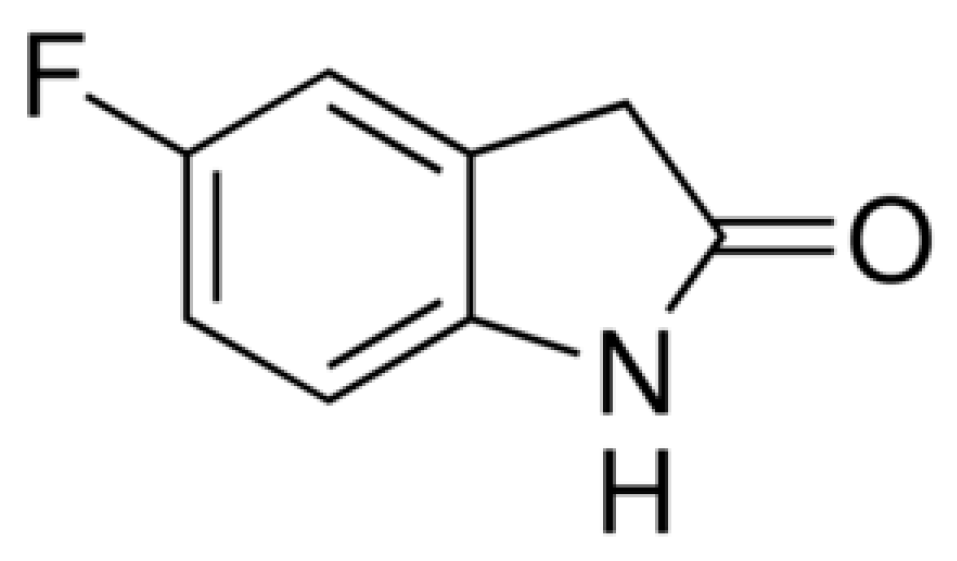
The Antinociceptive, Antioxidant and Anti-Inflammatory Effects of 5-Fluoro-2-Oxindole during Inflammatory Pain



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Induction of Inflammatory Pain
2.3. Mechanical Allodynia
2.4. Thermal Hyperalgesia
2.5. Western Blot Analysis
2.6. Experimental Procedures
2.7. Drugs
2.8. Statistical Analyses
3. Results
3.1. The Antinociceptive Effects of 5-Fluoro-2-Oxindole during Peripheral Inflammation
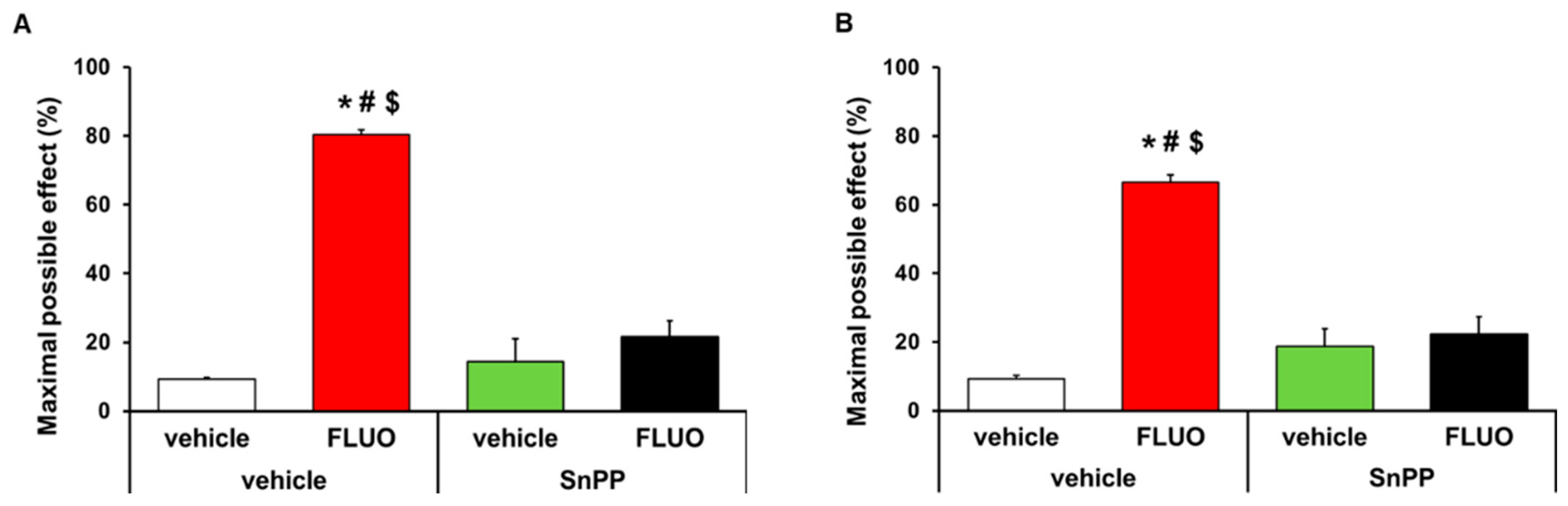
3.2. Reversion of the Antinocicetptive Effects of 5-Fluoro-2-Oxindole with SnPP
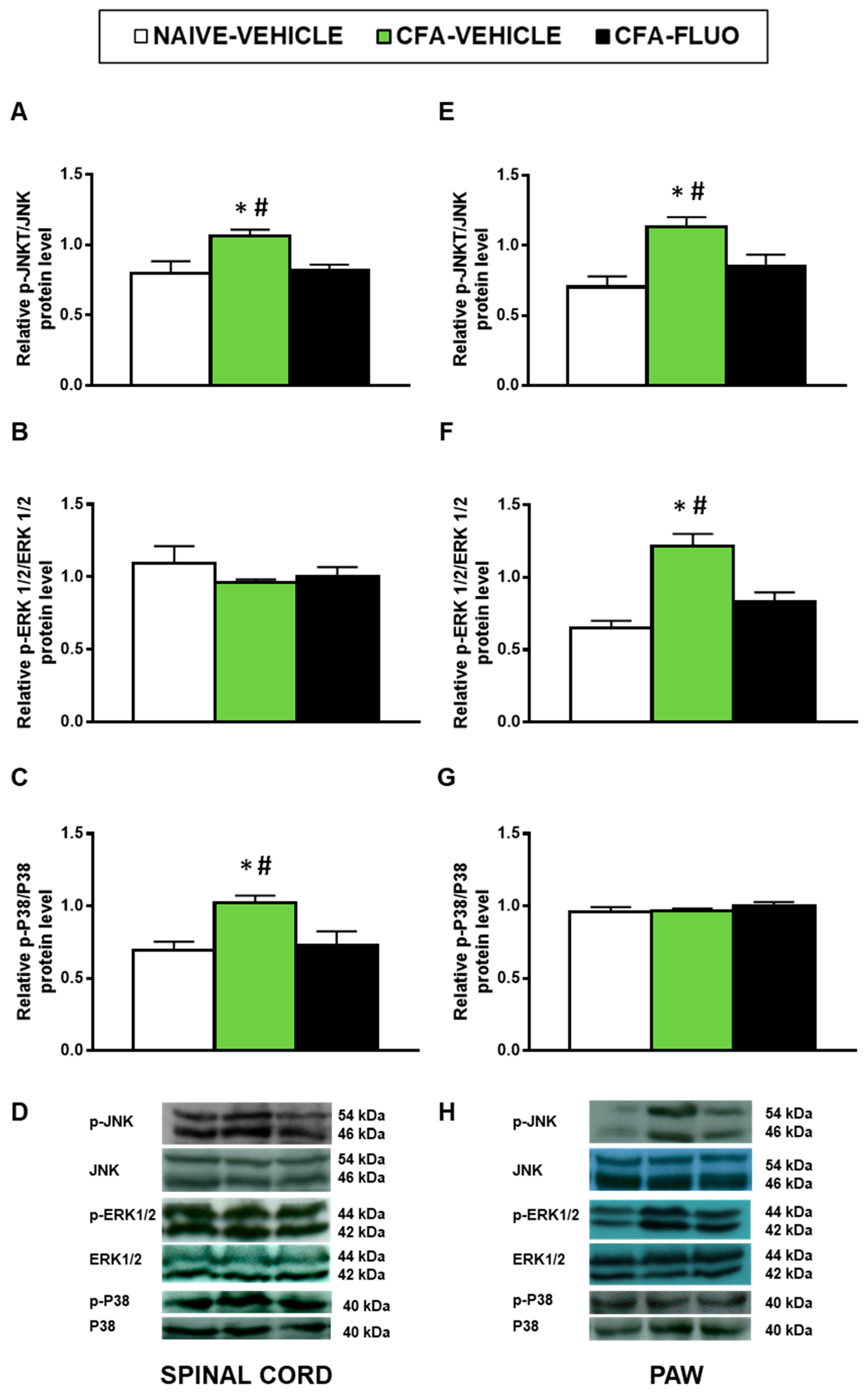
3.3. Effect of 5-Fluoro-2-Oxindole in the Expression of MAPK in the Spinal Cords and Paws of Mice Inflammatory Pain
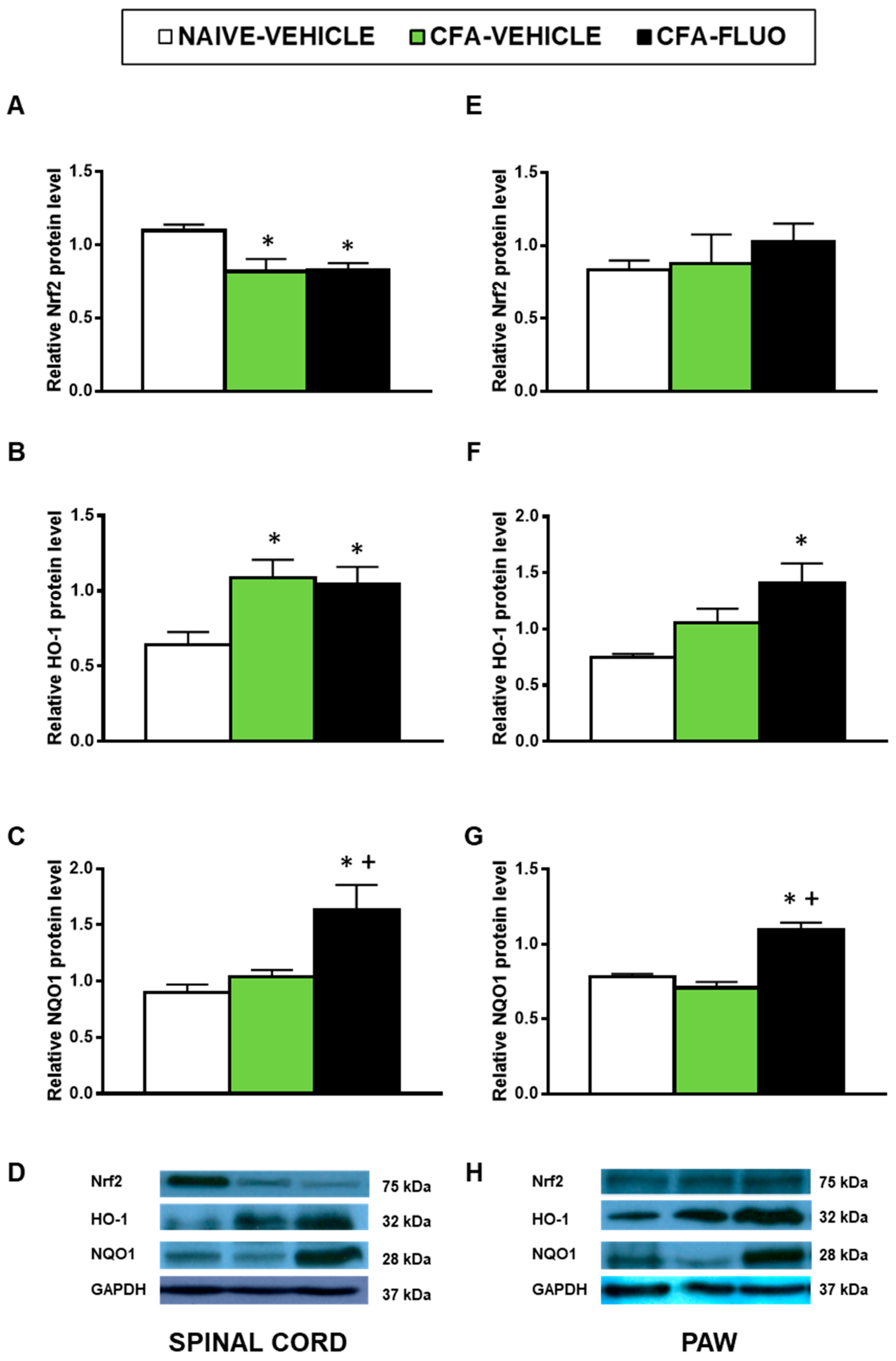
3.4. Effects of 5-Fluoro-2-Oxindole in the Expression of Antioxidant Proteins in the Spinal Cords and Paws of Animals with Peripheral Inflammation
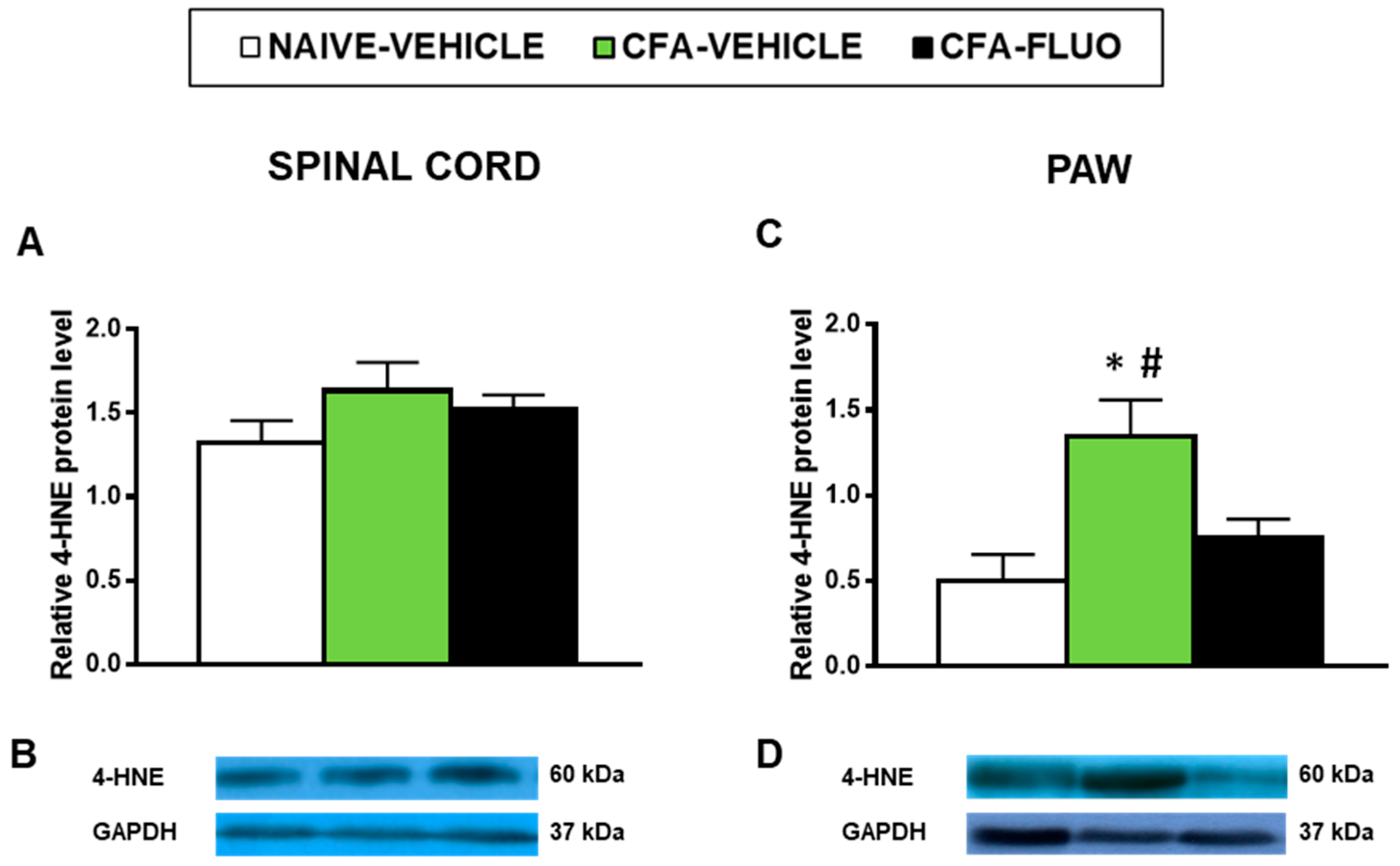
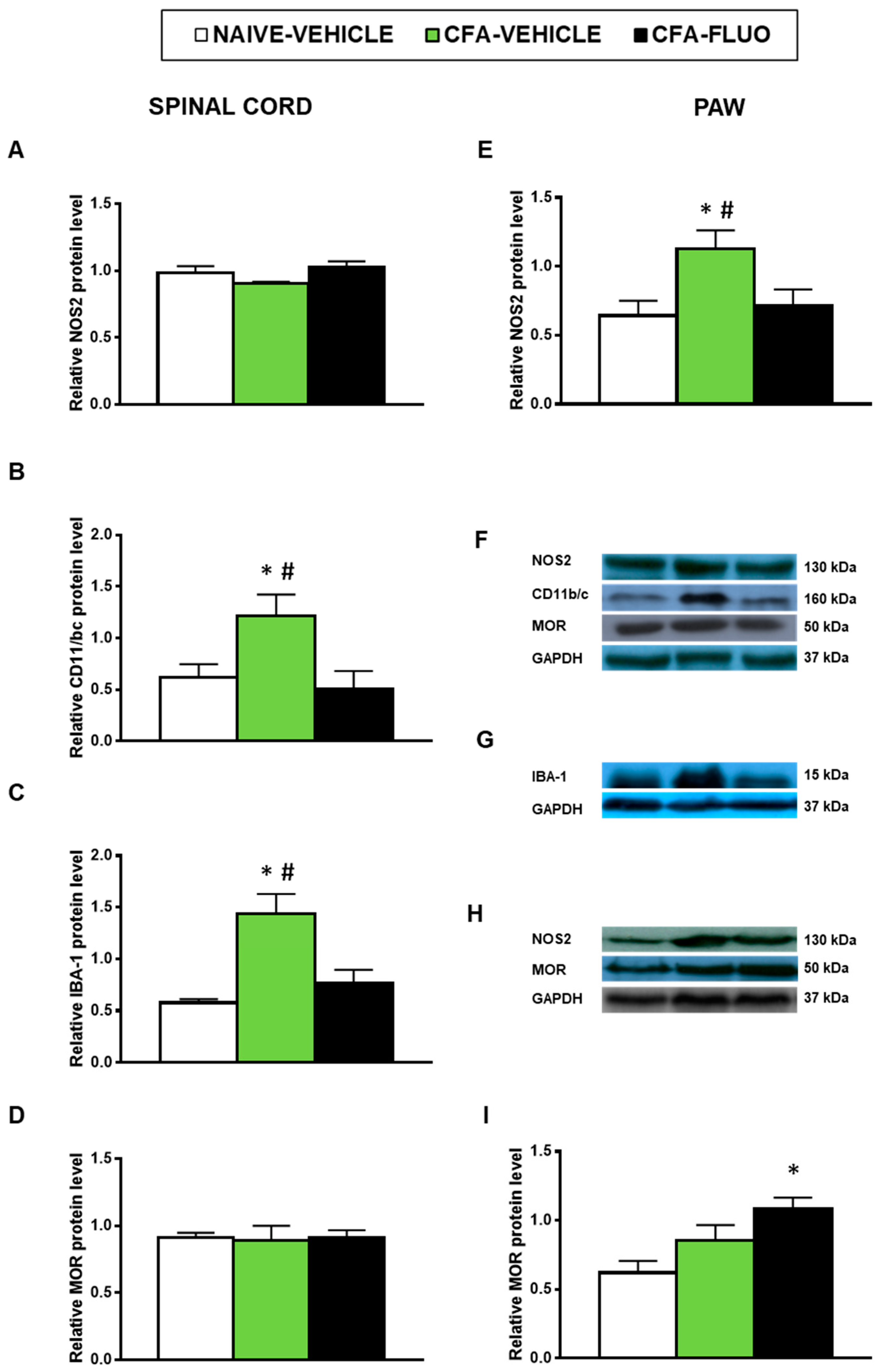
3.5. Effect of 5-Fluoro-2-Oxindole in the Protien Levels of 4-HNE, NOS2, CD11b/c, IBA-1 and MOR in the Spinal Cords and/or Paws of Animals with Peripheral Inflammation
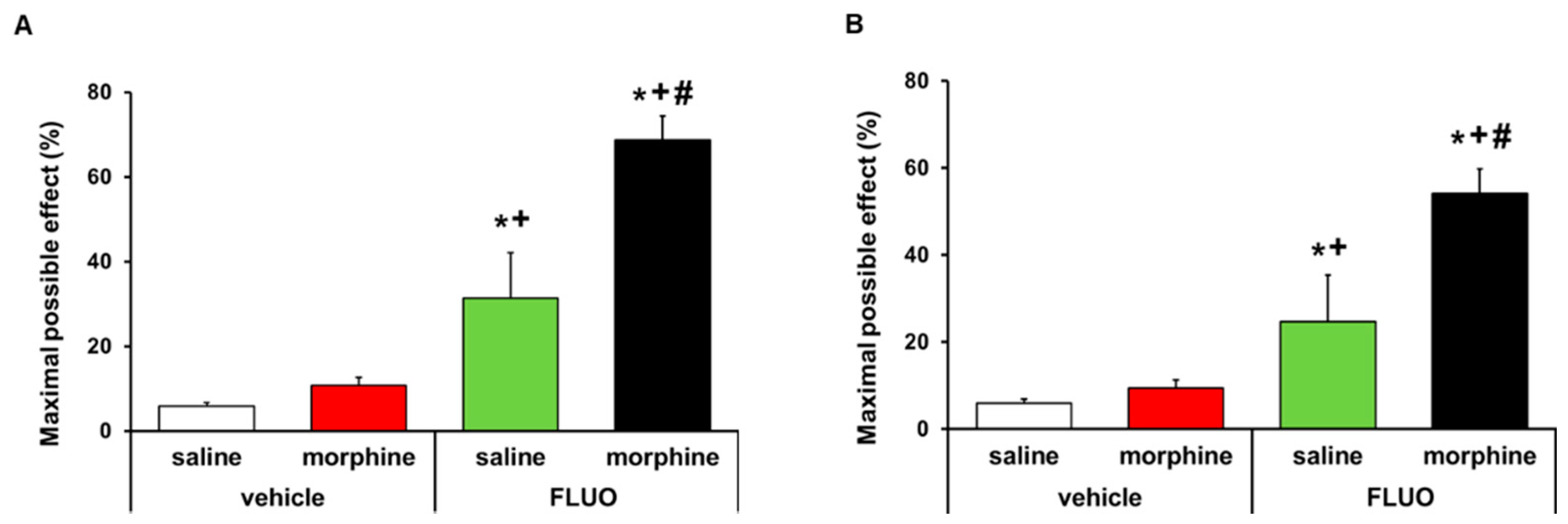
3.6. Treatment with 5-Fluoro-2-Oxindole Potencites the Local Antinociceptive Effects of Morphine
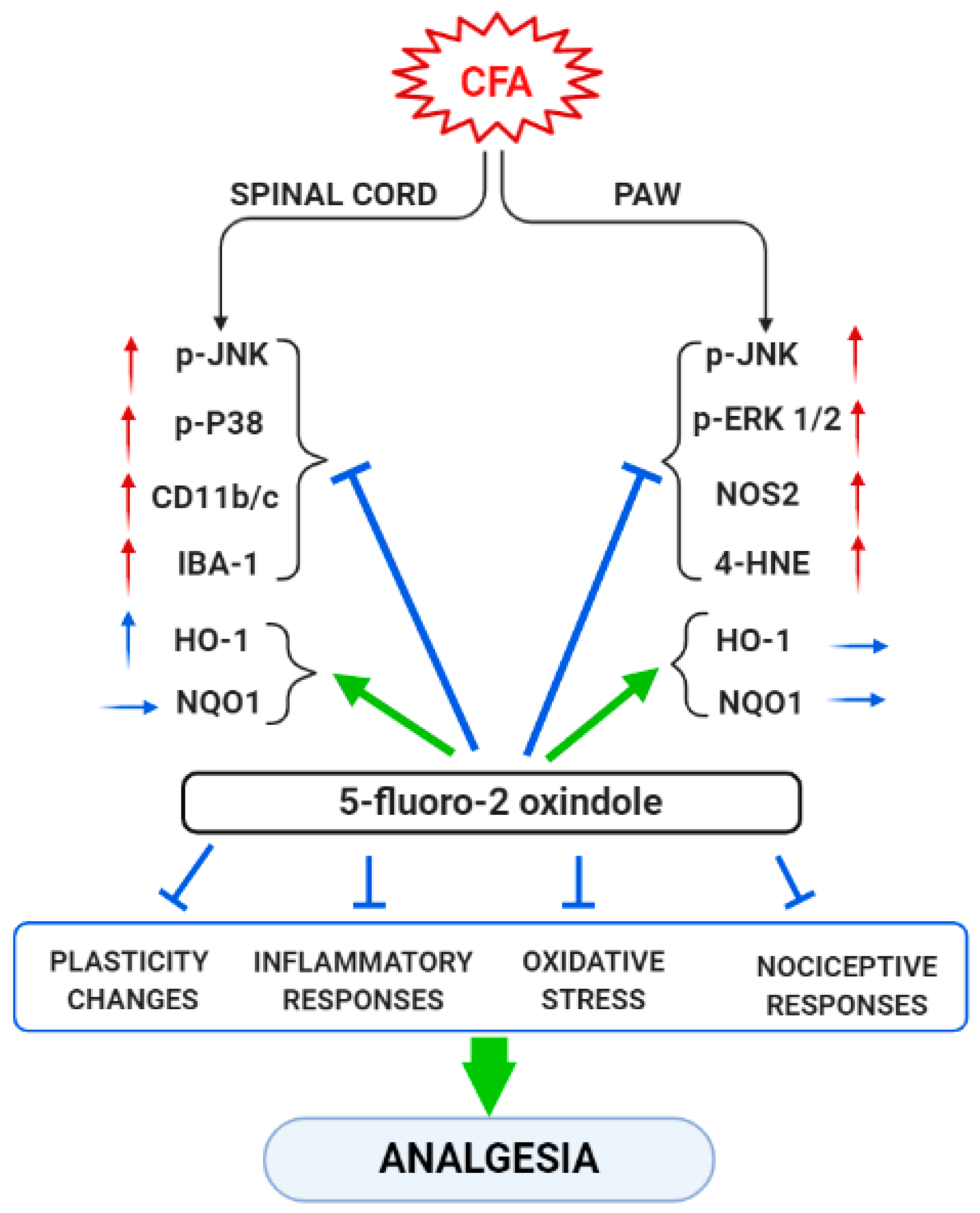
4. Discussion
5. Conclusions
Author Contributions
Funding

Conflicts of Interest
References
- Chen, C.X.; Jin, R.M.; Li, Y.K.; Zhong, J.; Yue, L.; Chen, S.C.; Zhou, J.Y. Inhibitory effect of rhynchophylline on platelet aggregation and thrombosis. Zhongguo yao li xue bao. Acta Pharmacol. Sin. 1992, 13, 126–130. [Google Scholar]
