The Modulatory Role of Vitamin C in Boldenone Undecylenate Induced Testicular Oxidative Damage and Androgen Receptor Dysregulation in Adult Male Rats

 ,
,  , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals
2.3. Experimental Design
- Group I (control group) orally given physiological saline.
- Group II (vehicle control group) was intramuscularly injected with 0.25 mL/kg b.wt. sesame oil as a vehicle once a week for eight weeks.
- Group III (Vit-C-treated group) was gavaged with Vit-C dissolved in distilled water (120 mg/kg b.wt.) every day for eight weeks [32].
- Group IV (BOL-treated group) was intramuscularly injected with BOL (5 mg/kg b.wt.) once a week for eight weeks, according to a previously mentioned dose of Bueno, et al. [33].
- Group V (BOL+Vit-C-treated group) was co-treated with BOL (5 mg/kg b.wt.) and Vit-C (120 mg/kg b.wt.) at the same declared duration and route.
2.4. Sampling
2.5. Semen Evaluation
2.6. Hormonal Assay
2.7. Testicular Enzymes Evaluation
2.8. Testicular Oxidative/Antioxidant Status
2.9. Histopathological Evaluation
2.10. Immunohistochemical Investigation of Androgen Receptor Expression in the Sertoli and Leydig Cells
2.11. Statistical Analysis
3. Results
3.1. Effect of BOL and Vit-C on the Body and Reproductive Organ Weights
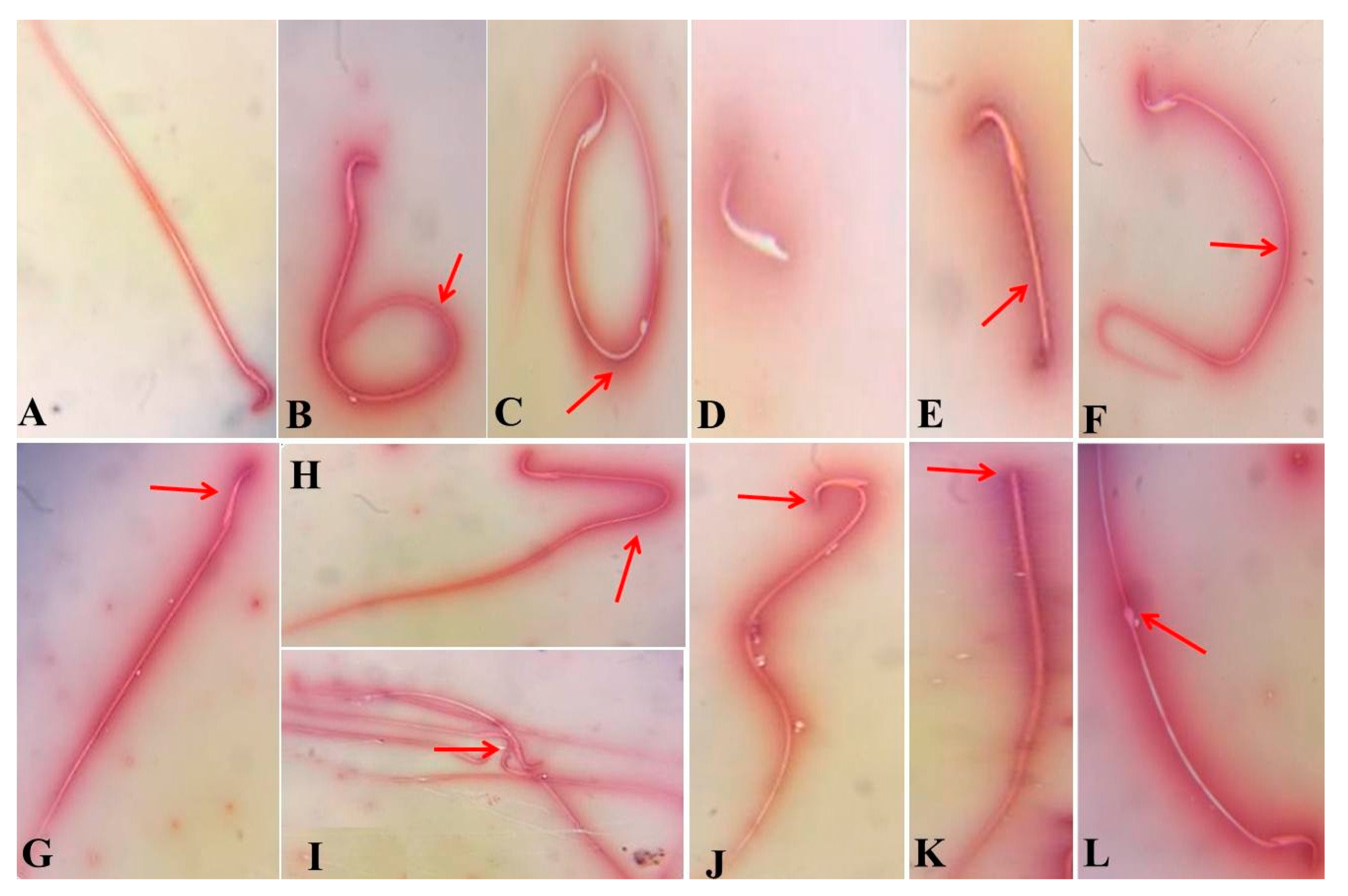
3.2. Effect of BOL and Vit-C on Spermiogram
3.3. Effect of BOL and Vit-C on Male Reproductive Hormones
3.4. Effect of BOL and Vit-C on Testicular Enzymes
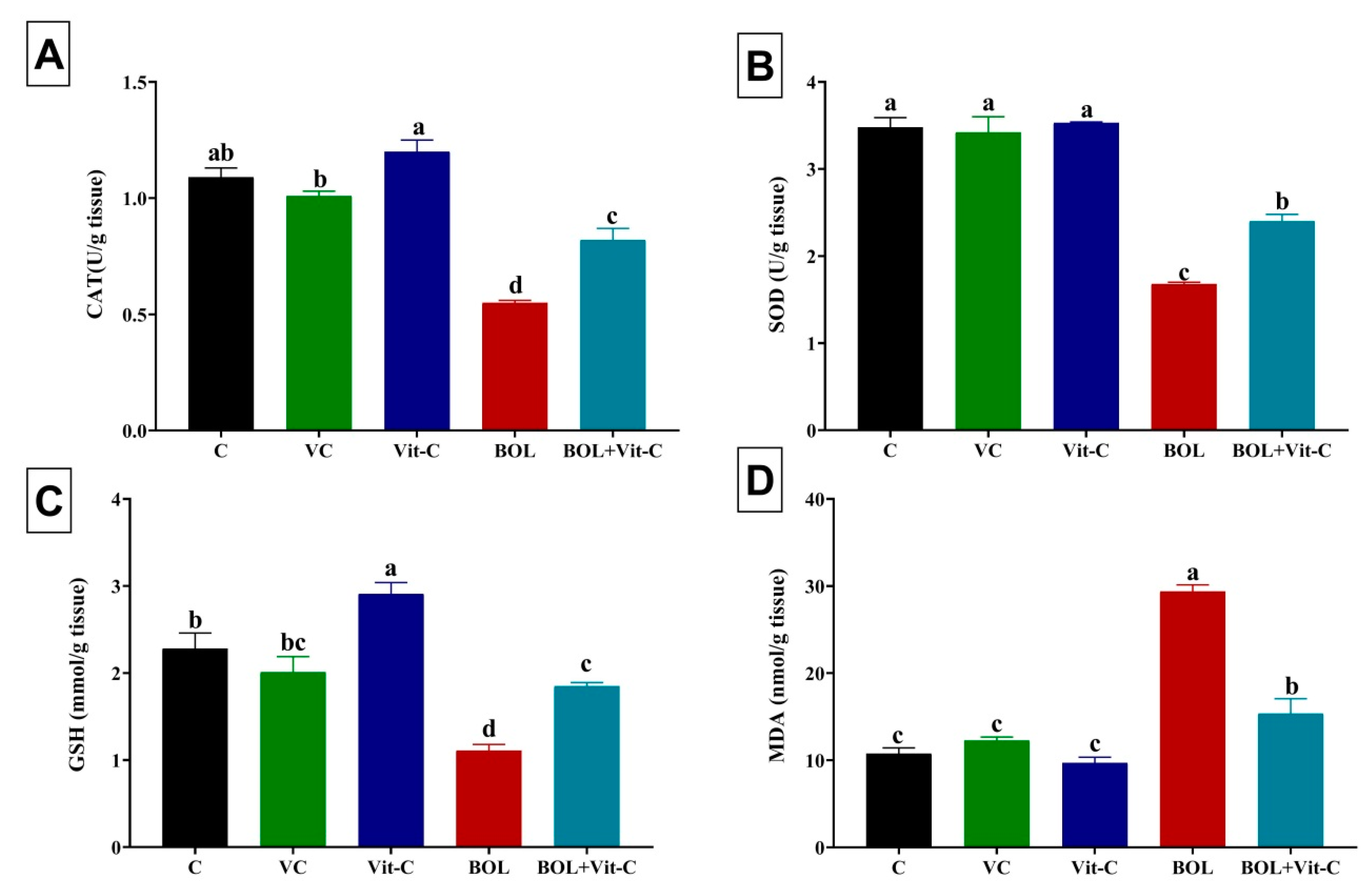
3.5. Effect of BOL and Vit-C on Testicular Antioxidant/Oxidative Status
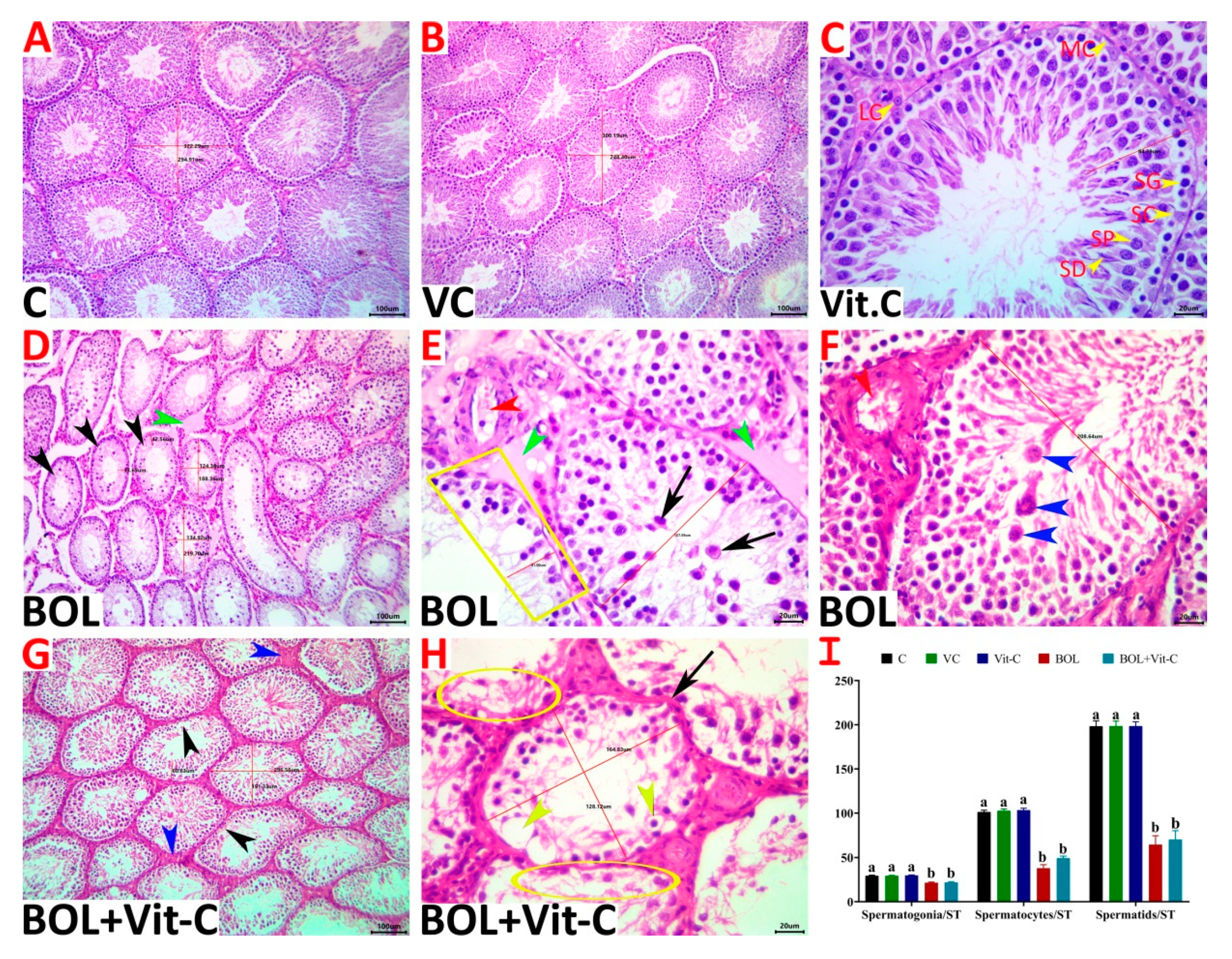
3.6. Histopathological Evaluation
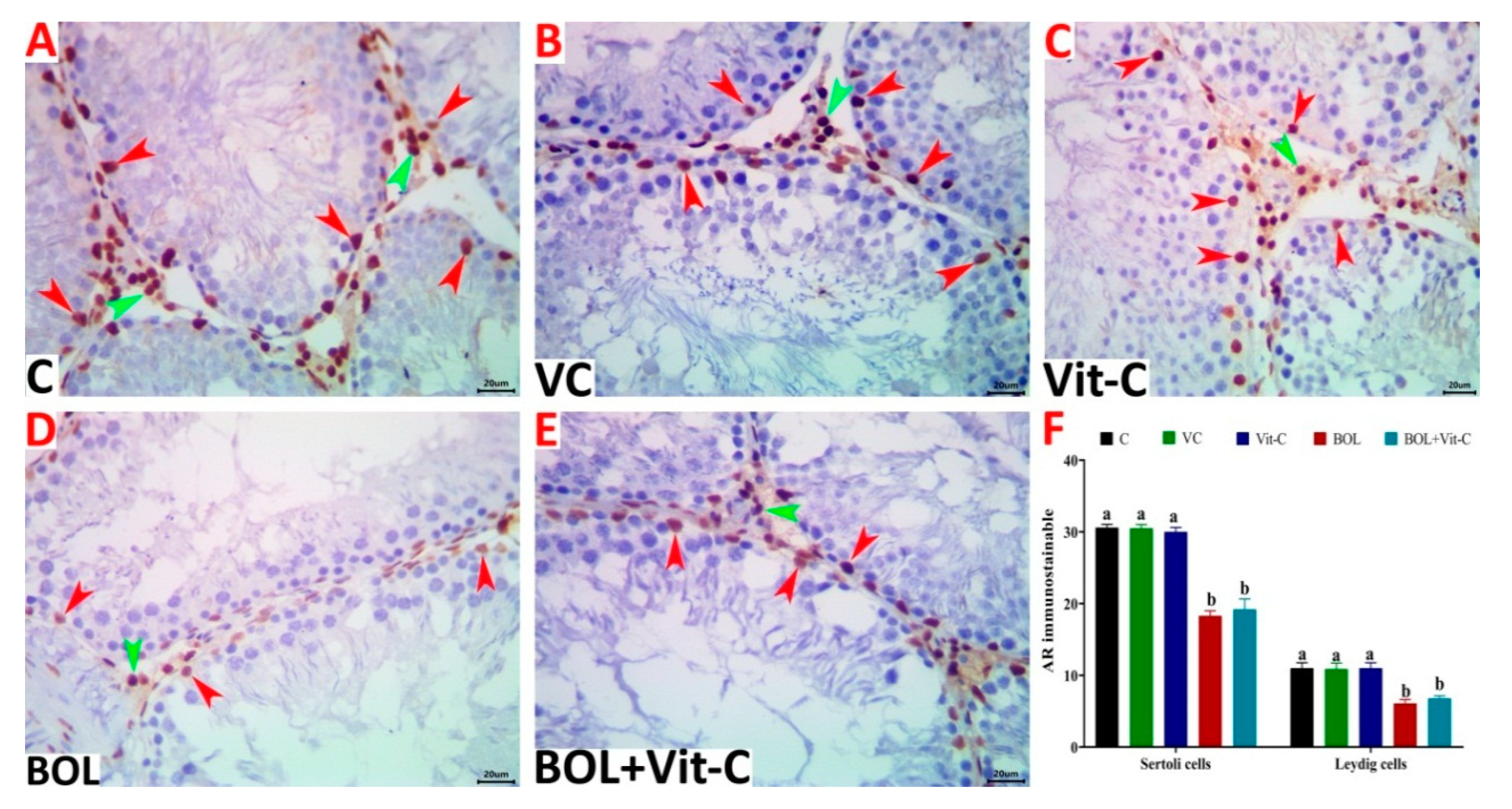
3.7. Immunohistochemical Assessment of AR
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Anaissie, J.; Pastuszak, A.W.; Khera, M. Chapter 4.1—Testosterone and Men’s Health. In Effects of Lifestyle on Men’s Health; Yafi, F.A., Yafi, N.R., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 235–251. [Google Scholar] [CrossRef]
- Kicman, A.T. Pharmacology of anabolic steroids. Br. J. Pharmacol. 2008, 154, 502–521. [Google Scholar] [CrossRef]
