Phytochemical Profile of Capsicum annuum L. cv Senise, Incorporation into Liposomes, and Evaluation of Cellular Antioxidant Activity

 ,
,  ,
,  and
and
Abstract
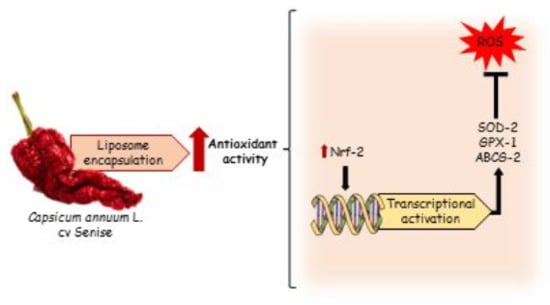
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Extract Preparation
2.3. LC-ESI/LTQOrbitrap/MS
2.4. Oxygen Radical Absorbance Capacity (ORAC) Assay
2.5. Liposome Preparation and Characterization
2.6. Cell Line and Culture Conditions
2.7. MTT Assay
2.8. Measurement of Intracellular Reactive Oxygen Species (ROS)
2.9. Quantitative RT-PCR
2.10. Statistical Analysis
3. Results
3.1. Phytochemical Profile of C. annuum Extract
3.2. Antioxidant Activity: ORAC Assay
3.3. Liposome Characterization
3.4. Effect of C. annuum Extract on Cell Viability and Intracellular ROS
3.5. Effect of C. annuum Extract on Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Flora, S. Role of free radicals and antioxidants in health and disease. Cell. Mol. Biol. 2007, 53, 1–2. [Google Scholar]
- Faraone, I.; Rai, D.K.; Russo, D.; Chiummiento, L.; Fernandez, E.; Choudhary, A.; Milella, L. Antioxidant, Antidiabetic, and Anticholinesterase Activities and Phytochemical Profile of Azorella glabra Wedd. Plants 2019, 8, 265. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Tsao, R. Dietary polyphenols, oxidative stress and antioxidant and anti-inflammatory effects. Curr. Opin. Food Sci. 2016, 8, 33–42. [Google Scholar] [CrossRef]
- Kaulmann, A.; Bohn, T. Carotenoids, inflammation, and oxidative stress—Implications of cellular signaling pathways and relation to chronic disease prevention. Nutr. Res. 2014, 34, 907–929. [Google Scholar] [CrossRef] [PubMed]
- Mignet, N.; Seguin, J.; Chabot, G.G. Bioavailability of Polyphenol Liposomes: A Challenge Ahead. Pharmaceutics 2013, 5, 457–471. [Google Scholar] [CrossRef]
- Bozzuto, G.; Molinari, A. Liposomes as nanomedical devices. Int. J. Nanomed. 2015, 10, 975–999. [Google Scholar] [CrossRef]
- Loizzo, M.R.; Pugliese, A.; Bonesi, M.; De Luca, D.; Menichini, F.; Tundis, R.; O’Brien, N. Influence of drying and cooking process on the phytochemical content, antioxidant and hypoglycaemic properties of two bell Capsicum annum L. cultivars. Food Chem. Toxicol. 2013, 53, 392–401. [Google Scholar] [CrossRef]
- Speranza, G.; Scalzo, R.L.; Morelli, C.F.; Rabuffetti, M.; Bianchi, G. Influence of drying techniques and growing location on the chemical composition of sweet pepper (Capsicum annuum L., var. Senise). J. Food Biochem. 2019, 43, e13031. [Google Scholar] [CrossRef]
- Moudache, M.; Colon, M.; Nerín, C.; Zaidi, F. Phenolic content and antioxidant activity of olive by-products and antioxidant film containing olive leaf extract. Food Chem. 2016, 212, 521–527. [Google Scholar] [CrossRef]
- Lima, C.F.; Fernandes-Ferreira, M.; Pereira-Wilson, C. Phenolic compounds protect HepG2 cells from oxidative damage: Relevance of glutathione levels. Life Sci. 2006, 79, 2056–2068. [Google Scholar] [CrossRef]
- Amorati, R.; Valgimigli, L. Advantages and limitations of common testing methods for antioxidants. Free. Radic. Res. 2015, 49, 633–649. [Google Scholar] [CrossRef] [PubMed]