- Li, H.Q.; Ip, S.P.; Yuan, Q.J.; Zheng, G.Q.; Tsim, K.; Dong, T.; Lin, G.; Han, Y.; Liu, Y.; Xian, Y.F.; et al. Isorhynchophylline ameliorates cognitive impairment via modulating amyloid pathology, tau hyperphosphorylation and neuroinflammation: Studies in a transgenic mouse model of Alzheimer’s disease. Brain Behav. Immun. 2019, 82, 264–278. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Su, Y.; Fa, X.E. Isorhynchophylline exerts anti-inflammatory and anti-oxidative activities in LPS-stimulated murine alveolar macrophages. Life Sci. 2019, 223, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Wang, W.; Wu, X. Isorhynchophylline exerts anti-asthma effects in mice by inhibiting the proliferation of airway smooth muscle cells: The involvement of miR-200a-mediated FOXC1/NF-κB pathway. Biochem. Biophys. Res. Commun. 2020, 521, 1055–1060. [Google Scholar] [CrossRef]
- Figueiredo, G.S.; Zardo, R.S.; Silva, B.V.; Violante, F.A.; Pinto, A.C.; Fernandes, P.D. Convolutamydine A and synthetic analogues have antinociceptive properties in mice. Pharmacol. Biochem. Behav. 2013, 103, 431–439. [Google Scholar] [CrossRef]
- Fernandes, P.D.; Zardo, R.S.; Figueiredo, G.S.; Silva, B.V.; Pinto, A.C. Anti-inflammatory properties of convolutamydine A and two structural analogues. Life Sci. 2014, 116, 16–24. [Google Scholar] [CrossRef]
- Giorno, T.; Silva, B.; Pinto, A.; Fernandes, P.D. Antinociceptive effect and mechanism of action of isatin, N-methyl isatin and oxopropyl isatin in mice. Life Sci. 2016, 151, 189–198. [Google Scholar] [CrossRef]
- Ferreira-Chamorro, P.; Redondo, A.; Riego, G.; Pol, O. Treatment with 5-fluoro-2-oxindole Increases the Antinociceptive Effects of Morphine and Inhibits Neuropathic Pain. Cell. Mol. Neurobiol. 2020. [Google Scholar] [CrossRef]