- El Osta, R.; Almont, T.; Diligent, C.; Hubert, N.; Eschwège, P.; Hubert, J. Anabolic steroids abuse and male infertility. Basic Clin. Androl. 2016, 26, 2. [Google Scholar]
- Jassim, A.M.; Alzamely, H.A.; Hamad, A.G. Study of the testicular damage induced by dianabol and its effect on morphological and histological changes in albino male rats. J. Agric. Vet. Sci. 2015, 8, 24–32. [Google Scholar]
- Pope, H.G., Jr.; Kanayama, G.; Athey, A.; Ryan, E.; Hudson, J.I.; Baggish, A. The lifetime prevalence of anabolic-androgenic steroid use and dependence in Americans: Current best estimates. Am. J. Addict. 2014, 23, 371–377. [Google Scholar] [CrossRef]
- Melnik, B.; Jansen, T.; Grabbe, S. Abuse of anabolic-androgenic steroids and bodybuilding acne: An underestimated health problem. J. Dtsch. Dermatol. Ges. 2007, 5, 110–117. [Google Scholar] [CrossRef]
- Friedl, K.E. Effects of Testosterone and Related Androgens on Athletic Performance in Men. In The Endocrine System in Sports and Exercise; John Wiley & Sons: Hoboken, NJ, USA, 2005; pp. 525–543. [Google Scholar] [CrossRef]
- Sarkey, S.; Azcoitia, I.; Garcia-Segura, L.M.; Garcia-Ovejero, D.; DonCarlos, L.L. Classical androgen receptors in non-classical sites in the brain. Horm. Behav. 2008, 53, 753–764. [Google Scholar] [CrossRef]
- Cohen, J.; Collins, R.; Darkes, J.; Gwartney, D. A league of their own: Demographics, motivations and patterns of use of 1,955 male adult non-medical anabolic steroid users in the United States. Int. Soc. Sports Nutr. 2007, 4, 12. [Google Scholar] [CrossRef]
- Tousson, E.; El-Moghazy, M.; Massoud, A.; Akel, A. Histopathological and immunohistochemical changes in the testes of rabbits after injection with the growth promoter boldenone. Reprod. Sci. 2012, 19, 253–259. [Google Scholar] [CrossRef]
- Oda, S.S.; El-Ashmawy, I.M. Adverse effects of the anabolic steroid, boldenone undecylenate, on reproductive functions of male rabbits. Int. J. Exp. Pathol. 2012, 93, 172–178. [Google Scholar] [CrossRef]
- Tousson, E.; El-Moghazy, M.; Massoud, A.; El-Atrash, A.; Sweef, O.; Akel, A. Physiological and biochemical changes after boldenone injection in adult rabbits. Toxicol. Ind. Health 2016, 32, 177–182. [Google Scholar] [CrossRef]
