signpost: Kerala, India, 2012; pp. 27–57. [Google Scholar]
- Jin, Y.A.D.; George, T.G.M.; Lovegrove, J.A.A. Randomised trial to investigate the effects of acute consumption of a blackcurrant juice drink on markers of vascular reactivity and bioavailability of anthocyanins in human subjects. Eur. J. Clin. Nutr. 2011, 65, 849–857. [Google Scholar] [CrossRef]
- Wu, X.; Cao, G.; Prior, R.L. Absorption and metabolism of anthocyanins in elderly women after consumption of elderberry or blueberry. J. Nutr. 2002, 132, 1865–1871. [Google Scholar] [CrossRef] [PubMed]
- Williamson, G.; Clifford, M.N. Colonic metabolites of berry polyphenols: The missing link to biological activity? Br. J. Nutr. 2010, 104 (Suppl. S3), S48–S66. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, M.; Müller-Ehl, L.; Passon, M.; Schieber, A. Development and validation of methods for the determination of anthocyanins in physiological fluids via UHPLC-MSn. Molecules 2020, 25, 518. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, I.A.; Mena, P.; Calani, L.; Borges, G.; Pereira-Caro, G.; Bresciani, L.; Del Rio, D.; Lean, M.E.J.; Crozier, A. New insights into the bioavailability of red raspberry anthocyanins and ellagitannins. Free Radic. Biol. Med. 2015, 89, 758–769. [Google Scholar] [CrossRef]
- Ferrars, R.M.; Cassidy, A.; Curtis, P.; Kay, C.D. Phenolic metabolites of anthocyanins following a dietary intervention study in post-menopausal women. Mol. Nutr. Food Res. 2014, 58, 490–502. [Google Scholar] [CrossRef]
- Ferrars, R.M.; Czank, C.; Saha, S.; Needs, P.W.; Zhang, Q.; Raheem, K.S.; Botting, N.P.; Kroon, P.A.; Kay, C.D. Methods for isolating, identifying, and quantifying anthocyanin metabolites in clinical samples. Anal. Chem. 2014, 86, 10052–10058. [Google Scholar] [CrossRef]
- Nurmi, T.; Mursu, J.; Heinonen, M.; Nurmi, A.; Hiltunen, R.; Voutilainen, S. Metabolism of berry anthocyanins to phenolic acids in humans. J. Agric. Food Chem. 2009, 57, 2274–2281. [Google Scholar] [CrossRef]
- Czank, C.; Cassidy, A.; Zhang, Q.; Morrison, D.J.; Preston, T.; Kroon, P.A.; Botting, N.P.; Kay, C.D. Human metabolism and elimination of the anthocyanin, cyanidin-3-glucoside: A 13C-tracer study. Am. J. Clin. Nutr. 2013, 97, 995–1003. [Google Scholar] [CrossRef]
- Novotny, J.A. Anthocyanin Bioavailability: Past Progress and Current Challenges. In Emerging Trends in Dietary Components for Preventing and Combating Disease; ACS Symposium Series; American Chemical Society: Washington, DC, USA, 2012; Chapter 32; pp. 559–568. [Google Scholar]
- Kim, I.; Moon, J.K.; Hur, S.J.; Lee, J. Structural changes in mulberry (Morus Microphylla. Buckl) and chokeberry (Aronia melanocarpa) anthocyanins during simulated in vitro human digestion. Food Chem. 2020, 318, 126449. [Google Scholar] [CrossRef]
- Gowd, V.; Karim, N.; Xie, L.; Shishir, M.R.I.; Xu, Y.; Chen, W. In vitro study of bioaccessibility, antioxidant, and α-glucosidase inhibitory effect of pelargonidin-3-O-glucoside after interacting with beta-lactoglobulin and chitosan/pectin. Int. J. Biol. Macromol. 2020, 154, 380–389. [Google Scholar] [CrossRef]
- Gowd, V.; Bao, T.; Chen, W. Antioxidant potential and phenolic profile of blackberry anthocyanin extract followed by human gut microbiota fermentation. Food Res. Int. 2019, 120, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Xiec, M.; Yanga, F.; Liu, J. Antioxidant activity of high purity blueberry anthocyanins and the effects on human intestinal microbiota. Food Sci. Technol. 2020, 117, 628101. [Google Scholar] [CrossRef]