- Aversano, R.; Contaldi, F.; Adelfi, M.G.; D’Amelia, V.; Diretto, G.; De Tommasi, N.; Vaccaro, C.; Vassallo, A.; Carputo, D. Comparative metabolite and genome analysis of tuber-bearing potato species. Phytochemsitry 2017, 137, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Tuberoso, C.; Serreli, G.; Montoro, P.; D’Urso, G.; Congiu, F.; Kowalczyk, A. Biogenic amines and other polar compounds in long aged oxidized Vernaccia di Oristano white wines. Food Res. Int. 2018, 111, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, A.; Anis, I.; Ali, Z.; Awadelkarim, S.; Khan, A.; Khalid, A.; Shah, M.R.; Galal, M.; Khan, I.A.; Choudhary, M.I. Methylenebissantin: A rare methylene-bridged bisflavonoid from Dodonaea viscosa which inhibits Plasmodium falciparum enoyl-ACP reductase. Bioorg. Med. Chem. Lett. 2012, 22, 610–612. [Google Scholar] [CrossRef]
- Muhammad, A.; Tel-Çayan, G.; Öztürk, M.; Duru, M.E.; Nadeem, S.; Anis, I.; Ng, S.W.; Shah, M. Phytochemicals from Dodonaea viscosa and their antioxidant and anticholinesterase activities with structure–activity relationships. Pharm. Biol. 2016, 54, 1–7. [Google Scholar] [CrossRef]
- Cottiglia, F.; Bonsignore, L.; Casu, L.; Deidda, D.; Pompei, R.; Casu, M.; Floris, C. Phenolic constituents from Ephedra Nebrodensis. Nat. Prod. Res. 2005, 19, 117–123. [Google Scholar] [CrossRef]
- Wahyuni, Y.; Ballester, A.R.; Sudarmonowati, E.; Bino, R.J.; Bovy, A.G. Metabolite biodiversity in pepper (Capsicum) fruits of thirty-two diverse accessions: Variation in health-related compounds and implications for breeding. Phytochemistry 2011, 72, 1358–1370. [Google Scholar] [CrossRef]
- Xavier, G.; Chandran, M.; George, T.; Beevi, S.N.; Mathew, T.B.; Paul, A.; Arimboor, R.; Vijayasree, V.; Pradeepkumar, G.T.; Rajith, R. Persistence and effect of processing on reduction of fipronil and its metabolites in chilli pepper (Capsicum annum L.) fruits. Environ. Monit. Assess. 2014, 186, 5429–5437. [Google Scholar] [CrossRef]
- Devari, S.; Jaglan, S.; Kumar, M.; Deshidi, R.; Guru, S.; Bhushan, S.; Kushwaha, M.; Gupta, A.P.; Gandhi, S.G.; Sharma, J.P.; et al. Capsaicin production by Alternaria alternata, an endophytic fungus from Capsicum annum; LC–ESI–MS/MS analysis. Phytochemistry 2014, 98, 183–189. [Google Scholar] [CrossRef]
- Yogendrarajah, P.; Van Poucke, C.; De Meulenaer, B.; De Saeger, S. Development and validation of a QuEChERS based liquid chromatography tandem mass spectrometry method for the determination of multiple mycotoxins in spices. J. Chromatogr. A 2013, 1297, 1–11. [Google Scholar] [CrossRef]
- Kawaguchi, Y.; Ochi, T.; Takaishi, Y.; Kawazoe, K.; Lee, K.-H. New Sesquiterpenes fromCapsicum annuum. J. Nat. Prod. 2004, 67, 1893–1896. [Google Scholar] [CrossRef] [PubMed]
- Marín, A.; Ferreres, F.; Tomás-Barberán, F.A.; Gil, M.I. Characterization and Quantitation of Antioxidant Constituents of Sweet Pepper (Capsicum annuumL.). J. Agric. Food Chem. 2004, 52, 3861–3869. [Google Scholar] [CrossRef] [PubMed]
- Sricharoen, P.; Lamaiphan, N.; Patthawaro, P.; Limchoowong, N.; Techawongstien, S.; Chanthai, S. Phytochemicals in Capsicum oleoresin from different varieties of hot chilli peppers with their antidiabetic and antioxidant activities due to some phenolic compounds. Ultrason. Sonochem. 2017, 38, 629–639. [Google Scholar] [CrossRef]
- Marincas, O.; Feher, I.; Magdas, D.A.; Puşcaş, R. Optimized and validated method for simultaneous extraction, identification and quantification of flavonoids and capsaicin, along with isotopic composition, in hot peppers from different regions. Food Chem. 2018, 267, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Cherchar, H.; Faraone, I.; D’Ambola, M.; Sinisgalli, C.; Piaz, F.D.; Oliva, P.; Kabouche, A.; Kabouche, Z.; Milella, L.; Vassallo, A. Phytochemistry and Antioxidant Activity of Aerial Parts of Phagnalon sordidum L. Planta Medica 2019, 85, 1008–1015. [Google Scholar] [CrossRef] [PubMed]