- Soma, L.; Uboh, C.; Guan, F.; McDonnell, S.; Pack, J. Pharmacokinetics of boldenone and stanozolol and the results of quantification of anabolic and androgenic steroids in race horses and nonrace horses. J. Vet. Pharmacol. Ther. 2007, 30, 101–108. [Google Scholar] [CrossRef]
- Reyes-Vallejo, L. Current use and abuse of anabolic steroids. Actas Urol. Esp. 2020, 44, 309–313. [Google Scholar] [CrossRef]
- Dohle, G.; Smit, M.; Weber, R. Androgens and male fertility. World J. Urol. 2003, 21, 341–345. [Google Scholar] [CrossRef]
- Goldman, A.L.; Pope, H.G., Jr.; Bhasin, S. The health threat posed by the hidden epidemic of anabolic steroid use and body image disorders among young men. J. Clin. Endocrinol. Metab. 2019, 104, 1069–1074. [Google Scholar] [CrossRef]
- Coward, R.M.; Rajanahally, S.; Kovac, J.R.; Smith, R.P.; Pastuszak, A.W.; Lipshultz, L.I. Anabolic steroid induced hypogonadism in young men. J. Urol. 2013, 190, 2200–2205. [Google Scholar] [CrossRef]
- Anawalt, B.D. Diagnosis and management of anabolic androgenic steroid use. J. Clin. Endocrinol. Metab. 2019, 104, 2490–2500. [Google Scholar] [CrossRef]
- Rahnema, C.D.; Lipshultz, L.I.; Crosnoe, L.E.; Kovac, J.R.; Kim, E.D. Anabolic steroid–induced hypogonadism: Diagnosis and treatment. Fertil. Steril. 2014, 101, 1271–1279. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.D.B.; Mendonça, A.A.S.; Sousa, R.C.; Silva, T.G.S.; Bigonha, S.M.; Santos, E.C.; Gonçalves, R.V.; Novaes, R.D. Food-drug interaction: Anabolic steroids aggravate hepatic lipotoxicity and nonalcoholic fatty liver disease induced by trans fatty acids. Food Chem. Toxicol. 2018, 116, 360–368. [Google Scholar] [CrossRef]
- Parente Filho, S.L.A.; Gomes, P.E.A.d.C.; Forte, G.A.; Lima, L.L.L.; Silva Júnior, G.B.d.; Meneses, G.C.; Martins, A.M.C.; Daher, E.D.F. Kidney disease associated with androgenic–anabolic steroids and vitamin supplements abuse: Be aware! Nefrología 2020, 40, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Saleh, R.A.; Bedaiwy, M.A. Role of reactive oxygen species in the pathophysiology of human reproduction. Fertil. Steril. 2003, 79, 829–843. [Google Scholar] [CrossRef]