- Zhu, Y.; Sun, H.; He, S.; Lou, Q.; Yu, M.; Tang, M.; Tu, L. Metabolism and prebiotics activity of anthocyanins from black rice (Oryza sativa L.) in vitro. PLoS ONE 2018, 13, 0195754. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, M.; Oruna-Concha, M.J.; Kolida, S.; Walton, G.E.; Kallithraka, S.; Spencer, J.P.E.; Gibson, G.R.; de Pascual-Teresa, S. Metabolism of anthocyanins by human gut microflora and their influence on gut bacterial growth. J. Agric. Food Chem. 2012, 60, 3882–3890. [Google Scholar] [CrossRef]
- Boto-Ordóñez, M.; Urpi-Sarda, M.; Queipo-Ortuño, M.I.; Tulipani, S.; Tinahones, F.J.; Andres-Lacueva, C. High levels of Bifidobacteria are associated with increased levels of anthocyanin microbial metabolites: A randomized clinical trial. Food Funct. 2014, 5, 1932–1938. [Google Scholar] [CrossRef]
- Garcia-Mazcorro, J.F.; Lage, N.N.; Mertens-Talcott, S.; Talcott, S.; Chew, B.; Dowd, S.E.; Kawas, J.R.; Noratto, G.D. Effect of dark sweet cherry powder consumption on the gut microbiota, short-chain fatty acids, and biomarkers of gut health in obese db/db mice. Peer J. 2018, 2018, 1–31. [Google Scholar] [CrossRef]
- Lacombe, A.; Li, R.W.; Klimis-Zacas, D.; Kristo, A.S.; Tadepalli, S.; Krauss, E.; Young, R.; Wu, V.C.H. Lowbush wild blueberries have the potential to modify gut microbiota and xenobiotic metabolism in the rat colon. PLoS ONE 2013, 8, 67497. [Google Scholar] [CrossRef]
- Flores, G.; Ruiz del Castillo, M.L.; Costabile, A.; Klee, A.; Bigetti Guergoletto, K.; Gibson, G.R. In vitro fermentation of anthocyanins encapsulated with cyclodextrins: Release, metabolism and influence on gut microbiota growth. J. Funct. Foods 2015, 16, 50–57. [Google Scholar] [CrossRef]
- Guergoletto, K.B.; Costabile, A.; Flores, G.; Garcia, S.; Gibson, G.R. In vitro fermentation of juçara pulp (Euterpe edulis) by human colonic microbiota. Food Chem. 2016, 196, 251–258. [Google Scholar] [CrossRef]
- Wu, Y.; Han, Y.; Tao, Y.; Li, D.; Xie, G.; Show, P.L.; Lee, S.Y. In vitro gastrointestinal digestion and fecal fermentation reveal the effect of different encapsulation materials on the release, degradation and modulation of gut microbiota of blueberry anthocyanin extract. Food Res. Int. 2020, 132, 109098. [Google Scholar] [CrossRef]
- Sharif, N.; Khoshnoudi-Nia, S.; Jafari, S.M. Nano/microencapsulation of anthocyanins; a systematic review and meta-analysis. Food Res. Int. 2020, 132, 109077. [Google Scholar] [CrossRef] [PubMed]
- Yousuf, B.; Gul, K.; Wani, A.A.; Singh, P. Health benefits of anthocyanins and their encapsulation for potential use in food systems: A review. Crit. Rev. Food Sci. Nutr. 2016, 56, 2223–2230. [Google Scholar] [CrossRef] [PubMed]
- Mahdavi, S.A.; Jafari, S.M.; Ghorbani, M.; Assadpoor, E. Spray-drying microencapsulation of anthocyanins by natural biopolymers: A review. Dry Technol. 2014, 32, 509–518. [Google Scholar] [CrossRef]
- Robert, P.; Fredes, C. The encapsulation of anthocyanins from berry-type fruits. Trends in foods. Molecules 2015, 20, 5875–5888. [Google Scholar] [CrossRef]
- Cavalcanti, R.N.; Santos, D.T.; Meireles, M.A.A. Non-thermal stabilization mechanisms of anthocyanins in model and food systems—An overview. Food Res. Int. 2011, 44, 499–509. [Google Scholar] [CrossRef]
- Fang, Z.; Bhandari, B. Encapsulation of polyphenols—A review. Trends Food Sci. Technol. 2010, 21, 510–523. [Google Scholar] [CrossRef]