- Wolf, S.A.; Boddeke, H.W.; Kettenmann, H. Microglia in physiology and disease. Annu. Rev. Physiol. 2017, 79, 619–643. [Google Scholar] [CrossRef]
- Donnelly, C.R.; Andriessen, A.S.; Chen, G.; Wang, K.; Jiang, C.; Maixner, W.; Ji, R.R. Central Nervous System Targets: Glial Cell Mechanisms in Chronic Pain. Neurotherapeutics 2020, 17, 846–860. [Google Scholar] [CrossRef]
- Redondo, A.; Chamorro, P.; Riego, G.; Leánez, S.; Pol, O. Treatment with Sulforaphane Produces Antinociception and Improves Morphine Effects during Inflammatory Pain in Mice. J. Pharmacol. Exp. Ther. 2017, 363, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Yuan, D.; Ma, B.; Yang, J.Y.; Xie, Y.Y.; Wang, L.; Zhang, L.J.; Kano, Y.; Wu, C.F. Anti-inflammatory effects of rhynchophylline and isorhynchophylline in mouse N9 microglial cells and the molecular mechanism. Int. Immunopharmacol. 2009, 9, 1549–1554. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Qu, R.; Zhu, S.; Zhang, R.; Ma, S. Rhynchophylline attenuates LPS-induced pro-inflammatory responses through down-regulation of MAPK/NF-κB signaling pathways in primary microglia. Phytother. Res. 2012, 26, 1528–1533. [Google Scholar] [CrossRef] [PubMed]
- Jurga, A.M.; Piotrowska, A.; Starnowska, J.; Rojewska, E.; Makuch, W.; Mika, J. Treatment with a carbon monoxide-releasing molecule (CORM-2) inhibits neuropathic pain and enhances opioid effectiveness in rats. Pharmacol. Rep. 2016, 68, 206–213. [Google Scholar] [CrossRef]
- Ferreira-Chamorro, P.; Redondo, A.; Riego, G.; Leánez, S.; Pol, O. Sulforaphane inhibited the nociceptive responses, anxiety- and depressive-like behaviors associated with neuropathic pain and improved the anti-allodynic effects of morphine in mice. Front. Pharmacol. 2018, 9, 1332. [Google Scholar] [CrossRef] [PubMed]