- Sherwood, L. Human Physiology from Cell to Systems, 6th ed.; Cengange Learning: Boston, MA, USA, 2011. [Google Scholar]
- Abd El-Hakim, Y.M.; Mohamed, W.A.; El-Metwally, A.E. Spirulina platensis attenuates furan reprotoxicity by regulating oxidative stress, inflammation, and apoptosis in testis of rats. Ecotoxicol. Environ. Saf. 2018, 161, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Abd-Elhakim, Y.M.; Ghoneim, M.H.; Ebraheim, L.L.; Imam, T.S. Taurine and hesperidin rescues carbon tetrachloride-triggered testicular and kidney damage in rat via modulating oxidative stress and inflammation. Life Sci. 2020, 254, 117782. [Google Scholar] [CrossRef] [PubMed]
- Jacob, R.A.; Planalto, F.S.; Agee, R.E. Cellular ascorbate depletion in healthy men. J. Nutr. 1992, 122, 1111–1118. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, G.S.; Fernandez, C.D.; Campos, K.E.; Damasceno, D.C.; Anselmo-Franci, J.A.; Kempinas, W.D. Vitamin C partially attenuates male reproductive deficits in hyperglycemic rats. Reprod. Biol. Endocrinol. 2011, 9, 100. [Google Scholar] [CrossRef]
- Kini, R.D.; Nayanatara, A.K.; Pai, S.; Ramswamy, C.; Bhat, M.R.; Mantur, V.S. Infertility in male wistar rats induced by cadmium chloride: Role of ascorbic acid. J. Chin. Clin. Med. 2009, 4, 616–621. [Google Scholar]
- Sonmez, M.; Demirci, E. The effect of intramuscular vitamin C administration on semen quality in rams. Health Vet. Sci. 2003, 17, 195–201. [Google Scholar]
- Fazeli, P.; Zamiri, M.; Farshad, A.; Khalili, B. Short paper: Effects of vitamin C on testicular and seminal characteristics of markhoz goats. Iran. J. Vet. Res. 2010, 11, 11,267–272. [Google Scholar]
- Lewis, S.E.; Aitken, R.J.; Conner, S.J.; De Iuliis, G.; Evenson, D.P.; Henkel, R.; Giwercman, A.; Gharagozloo, P. The impact of sperm DNA damage in assisted conception and beyond: Recent advances in diagnosis and treatment. Reprod. Biomed. Online 2013, 27, 325–337. [Google Scholar] [CrossRef]