- Fernandes, A.; Rocha, M.; Santos, L.; Brás, J.; Oliveira, J.; Mateus, N.; de Freitas, V. Blackberry anthocyanins: β-Cyclodextrin fortification for thermal and gastro- intestinal stabilization. Food Chem. 2018, 245, 426–431. [Google Scholar] [CrossRef]
- Huang, Y.; Zhou, W.B. Microencapsulation of anthocyanins through two-step emulsification and release characteristics during in vitro digestion. Food Chem. 2019, 278, 357–363. [Google Scholar] [CrossRef]
- De Pascual-Teresa, S.; Sanchez-Ballesta, M.T. Anthocyanins: From plant to health. Phytochem. Rev. 2008, 7, 281–299. [Google Scholar] [CrossRef]
- Clifford, M.N. Anthocyanins—Nature, occurrence and dietary sources. J. Sci. Food Agric. 2000, 80, 1063–1072. [Google Scholar] [CrossRef]
- Shim, S.H.; Kim, J.M.; Choi, C.Y.; Kim, C.Y.; Park, K.H. Ginkgo biloba extract and bilberry anthocyanins improve visual function in patients with normal tension glaucoma. J. Med. Food 2012, 15, 818–823. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Kaleem, M.; Ahmed, Z.; Shafiq, H. Therapeutic potential of flavonoids and their mechanism of action against microbial and viral infections—A review. Food Res. Int. 2015, 77, 221–235. [Google Scholar] [CrossRef]
- Miyake, S.; Takahashi, N.; Sasaki, M.; Takahashi, N.; Ozawa, Y. Vision preservation during retinal inflammation by anthocyanin- rich bilberry extract: Cellular and molecular mechanism. Lab. Investig. 2012, 92, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Lee, H.K.; Kim, C.Y.; Hong, Y.J.; Choe, C.M.; You, T.W.; Seong, G.J. Purified high-dose anthocyanoside oligomer administration improves nocturnal vision and clinical symptoms in myopia subjects. Br. J. Nutr. 2005, 93, 895–899. [Google Scholar] [CrossRef]
- Thiraphatthanavong, P.; Wattanathorn, J.; Muchimapura, S.; Wipawee, T.M.; Wannanon, P.; Terdthai, T.U.; Suriharn, B.; Lertrat, K. Preventive effect of Zea mays L. (purple waxy corn) on experimental diabetic cataract. BioMed Res. Int. 2014, 2014, 507435. [Google Scholar] [CrossRef]
- Mok, J.W.; Chang, D.J.; Joo, C.K. Antiapoptotic effects of anthocyanin from the seed coat of black soybean against oxidative damage of human lens epithelial cell induced by H2O2. Curr. Eye Res. 2014, 39, 1090–1098. [Google Scholar] [CrossRef]
- Paik, S.S.; Jeong, E.; Jung, S.W.; Ha, T.J.; Kang, S.; Sim, S.; Jeon, J.H.; Chun, M.H.; Kim, I.B. Anthocyanins from the seed coat of black soybean reduce retinal degeneration induced by N-methyl-N-nitrosourea. Exp. Eye Res. 2012, 97, 55–62. [Google Scholar] [CrossRef]
- Ohguro, H.; Ohguro, I.; Katai, M.; Tanaka, S. Two-year randomized, placebo-controlled study of black currant anthocyanins on visual field in glaucoma. Ophthalmologica 2012, 228, 26–35. [Google Scholar] [CrossRef]
- Yang, Y.; Shi, Z.; Reheman, A.; Jin, W.; Li, C.; Zhu, G.; Wang, Y.; Freedman, J.J.; Ling, W.; Ni, H. Anthocyanins inhibit platelet activation and attenuate thrombus growth in both human and murine thrombosis models. Blood 2010, 116, 3197. [Google Scholar] [CrossRef]
- Qin, Y.; Xia, M.; Ma, J.; Hao, Y.; Liu, J.; Mou, H.; Cao, L.; Ling, W. Anthocyanin supplementation improves serum LDL-and HDL-cholesterol concentrations associated with the inhibition of cholesteryl ester transfer protein in dyslipidemic subjects. Am. J. Clin. Nutr. 2009, 90, 485–492. [Google Scholar] [CrossRef]
- Cassidy, A.; Bertoia, M.; Chiuve, S.; Flint, A.; Forman, J.; Rimm, E.B. Habitual intake of anthocyanins and flavanones and risk of cardiovascular disease in men. Am. J. Clin. Nutr. 2016, 104, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Bell, D.R.; Gochenaur, K. Direct vasoactive and vasoprotective properties of anthocyanin-rich extracts. J. Appl. Physiol. 2006, 100, 1164–1170. [Google Scholar] [CrossRef] [PubMed]