- Riego, G.; Redondo, A.; Leánez, S.; Pol, O. Mechanism implicated in the anti-allodynic and anti-hyperalgesic effects induced by the activation of heme oxygenase 1/carbon monoxide signaling pathway in the central nervous system of mice with neuropathic pain. Biochem. Pharmacol. 2018, 148, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Díaz, A.F.; Polo, S.; Gallardo, N.; Leánez, S.; Pol, O. Analgesic and antidepressant effects of oltipraz on neuropathic pain in mice by modulating microglial activation. J. Clin. Med. 2019, 8, 890. [Google Scholar] [CrossRef]
- Hervera, A.; Leánez, S.; Motterlini, R.; Pol, O. Treatment with carbon monoxide-releasing molecules and an HO-1 inducer enhances the effects and expression of µ-opioid receptors during neuropathic pain. Anesthesiology 2013, 118, 1180–1197. [Google Scholar] [CrossRef]
- Carcolé, M.; Castany, S.; Leánez, S.; Pol, O. Treatment with a heme oxygenase 1 inducer enhances the antinociceptive effects of µ-opioid, δ-opioid, and cannabinoid 2 receptors during inflammatory pain. J. Pharmcol. Exp. Ther. 2014, 351, 224–232. [Google Scholar] [CrossRef]
- Leánez, S.; Hervera, A.; Pol, O. Peripheral antinociceptive effects of mu- and delta-opioid receptor agonists in NOS2 and NOS1 knockout mice during chronic inflammatory pain. Eur. J. Pharmacol. 2009, 602, 41–49. [Google Scholar] [CrossRef]
- Chaplan, S.R.; Bach, F.W.; Pogrel, J.W.; Chung, J.M.; Yaksh, T.L. Quantitative assessment of tactile allodynia in the rat paw. J. Neurosci. Methods 1994, 53, 55–63. [Google Scholar] [CrossRef]