- Nisar, N.A.; Sultana, M.; Baba, N.A.; Para, P.A.; Waiz, H.A.; Bhat, S.A.; Zargar, F.A.; Ahmad, I. Ameliorative effect of vitamin C on the haematological changes induced by exposure of chlorpyriphos and lead acetate in Wistar rats. Comp. Clin. Pathol. 2014, 23, 829–834. [Google Scholar] [CrossRef]
- Bueno, A.; Carvalho, F.B.; Gutierres, J.M.; Lhamas, C.L.; Brusco, I.; Oliveira, S.M.; Amaral, M.G.; Dorneles, G.; Sorraila, J.; Duarte, M.M. Impacts of dose and time of boldenone and stanazolol exposure in inflammatory markers, oxidative and nitrosative stress and histopathological changes in the rat testes. Theriogenology 2017, 90, 101–108. [Google Scholar] [CrossRef]
- Slott, V.L.; Suarez, J.D.; Perreault, S.D. Rat sperm motility analysis: Methodologic considerations. Reprod. Toxicol. 1991, 5, 449–458. [Google Scholar]
- Robb, G.; Amann, R.; Killian, G. Daily sperm production and epididymal sperm reserves of pubertal and adult rats. Reproduction 1978, 54, 103–107. [Google Scholar]
- Filler, R. Methods for evaluation of rat epididymal sperm morphology. In Methods in Toxicology: Male Reproductive Toxicology; Chapin, R.E., Heildel, J.J., Eds.; Academic Press: San Diego, CA, USA, 1993; pp. 334–343. [Google Scholar]
- Zirkin, B.R.; Chen, H. Regulation of Leydig cell steroidogenic function during aging. Biol. Reprod. 2000, 63, 977–981. [Google Scholar] [PubMed]
- Sinha, A.K. Colorimetric assay of catalase. Anal. Biochem. 1972, 47, 389–394. [Google Scholar] [PubMed]
- Nishikimi, M.; Rao, N.A.; Yagi, K. The occurrence of superoxide anion in the reaction of reduced phenazine methosulfate and molecular oxygen. Biochem. Biophys. Res. Commun. 1972, 46, 849–854. [Google Scholar] [PubMed]
- Beutler, E.; Duron, O.; Kelly, B.M. Improved method for the determination of blood glutathione. J. Lab. Clin. Med. 1963, 61, 882–888. [Google Scholar]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar]
- Kittel, B.; Ruehl-Fehlert, C.; Morawietz, G.; Klapwijk, J.; Elwell, M.R.; Lenz, B.; O’Sullivan, M.G.; Roth, D.R.; Wadsworth, P.F. Revised guides for organ sampling and trimming in rats and mice–Part 2: A joint publication of the RITA and NACAD groups. Exp. Toxicol. Pathol. 2004, 55, 413–431. [Google Scholar] [PubMed]