- Toufektsian, M.C.; De Lorgeril, M.; Nagy, N.; Salen, P.; Donati, M.B.; Giordano, L.; Mock, H.-P.; Peterek, S.; Matros, A.; Petroni., K.; et al. Chronic dietary intake of plant-derived anthocyanins protects the rat heart against ischemia-reperfusion injury. J. Nutr. 2008, 138, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Suarez, J.M.; Giampieri, F.; Tulipani, S.; Casoli, T.; di Stefano, G. One month strawberry-rich anthocyanin supplementation ameliorates cardiovascular risk, oxidative stress markers and platelet activation in humans. J. Nutr. Biochem. 2014, 25, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.H.; Ahn, I.S.; Kim, S.O.; Kong, C.S.; Chung, H.Y.; Do, M.S.; Park, K.Y. Anti-obesity and hypolipidemic effects of black soybean anthocyanins. J. Med. Food 2007, 10, 552–556. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Yu, Z.; Tang, Q.; Song, H.; Gao, Z.; Chen, W.; Zheng, X. Honeysuckle anthocyanin supplementation prevents diet-induced obesity in C57BL/6 mice. Food Funct. 2013, 4, 1654–1661. [Google Scholar] [CrossRef] [PubMed]
- Jankowski, A.; Jankowska, B.; Niedworok, J. The effects of anthocyanin dye from grapes on experimental diabetes. Folia Med. Cracov. 2000, 41, 5–15. [Google Scholar]
- Jayaprakasam, B.; Vareed, S.K.; Olson, L.K.; Nair, M.G. Insulin secretion by bioactive anthocyanins and anthocyanidins present in fruits. J. Agric. Food Chem. 2005, 53, 28–31. [Google Scholar] [CrossRef]
- Tsuda, T.; Ueno, Y.; Aoki, H.; Koda, T.; Horio, F.; Takahashi, N.; Kawada, T.; Osawa, T. Anthocyanin enhances adipocytokine secretion and adipocyte-specific gene expression in isolated rat adipocytes. Biochem. Biophys. Res. Commun. 2004, 316, 149–157. [Google Scholar] [CrossRef]
- Tsuda, T.; Horio, F.; Uchida, K.; Aoki, H.; Osawa, T. Dietary cyanidin 3- O-β-D-glucoside-rich purple corn color prevents obesity and ameliorates hyperglycemia in mice. J. Nutr. 2003, 133, 2125–2130. [Google Scholar] [CrossRef]
- Takikawa, M.; Inoue, S.; Horio, F.; Tsuda, T. Dietary anthocyanin-rich bilberry extract ameliorates hyperglycemia and insulin sensitivity via activation of AMP-activated protein kinase in diabetic mice. J. Nutr. 2010, 140, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhang, Y.; Liu, Y.; Sun, R.; Xia, M. Purified anthocyanin supplementation reduces dyslipidemia, enhances antioxidant capacity, and prevents insulin resistance in diabetic patients. J. Nutr. 2015, 145, 742–748. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.K.; Lim, S.S.; Lee, J.Y.; Yeo, K.M.; Kang, Y.-H. Anthocyanin-rich purple corn extract inhibit diabetes-associated glomerular angiogenesis. PLoS ONE 2013, 8, 79823. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Johnson, M.H.; Lila, M.A.; Yousef, G.; de Mejia, E.G. Berry and citrus phenolic compounds inhibit dipeptidyl peptidase IV: Implications in diabetes management. Evid. Based Complement. Alternat. Med. 2013, 2013, 479505. [Google Scholar] [CrossRef]
- Koh, E.S.; Lim, J.H.; Kim, M.Y.; Chung, S.; Shin, S.J.; Choi, B.S.; Kim, H.W.; Hwang, S.Y.; Kim, S.W.; Park, C.W.; et al. Anthocyanin-rich Seoritae extract ameliorates renal lipotoxicity via activation of AMP-activated protein kinase in diabetic mice. J. Transl. Med. 2015, 13, 203. [Google Scholar] [CrossRef]