- Hargreaves, K.; Dubner, R.; Brown, F.; Flores, C.; Joris, J. A new and sensitive method for measuring thermal nociception in cutaneous hyperalgesia. Pain 1988, 32, 77–88. [Google Scholar] [CrossRef]
- Godai, K.; Kanmura, Y. Heme oxygenase-1 inducer and carbon monoxide-releasing molecule enhance the effects of gabapentinoids by modulating glial activation during neuropathic pain in mice. Pain Rep. 2018, 3, e677. [Google Scholar] [CrossRef] [PubMed]
- Middlemas, A.B.; Agthong, S.; Tomlinson, D.R. Phosphorylation of c-Jun N-terminal kinase (JNK) in sensory neurones of diabetic rats, with possible effects on nerve conduction and neuropathic pain: Prevention with an aldose reductase inhibitor. Diabetologia 2006, 49, 580–587. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.J.; Ji, R.R. Light touch induces ERK activation in superficial dorsal horn neurons after inflammation: Involvement of spinal astrocytes and JNK signaling in touch-evoked central sensitization and mechanical allodynia. J. Neurochem. 2010, 115, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.J.; Xu, Z.Z.; Liu, Y.C.; Wen, Y.R.; Decosterd, I.; Ji, R.R. The c-Jun N-terminal kinase 1 (JNK1) in spinal astrocytes is required for the maintenance of bilateral mechanical allodynia under a persistent inflammatory pain condition. Pain 2010, 148, 309–319. [Google Scholar] [CrossRef]
- Borges, G.; Berrocoso, E.; Mico, J.A.; Neto, F. ERK1/2: Function, signaling and implication in pain and pain-related anxio-depressive disorders. Prog. Neuropsychopharmacol. Biol. Psychiatry 2015, 60, 77–92. [Google Scholar] [CrossRef]
- Yin, X.; Jing, Y.; Chen, Q.; Abbas, A.B.; Hu, J.; Xu, H. The intraperitoneal administration of MOTS-c produces antinociceptive and anti-inflammatory effects through the activation of AMPK pathway in the mouse formalin test. Eur. J. Pharmacol. 2020. [Google Scholar] [CrossRef]