- Armentano, M.F.; Caterino, M.; Miglionico, R.; Ostuni, A.; Pace, M.C.; Cozzolino, F.; Monti, M.; Milella, L.; Carmosino, M.; Pucci, P.; et al. New insights on the functional role of URG7 in the cellular response to ER stress. Biol. Cell 2018, 110, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Miglionico, R.; Ostuni, A.; Armentano, M.F.; Milella, L.; Crescenzi, E.; Carmosino, M.; Bisaccia, F. ABCC6 Knockdown in HepG2 Cells Induces a Senescent-like Cell Phenotype. Cell. Mol. Biol. Lett. 2017, 22, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Lekala, C.S.; Madani, K.S.H.; Phan, A.D.T.; Maboko, M.M.; Fotouo, H.; Soundy, P.; Sultanbawa, Y.; Sivakumar, D.; Fotouo, H. Cultivar-specific responses in red sweet peppers grown under shade nets and controlled-temperature plastic tunnel environment on antioxidant constituents at harvest. Food Chem. 2019, 275, 85–94. [Google Scholar] [CrossRef]
- Hayes, J.; Allen, P.; Brunton, N.; O’Grady, M.; Kerry, J.P. Phenolic composition and in vitro antioxidant capacity of four commercial phytochemical products: Olive leaf extract (Olea europaea L.), lutein, sesamol and ellagic acid. Food Chem. 2011, 126, 948–955. [Google Scholar] [CrossRef]
- Wang, X.; Hai, C.; Liang, X.; Yu, S.; Zhang, W.; Li, Y. The protective effects of Acanthopanax senticosus Harms aqueous extracts against oxidative stress: Role of Nrf2 and antioxidant enzymes. J. Ethnopharmacol. 2010, 127, 424–432. [Google Scholar] [CrossRef]
- Wu, S.; Yue, Y.; Tian, H.; Li, Z.; Li, X.; He, W.; Ding, H. Carthamus red from Carthamus tinctorius L. exerts antioxidant and hepatoprotective effect against CCl4-induced liver damage in rats via the Nrf2 pathway. J. Ethnopharmacol. 2013, 148, 570–578. [Google Scholar] [CrossRef] [PubMed]
- Salvia, A.M.; Cuviello, F.; Coluzzi, S.; Nuccorini, R.; Attolico, I.; Pascale, S.P.; Bisaccia, F.; Pizzuti, M.; Ostuni, A. Expression of Some ATP-Binding Cassette Transporters in Acute Myeloid Leukemia. Hematol. Rep. 2017, 9, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Bonomo, M.G.; Di Tomaso, K.; Calabrone, L.; Salzano, G. Ethanol stress in Oenococcus oeni: Transcriptional response and complex physiological mechanisms. J. Appl. Microbiol. 2018, 125, 2–15. [Google Scholar] [CrossRef]
- Brechbuhl, H.M.; Gould, N.; Kachadourian, R.; Riekhof, W.R.; Voelker, D.R.; Day, B.J. Glutathione Transport Is a Unique Function of the ATP-binding Cassette Protein ABCG2*. J. Biol. Chem. 2010, 285, 16582–16587. [Google Scholar] [CrossRef] [PubMed]
- Higashikuni, Y.; Sainz, J.; Nakamura, K.; Takaoka, M.; Enomoto, S.; Iwata, H.; Tanaka, K.; Sahara, M.; Hirata, Y.; Nagai, R.; et al. The ATP-Binding Cassette Transporter ABCG2 Protects Against Pressure Overload-Induced Cardiac Hypertrophy and Heart Failure by Promoting Angiogenesis and Antioxidant Response. Arter. Thromb. Vasc. Biol. 2011, 32, 654–661. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Callaghan, D.; Juzwik, C.; Xiong, H.; Huang, P.; Zhang, W. ABCG2 reduces ROS-mediated toxicity and inflammation: A potential role in Alzheimer’s disease. J. Neurochem. 2010, 114, 1590–1604. [Google Scholar] [CrossRef]
- Cooray, H.C.; Janvilisri, T.; Van Veen, H.W.; Hladky, S.B.; A Barrand, M. Interaction of the breast cancer resistance protein with plant polyphenols. Biochem. Biophys. Res. Commun. 2004, 317, 269–275. [Google Scholar] [CrossRef]