- Liu, Y.; Li, D.; Zhang, Y.; Sun, R.; Xia, M. Anthocyanin increases adiponectin secretion and protects against diabetes-related endothelial dysfunction. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E975–E988. [Google Scholar] [CrossRef]
- Pojer, E.; Mattivi, F.; Johnson, D.; Stockley, C.S. The case for anthocyanin consumption to promote human health: A review. Compr. Rev. Food Sci. Food Saf. 2013, 12, 483–508. [Google Scholar] [CrossRef]
- Genskowsky, E.; Puente, L.A.; Perez-Alvarez, J.A.; Fernández-López, J.; Muñoz, L.A.; Viuda-Martos, M. Determination of polyphenolic profile, antioxidant activity and antibacterial properties of maqui [Aristotelia chilensis (Molina) Stuntz] a Chilean blackberry. J. Sci. Food Agr. 2016, 96, 4235–4242. [Google Scholar] [CrossRef]
- Côté, J.; Caillet, S.; Doyon, G.; Ng, K. Antimicrobial effect of cranberry juice and extracts. Food Cont. 2011, 22, 1413–1418. [Google Scholar] [CrossRef]
- Puupponen-Pimiä, R.; Nohynek, L.; Meier, C.; Kähkönen, M.; Heinonen, M.; Hopia, A.; Oksman-Caldentey, K.M. Antimicrobial properties of phenolic compounds from berries. J. Appl. Microbiol. 2001, 90, 494–507. [Google Scholar] [CrossRef]
- Wang, L.S.; Hecht, S.S.; Carmella, S.G.; Yu, N.; Larue, B.; Henry, C.; McIntyre, C.; Rocha, C.; Lechner, J.F.; Stoner, G.D. Anthocyanins in black raspberries prevent esophageal tumors in rats. Cancer Prev. Res. 2009, 2, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Faria, A.; Pestana, D.; Teixeira, D.; de Freitas, V.; Mateus, N.; Calhau, C. Blueberry anthocyanins and pyruvic acid adducts: Anticancer properties in breast cancer cell lines. Phytother. Res. 2010, 24, 1862–1869. [Google Scholar] [CrossRef]
- Hui, C.; Bin, Y.; Xiaoping, Y.; Long, Y.; Chunye, C.; Mantian, M.; Wenhua, L. Anticancer activities of an anthocyanin-rich extract from black rice against breast cancer cells in vitro and in vivo. Nutr. Cancer 2010, 62, 1128–1136. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Y.; Zhou, J.; Luo, L.P.; Han, B.; Li, F.; Chen, J.Y.; Zhu, Y.F.; Chen, W.; Yu, X.P. Black rice anthocyanins suppress metastasis of breast cancer cells by targeting RAS/RAF/MAPK pathway. BioMed Res. Int. 2015, 2015, 414250. [Google Scholar] [CrossRef] [PubMed]
- Malik, M.; Zhao, C.; Schoene, N.; Guisti, M.M.; Moyer, M.P.; Magnuson, B.A. Anthocyanin-rich extract from Aronia meloncarpa E. induces a cell cycle block in colon cancer but not normal colonic cells. Nutr. Cancer 2003, 46, 186–196. [Google Scholar] [CrossRef] [PubMed]
- Lala, G.; Malik, M.; Zhao, C.; He, J.; Kwon, Y.; Giusti, M.M.; Magnuson, B.A. Anthocyanin-rich extracts inhibit multiple biomarkers of colon cancer in rats. Nutr. Cancer 2006, 54, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.; Xu, J.; Kim, J.; Chen, T.Y.; Su, X.; Standard, J.; Carey, E.; Griffin, J.; Herndon, B.; Katz, B.; et al. Role of anthocyanin-enriched purple-fleshed sweet potato p40 in colorectal cancer prevention. Mol. Nutr. Food Res. 2013, 57, 1908–1917. [Google Scholar] [CrossRef]
- Jang, H.; Ha, U.S.; Kim, S.J.; Yoon, B.I.; Han, D.S.; Yuk, S.M.; Kim, S.W. Anthocyanin extracted from black soybean reduces prostate weight and promotes apoptosis in the prostatic hyperplasia-induced rat model. J. Agric. Food Chem. 2010, 58, 12686–12691. [Google Scholar] [CrossRef]
- Shin, D.Y.; Lee, W.S.; Kim, S.H.; Kim, M.J.; Yun, J.W.; Lu, J.N.; Lee, S.J.; Tsoy, I.; Kim, H.J.; Ryu, C.H.; et al. Anti-invasive activity of anthocyanins isolated from Vitis coignetiae in human hepatocarcinoma cells. J. Med. Food. 2009, 12, 967–972. [Google Scholar] [CrossRef]