- Hu, F.; Zhang, H.H.; Yang, B.X.; Huang, J.L.; Shun, J.L.; Kong, F.J.; Xu, P.; Chen, Z.G.; Lu, J.M. Cdk5 contributes to inflammation-induced thermal hyperalgesia mediated by the p38 MAPK pathway in microglia. Brain Res. 2015, 1619, 166–175. [Google Scholar]
- Martini, A.C.; Berta, T.; Forner, S.; Chen, G.; Bento, A.F.; Ji, R.R.; Rae, G.A. Lipoxin A4 inhibits microglial activation and reduces neuroinflammation and neuropathic pain after spinal cord hemisection. J. Neuroinflammation 2016, 13, 75. [Google Scholar] [CrossRef]
- Rojewska, E.; Popiolek-Barczyk, K.; Jurga, A.M.; Makuch, W.; Przewlocka, B.; Mika, J. Involvement of pro- and antinociceptive factors in minocycline analgesia in rat neuropathic pain model. J. Neuroimmunol. 2014, 277, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Boettger, M.K.; Reif, A.; Schmitt, A.; Uçeyler, N.; Sommer, C. Nitric oxide synthase modulates CFA-induced thermal hyperalgesia through cytokine regulation in mice. Mol. Pain 2010, 6, 13. [Google Scholar] [CrossRef] [PubMed]
- Moreno, P.; Cazuza, R.A.; Mendes-Gomes, J.; Díaz, A.F.; Polo, S.; Leánez, S.; Leite-Panissi, C.R.A.; Pol, O. The Effects of Cobalt Protoporphyrin IX and Tricarbonyldichlororuthenium (II) Dimer Treatments and Its Interaction with Nitric Oxide in the Locus Coeruleus of Mice with Peripheral Inflammation. Int. J. Mol. Sci. 2019, 20, 2211. [Google Scholar] [CrossRef] [PubMed]
- Negi, G.; Nakkina, V.; Kamble, P.; Sharma, S.S. Heme oxygenase-1, a novel target for the treatment of diabetic complications: Focus on diabetic peripheral neuropathy. Pharmcol. Res. 2015, 102, 158–167. [Google Scholar] [CrossRef] [PubMed]