- Bancroft, J.; Suvarna, S.; Layton, C. Bancroft’s Theory and Practice of Histological Techniques-Immunohistochemical and Immunofluorescent Techniques; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Hsu, S.-M.; Raine, L.; Fanger, H. Use of avidin-biotin-peroxidase complex (ABC) in immunoperoxidase techniques: A comparison between ABC and unlabeled antibody (PAP) procedures. J. Histochem. Cytochem. 1981, 29, 577–580. [Google Scholar] [PubMed]
- Saleh, N.; Waded, E. Immune response following the administration of the anabolic steroid Boldenone Undecylenate in rabbits. Stem Cell 2014, 5, 80e7. [Google Scholar]
- Arnold, A.; Peralta, J.; Thonney, M. Ontogeny of growth hormone, insulin-like growth factor-I, estradiol and cortisol in the growing lamb: Effect of testosterone. J. Endocrinol. 1996, 150, 391–399. [Google Scholar]
- de Souza, G.L.; Hallak, J. Anabolic steroids and male infertility: A comprehensive review. BJU Int. 2011, 108, 1860–1865. [Google Scholar] [PubMed]
- Kehinde, O.S.; Christianah, O.I.; Oyetunji, O.A. Ascorbic acid and sodium benzoate synergistically aggravates testicular dysfunction in adult Wistar rats. Int. J. Physiol. Pathophysiol. Pharmacol. 2018, 10, 39. [Google Scholar]
- Al-Sagheer, A.A.; Daader, A.H.; Gabr, H.A.; Abd El-Moniem, E.A. Palliative effects of extra virgin olive oil, gallic acid, and lemongrass oil dietary supplementation on growth performance, digestibility, carcass traits, and antioxidant status of heat-stressed growing New Zealand White rabbits. Environ. Sci. Pollut. Res. 2017, 24, 6807–6818. [Google Scholar]
- Alsaht, A.A.; Bassiony, S.M.; Abdel-Rahman, G.A.; Shehata, S.A. Effect of cinnamaldehyde thymol mixture on growth performance and some ruminal and blood constituents in growing lambs fed high concentrate diet. Life Sci. J. 2014, 11, 240–248. [Google Scholar]
- Cannizzo, F.T.; Zancanaro, G.; Spada, F.; Mulasso, C.; Biolatti, B. Pathology of the testicle and sex accessory glands following the administration of boldenone and boldione as growth promoters in veal calves. J. Vet. Med. Sci. 2007, 69, 1109–1116. [Google Scholar] [PubMed]