- Bontempo, P.; de Masi, L.; Carafa, V.; Rigano, D.; Scisciola, L.; Iside, C.; Grassi, R.; Molinari, A.M.; Aversano, R.; Nebbioso, A.; et al. Anticancer activities of anthocyanin extract from genotyped Solanum tuberosum L. “Vitelotte”. J. Funct. Foods 2015, 19, 584–593. [Google Scholar] [CrossRef]
- Strathearn, K.E.; Yousef, G.G.; Grace, M.H.; Roy, S.L.; Tambe, M.A.; Ferruzzi, M.G.; Wu, Q.-L.; Simon, J.E.; Lila, M.A.; Rochet, J.-C. Neuroprotective effects of anthocyanin and proanthocyanidin-rich extracts in cellular models of Parkinson’s disease. Brain Res. 2014, 1555, 60–77. [Google Scholar] [CrossRef] [PubMed]
- Shih, P.-H.; Chan, Y.-C.; Liao, J.-W.; Wang, M.-F.; Yen, G.-C. Antioxidant and cognitive promotion effects of anthocyanin-rich mulberry (Morus atropurpurea L.) on senescence-accelerated mice and prevention of Alzheimer’s disease. J. Nutr. Biochem. 2010, 21, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Gutierres, J.M.; Carvalho, F.B.; Schetinger, M.R.C.; Agostinho, P.; Marisco, P.C. Neuroprotective effect of anthocyanins on acetylcholinesterase activity and attenuation of scopolamine-induced amnesia in rats. Int. J. Dev. Neurosci. 2014, 33, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Rehman, S.U.; Shah, S.A.; Ali, T.; Chung, J.I.; Kim, M.O. Anthocyanins reversed D-galactose-induced oxidative stress and neuroinflammation mediated cognitive impairment in adult rats. Mol. Neurobiol. 2017, 54, 255–271. [Google Scholar] [CrossRef] [PubMed]
- Amin, F.U.; Shah, S.A.; Badshah, H.; Khan, M.; Kim, M.O. Anthocyanins encapsulated by PLGA@PEG nanoparticles potentially improved its free radical scavenging capabilities via p38/JNK pathway against Abeta1-42-induced oxidative stress. J. Nanobiotechnol. 2017, 15, 12. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Rehman, S.U.; Amin, F.U.; Kim, M.O. Enhanced neuroprotection of anthocyanin-loaded PEG-gold nanoparticles against Abeta1-42-induced neuroinflammation and neurodegeneration via the NF-KB /JNK/GSK3beta signaling pathway. Nanomedicine 2017, 13, 2533–2544. [Google Scholar] [CrossRef]
- Ali, T.; Kim, M.J.; Rehman, S.U.; Ahmad, A.; Kim, M.O. Anthocyanin-Loaded PEG-Gold nanoparticles enhanced the neuroprotection of anthocyanins in an Abeta1-42 mouse model of Alzheimer’s disease. Mol. Neurobiol. 2017, 54, 6490–6506. [Google Scholar] [CrossRef]
- Nomi, Y.; Iwasaki-Kurashige, K.; Matsumoto, H. Therapeutic effects of anthocyanins for vision and eye health. Molecules 2019, 24, 3311. [Google Scholar] [CrossRef]
- Ghosh, D.; Konishi, T. Anthocyanins and anthocyanin-rich extracts: Role in diabetes and eye function. Asian Pac. J. Clin. Nutr. 2007, 16, 200–208. [Google Scholar]
- Tsuda, T. Anthocyanins as functional food factors: Chemistry, nutrition and health promotion. Food Sci. Technol. Res. 2012, 18, 315–324. [Google Scholar] [CrossRef]
- Smeriglio, A.; Monteleone, D.; Trombetta, D. Health effects of Vaccinium myrtillus L.: Evaluation of efficacy and technological strategies for preservation of active ingredients. Mini Rev. Med. Chem. 2014, 14, 567–584. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.K.; Lee, H.J.; Shih, Y.W.; Chyau, C.C.; Wang, C.J. Mulberry anthocyanins extracts inhibit LDL oxidation and macrophage-derived foam cell formation induced by oxidative LDL. J. Food Sci. 2008, 73, 4113–4121. [Google Scholar] [CrossRef] [PubMed]