- McDonnell, C.; Leánez, S.; Pol, O. The induction of the transcription factor Nrf2 enhances the antinociceptive effects of delta-opioid receptors in diabetic mice. PLoS ONE 2017, 12, e0180998. [Google Scholar] [CrossRef]
- McDonnell, C.; Leánez, S.; Pol, O. The Inhibitory Effects of Cobalt Protoporphyrin IX and Cannabinoid 2 Receptor Agonists in Type 2 Diabetic Mice. Int. J. Mol. Sci. 2017, 18, 2268. [Google Scholar] [CrossRef]
- Negrete, R.; Hervera, A.; Leánez, S.; Pol, O. Treatment with a carbon monoxide-releasing molecule inhibits chronic inflammatory pain in mice: Nitric oxide contribution. Psychopharmacology 2014, 231, 853–861. [Google Scholar] [CrossRef]
- Nioi, P.; Hayes, J.D. Contribution of NAD(P)H:quinone oxidoreductase 1 to protection against carcinogenesis, and regulation of its gene by the Nrf2 basic-region leucine zipper and the arylhydrocarbon receptor basic helix-loop-helix transcription factors. Mutat. Res. 2004, 555, 149–171. [Google Scholar] [CrossRef]
- Luo, S.; Kang, S.S.; Wang, Z.H.; Liu, X.; Day, J.X.; Wu, Z.; Peng, J.; Xiang, D.; Springer, W.; Ye, K. Akt Phosphorylates NQO1 and Triggers its Degradation, Abolishing Its Antioxidative Activities in Parkinson’s Disease. J. Neurosci. 2019, 39, 7291–7305. [Google Scholar] [CrossRef]
- Trevisani, M.; Siemens, J.; Materazzi, S.; Bautista, D.M.; Nassini, R.; Campi, B.; Imamachi, N.; Andrè, E.; Patacchini, R.; Cottrell, G.S.; et al. 4-Hydroxynonenal, an endogenous aldehyde, causes pain and neurogenic inflammation through activation of the irritant receptor TRPA1. Proc. Natl. Acad. Sci. USA 2007, 104, 13519–13524. [Google Scholar] [CrossRef]