- Fronczak, C.M.; Kim, E.D.; Barqawi, A.B. The insults of illicit drug use on male fertility. J. Androl. 2012, 33, 515–528. [Google Scholar]
- Kim, E.D.; Crosnoe, L.; Bar-Chama, N.; Khera, M.; Lipshultz, L.I. The treatment of hypogonadism in men of reproductive age. Fertil. Steril. 2013, 99, 718–724. [Google Scholar]
- Elmasry, T.A.; Al-Shaalan, N.H.; Tousson, E.; El-Morshedy, K.; Al-Ghadeer, A. Star anise extracts modulation of reproductive parameters, fertility potential and DNA fragmentation induced by growth promoter Equigan in rat testes. Braz. J. Pharm. Sci. 2018, 54. [Google Scholar] [CrossRef]
- Paoli, D.; Gallo, M.; Rizzo, F.; Baldi, E.; Francavilla, S.; Lenzi, A.; Lombardo, F.; Gandini, L. Mitochondrial membrane potential profile and its correlation with increasing sperm motility. Fertil. Steril. 2011, 95, 2315–2319. [Google Scholar]
- Nakada, K.; Sato, A.; Yoshida, K.; Morita, T.; Tanaka, H.; Inoue, S.-I.; Yonekawa, H.; Hayashi, J.-I. Mitochondria-related male infertility. Proc. Natl. Acad. Sci. USA 2006, 103, 15148–15153. [Google Scholar] [CrossRef] [PubMed]
- Juan, M.; Alicia, P.; Oriana, B.; Bing, Z.; Roberto, S.; David, W. Vitamin C is a kinase inhibitor. Mol. Cell Biol. 2004, 24, 6645–6652. [Google Scholar]
- Nair, S.; Rajamohan, T. The Role of Coconut Water on Nicotine-Induced Reproductive Dysfunction in Experimental Male Rat Model. Food Nutr. Sci. 2014, 5, 1121–1130. [Google Scholar] [CrossRef]
- Barrett, K.E.; Barman, S.M.; Boitano, S.; Brooks, H.L. Ganong’s Review of Medical Physiology; McGraw-Hill Education: New York, NY, USA, 2016. [Google Scholar]
- Guyton, A.C.; Hall, J.E. Textbook of Medical Physiology, 10th ed.; Saunders: Phliadelphia, PA, USA, 2000. [Google Scholar]
- Tatem, A.J.; Beilan, J.; Kovac, J.R.; Lipshultz, L.I. Management of anabolic steroid-induced infertility: Novel strategies for fertility maintenance and recovery. World J. Men’s Health 2020, 38, 141–150. [Google Scholar] [CrossRef]
- Ishak, M.; Omar, C. Effect of Testosterone Enanthate Injection and Castration on some Productive Traits of Karadi Lambs. J. Zankoy Sulaimani Part A 2013, 16, 219–230. [Google Scholar] [CrossRef]