- Caddeo, C.; Nacher, A.; Vassallo, A.; Armentano, M.F.; Pons, R.; Fernàndez-Busquets, X.; Carbone, C.; Valenti, N.; Fadda, A.M.; Manconi, M. Effect of quercetin and resveratrol co-incorporated in liposomes against inflammatory/oxidative response associated with skin cancer. Int. J. Pharm. 2016, 513, 153–163. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Rt (min) | Molecular Formula | MW | [M − H]− | [M − H]+ | MS/MS |
|---|---|---|---|---|---|---|
| Caffeic acid | 1.19 | C9H8O4 | 180.15 | 179 | 135,174 | |
| Luteolin (apiosyl acetyl) glucoside | 1.33 | C20H30O16 | 621.14 | 620 | 327 | |
| Apigenin-6,8-di-C-glucoside | 1.99 | C27H30O15 | 594.15 | 593 | 473 | |
| Vitexin | 2.34 | C21H20O10 | 432.38 | 431 | 283,311 | |
| Isoquercetin | 4.95 | C21H20O12 | 464.09 | 463 | 301 | |
| Rutin | 5.64 | C27H30O16 | 610.52 | 609 | 225,387 | |
| Kaempferol-3-O-glucoside | 6.01 | C21H20O11 | 448.38 | 447 | 285 | |
| Catechin | 33.08 | C15H14O6 | 290.21 | 289 | 203 | |
| 2,4-Di-tert-butylphenol | 33.10 | C14H22O | 206.32 | 205 | 189 | |
| Capsiate | 33.11 | C18H26O4 | 306.40 | 305 | 151,289 | |
| Ascorbic acid | 33.14 | C6H8O6 | 176.12 | 175 | 112 | |
| Dihydrocapsiate | 1.10 | C18H28O5 | 308.40 | 309 | 278,295 | |
| Luteolin | 1.29 | C15H10O6 | 286.24 | 287 | 153,171.2 | |
| Kaempherol | 1.39 | C15H10O6 | 286.23 | 287 | 241 | |
| Nordihydrocapsaicin | 1.72 | C17H27NO3 | 293.41 | 294 | 152 | |
| Tocopherol | 7.80 | C29H50O2 | 430.71 | 431 | 416 | |
| Myricetin | 12.53 | C15H10O8 | 318.23 | 319 | 227,207 | |
| Capsaicin | 15.56 | C18H27NO3 | 305.41 | 306 | 137,227 | |
| Dihydrocapsaicin | 17.43 | C18H29NO3 | 307.43 | 308 | 122,207 | |
| β-carotene | 18.22 | C40H56 | 536.87 | 537 | 277,353 | |
| Canusesnol F | 19.47 | C15H22O4 | 266.33 | 267 | 207,247 | |
| Capsorubin | 26.62 | C40H56O4 | 600.88 | 601 | 411,582 | |
| Antheraxanthin | 27.77 | C40H56O3 | 584.88 | 585 | 145 | |
| β-cryptoxanthin | 32.98 | C40H56O | 552.88 | 553 | 461 |
| Mean Diameter (nm) | P.I. # | Zeta Potential (mV) | |
|---|---|---|---|
| Empty liposomes | 81.6 ± 6.8 | 0.28 | −16.7 ± 3.7 |
| C. annuum liposomes | 83.8 ± 4.7 | 0.26 | −17.5 ± 4.2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sinisgalli, C.; Faraone, I.; Vassallo, A.; Caddeo, C.; Bisaccia, F.; Armentano, M.F.; Milella, L.; Ostuni, A. Phytochemical Profile of Capsicum annuum L. cv Senise, Incorporation into Liposomes, and Evaluation of Cellular Antioxidant Activity. Antioxidants 2020, 9, 428. https://doi.org/10.3390/antiox9050428
Sinisgalli C, Faraone I, Vassallo A, Caddeo C, Bisaccia F, Armentano MF, Milella L, Ostuni A. Phytochemical Profile of Capsicum annuum L. cv Senise, Incorporation into Liposomes, and Evaluation of Cellular Antioxidant Activity. Antioxidants. 2020; 9(5):428. https://doi.org/10.3390/antiox9050428
Chicago/Turabian StyleSinisgalli, Chiara, Immacolata Faraone, Antonio Vassallo, Carla Caddeo, Faustino Bisaccia, Maria Francesca Armentano, Luigi Milella, and Angela Ostuni. 2020. "Phytochemical Profile of Capsicum annuum L. cv Senise, Incorporation into Liposomes, and Evaluation of Cellular Antioxidant Activity" Antioxidants 9, no. 5: 428. https://doi.org/10.3390/antiox9050428
APA StyleSinisgalli, C., Faraone, I., Vassallo, A., Caddeo, C., Bisaccia, F., Armentano, M. F., Milella, L., & Ostuni, A. (2020). Phytochemical Profile of Capsicum annuum L. cv Senise, Incorporation into Liposomes, and Evaluation of Cellular Antioxidant Activity. Antioxidants, 9(5), 428. https://doi.org/10.3390/antiox9050428