- Rechner, A.R.; Kroner, C. Anthocyanins and colonic metabolites of dietary polyphenols inhibit platelet function. Thromb. Res. 2005, 116, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Luís, Â.; Domingues, F.; Pereira, L. Association between berries intake and cardiovascular diseases risk factors: A systematic review with meta-analysis and trial sequential analysis of randomized controlled trials. Food Funct. 2018, 9, 740–757. [Google Scholar] [CrossRef] [PubMed]
- García-Conesa, M.T.; Chambers, K.; Combet, E.; Pinto, P.; Garcia-Aloy, M.; Andresń-Lacueva, C.; dePascual-Teresa, S.; Mena, P.; Konic Ristic, A.; Hollands, W.J.; et al. Meta-Analysis of the effects of foods and derived products containing ellagitannins and anthocyanins on cardiometabolic biomarkers: Analysis of factors influencing variability of the individual responses. Int. J. Mol. Sci. 2018, 19, 694. [Google Scholar] [CrossRef] [PubMed]
- Ksonzékova, P.; Mariychuk, R.; Eliasova, A.; Mudronova, D.; Csank, T.; Kiraly, J.; Marcincakova, D.; Pistl, J.; Tkacikova, L. In vitro studies of biological activities of anthocyanin-rich berry extracts on porcine intestinal epithelial cells. J. Sci. Food Agric. 2016, 96, 1093–1100. [Google Scholar] [CrossRef]
- Bone, K.; Mills, S. Principles and Practice of Phytotherapy: Modern Herbal Medicine; Churchill Livingstone: Edinburg, TX, USA, 2013. [Google Scholar]
- Xie, L.; Su, H.; Sun, C.; Zheng, X.; Chen, W. Recent advances in understanding the anti-obesity activity of anthocyanins and their biosynthesis in microorganisms. Trends Food Sci. Technol. 2018, 72, 13–24. [Google Scholar] [CrossRef]
- Rupasinghe, H.P.V.; Arumuggam, N. Health benefits of anthocyanins. Food Chem. Func. Anal. 2019, 2019, 123–158. [Google Scholar]
- Heyman, L.; Axling, U.; Blanco, N.; Sterner, O.; Holm, C.; Berger, K. Evaluation of beneficial metabolic effects of berries in high-fat fed C57BL/6J mice. J. Nutr. Metab. 2014, 2014, 403041. [Google Scholar] [CrossRef]
- Lachin, T.; Reza, H. Anti diabetic effect of cherries in alloxan induced diabetic rats. Recent Pat. Endocr. Metab. Immune Drug Discov. 2012, 6, 67–72. [Google Scholar] [CrossRef]
- Orqueda, M.E.; Torres, S.; Zampini, I.C.; Cattaneo, F.; Fernandez Di Pardo, A.; Valle, E.M.; Jimenez-Aspee, F.; Schmeda-Hirschmann, G.; Isla, M.I. Integral use of Argentinean Solanum betaceum red fruits as functional food ingredient to prevent metabolic syndrome: Effect of in vitro simulated gastroduodenal digestión. Heliyon 2020, 6, 03387. [Google Scholar] [CrossRef] [PubMed]
- Wedick, N.M.; Pan, A.; Cassidy, A.; Rimm, E.B.; Sampson, L.; Rosner, B.; Willett, W.; Hu, F.B.; Sun, Q.; van Dam, R.M. Dietary flavonoid intakes and risk of type 2 diabetes in US men and women. Am. J. Clin. Nutr. 2012, 95, 925–933. [Google Scholar] [CrossRef] [PubMed]
- Cisowska, A.; Wojnicz, D.; Hendrich, A.B. Anthocyanins as antimicrobial agents of natural plant origin. Nat. Prod. Commun. 2011, 6, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Helander, I.M.; Alakomi, H.L.; Latva-Kala, K.; Mattila-Sandholm, T.; Pol, I.; Smid, E.J.; Gorris, L.G.M.; von Wright, A. Characterization of the action of selected essential oil components on Gram-negative bacteria. J. Agric. Food Chem. 1998, 46, 3590–3595. [Google Scholar] [CrossRef]
- Riaz, M.; Zia-Ul-Haq, M.; Saad, B. Anthocyanins and Human Health: Biomolecular and Therapeutical Aspects; Springer: New York, NY, USA, 2016. [Google Scholar]
- Wallace, T.C.; Giusti, M.M. Anthocyanins in Health and Disease; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar]