- Sauer, R.S.; Krummenacher, I.; Bankoglu, E.E.; Yang, S.; Oehler, B.; Schöppler, F.; Mohammadi, M.; Güntzel, P.; Ben-Kraiem, A.; Holzgrabe, U.; et al. Stabilization of Delphinidin in Complex with Sulfobutylether-β-Cyclodextrin Allows for Antinociception in Inflammatory Pain. Antioxid. Redox Signal. 2020. [Google Scholar] [CrossRef] [PubMed]
- Ilari, S.; Giancotti, L.A.; Lauro, F.; Dagostino, C.; Gliozzi, M.; Malafoglia, V.; Sansone, L.; Palma, E.; Tafani, M.; Russo, M.A.; et al. Antioxidant modulation of sirtuin 3 during acute inflammatory pain: The ROS control. Pharmacol. Res. 2020, 157, 104851. [Google Scholar] [CrossRef] [PubMed]
- Castany, S.; Carcolé, M.; Leánez, S.; Pol, O. The Induction of Heme Oxygenase 1 Decreases Painful Diabetic Neuropathy and Enhances the Antinociceptive Effects of Morphine in Diabetic Mice. PLoS ONE 2016, 11, e0146427. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Redondo, A.; Riego, G.; Pol, O. The Antinociceptive, Antioxidant and Anti-Inflammatory Effects of 5-Fluoro-2-Oxindole during Inflammatory Pain. Antioxidants 2020, 9, 1249. https://doi.org/10.3390/antiox9121249
Redondo A, Riego G, Pol O. The Antinociceptive, Antioxidant and Anti-Inflammatory Effects of 5-Fluoro-2-Oxindole during Inflammatory Pain. Antioxidants. 2020; 9(12):1249. https://doi.org/10.3390/antiox9121249
Chicago/Turabian StyleRedondo, Alejandro, Gabriela Riego, and Olga Pol. 2020. "The Antinociceptive, Antioxidant and Anti-Inflammatory Effects of 5-Fluoro-2-Oxindole during Inflammatory Pain" Antioxidants 9, no. 12: 1249. https://doi.org/10.3390/antiox9121249
APA StyleRedondo, A., Riego, G., & Pol, O. (2020). The Antinociceptive, Antioxidant and Anti-Inflammatory Effects of 5-Fluoro-2-Oxindole during Inflammatory Pain. Antioxidants, 9(12), 1249. https://doi.org/10.3390/antiox9121249