- Fraga, C.G. (Ed.) Plant Phenolics and Human Health: Biochemistry, Nutrition and Pharmacology; Wiley: New York, NY, USA, 2010. [Google Scholar]
- He, J.; Giusti, M.M. Anthocyanins: Natural colorants with health-promoting properties. Annu. Rev. Food Sci. Technol. 2010, 1, 163–187. [Google Scholar] [CrossRef]
- Galvano, F.; Salamone, F.; Nicolosi, A.; Vitaglione, P. Anthocyanins-based drugs for colon cancer treatment: The nutriotionist’s point of view. Cancer Chemother. Pharmacol. 2009, 64, 431–432. [Google Scholar] [CrossRef]
- Webb, M.R.; Min, K.; Ebeler, S.E. Anthocyanins interactions with DNA: Intercalation, topoisomerase I inhibition and oxidative reactions. J. Food Biochem. 2008, 32, 176–196. [Google Scholar] [CrossRef]
- Fernandes, I.; Faria, A.; Azevedo, J.; Soares, S.; Calhau, C.; De Freitas, V.; Mateus, N. Influence of anthocyanins, derivative pigments and other catechol and pyrogallol-type phenolics on breast cancer cell proliferation. J. Agric. Food Chem. 2010, 58, 3785–3792. [Google Scholar] [CrossRef]
- Sun, J.; Hai Liu, R. Cranberry phytochemical extracts induce cell cycle arrest and apoptosis in human MCF-7 breast cancer cells. Cancer Lett. 2006, 241, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Hoshyar, R.; Mahboob, Z.; Zarban, A. The antioxidant and chemical properties of Berberis vulgaris and its cytotoxic effect on human breast carcinoma cells. Cytotechnology 2015, 68, 1207–1213. [Google Scholar] [CrossRef]
- Im, N.K.; Jang, W.J.; Jeong, C.H.; Jeong, G.S. Delphinidin suppresses PMA-induced MMP-9 expression by blocking the NF-kappaB activation through MAPK signaling pathways in MCF-7 human breast carcinoma cells. J. Med. Food. 2014, 17, 855–861. [Google Scholar] [CrossRef] [PubMed]
- Forester, S.C.; Choy, Y.Y.; Waterhouse, A.L.; Oteiza, P.I. The anthocyanin metabolites gallic acid, 3-O-methylgallic acid, and 2,4,6- trihydroxybenzaldehyde decrease human colon cancer cell viability by regulating pro-oncogenic signals. Mol. Carcinog. 2014, 53, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.N.; Kuo, W.H.; Chiang, C.L.; Chiou, H.L.; Hsieh, Y.S.; Chu, S.C. Black rice anthocyanins inhibit cancer cells invasion via repressions of MMPs and u-PA expression. Chem. Biol. Interact. 2006, 163, 218–229. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wang, P.; Luo, Y.; Zhao, M.; Chen, F. Health benefits of anthocyanins and molecular mechanisms: Update from recent decade. Crit. Rev. Food Sci. Nutr. 2017, 57, 1729–1741. [Google Scholar] [CrossRef] [PubMed]
- Jing, P.; Bomser, J.A.; Schwartz, S.J.; He, J.; Magnuson, B.A.; Giusti, M.M. Structure–function relationships of anthocyanins from various anthocyanin-rich extracts on the inhibition of colon cancer cell growth. J. Agric. Food Chem. 2008, 56, 9391–9398. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Giusti, M.M.; Malik, M.; Moyer, M.P.; Magnuson, B.A. Effects of commercial anthocyanin-rich extracts on colonic cancer and nontumorigenic colonic cell growth. J. Agric. Food Chem. 2004, 52, 6122–6128. [Google Scholar] [CrossRef]
- Winter, A.N.; Bickford, P.C. Anthocyanins and their metabolites as therapeutic agents for neurodegenerative disease. Antioxidants 2019, 8, 333. [Google Scholar] [CrossRef]
- Klinkenberg, I.; Blokland, A. The validity of scopolamine as a pharmacological model for cognitive impairment: A review of animal behavioral studies. Neurosci. Biobehav. Rev. 2010, 34, 1307–1350. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).