- Rasul, K.H.; Aziz, F.M. The Effect of Sustanon (Testosterone Derivatives) Taken by Athletes on the Testis of Rat. Jordan J. Biol. Sci. 2012, 5, 113–119. [Google Scholar]
- Borges, T.; Eisele, G.; Byrd, C. Review of Androgenic Anabolic Steroid Use; Oak Ridge Institute for Science and Education: Oak Ridge, TN, USA, 2001. [Google Scholar]
- Llewellyn, W. Anabolics; Molecular Nutrition LLC: Jupiter, FL, USA, 2011. [Google Scholar]
- Hayes, F.J.; Seminara, S.B.; DeCruz, S.; Boepple, P.A.; Crowley, W.F., Jr. Aromatase inhibition in the human male reveals a hypothalamic site of estrogen feedback. J. Clin. Endocrinol. Metab. 2000, 85, 3027–3035. [Google Scholar] [PubMed]
- Ghazy, E.J.; Elballal, S.S. Effect of the anabolic steroid, boldenone undecylenate on reproductive performance of male rabbits. J. Reprod. Infertil. 2010, 50, 8–17. [Google Scholar]
- Anvari, M.; Pourentezari, M.; Talebi, A.; Mangoli, M.; Khalili, M. The improvement of sperm parameters and chromatin quality by vitamin E supplement in mice. Researcher 2012, 4, 43–49. [Google Scholar]
- Ali, E.; Tousson, E.; el Daim, H. Effects of short and long term growth promoter boldenone undecylenate treatments on antioxidant enzyme activities and oxidative stress markers in rabbit muscles. Pharmacologia 2013, 4, 576–581. [Google Scholar] [CrossRef]
- Ibrahim, S.S.; Said, A.M. Effect of lycopene in amelioration of testicular and renal toxicity induced by boldenone undecylenate in male albino rats. Int. J. Med. Biomed. Stud. 2019, 3. [Google Scholar] [CrossRef]
- Pinheiro, J.L.; Maia, B.P.; Lima, A.B.d.; Domingues, R.J.d.S.; Oliveira, F.R.T.; Freitas, J.J.d.S.; Kietzer, K.S. Nandrolone decanoate is prooxidant in the myocardium of exercised or sedentary rats. Rev. Bras. Med. Esporte 2020, 26, 16–20. [Google Scholar] [CrossRef]
- Mangoli, E.; Talebi, A.R.; Anvari, M.; Taheri, F.; Vatanparast, M.; Rahiminia, T.; Hosseini, A. Vitamin C attenuates negative effects of vitrification on sperm parameters, chromatin quality, apoptosis and acrosome reaction in neat and prepared normozoospermic samples. Taiwan. J. Obstet. Gynecol. 2018, 57, 200–204. [Google Scholar] [CrossRef] [PubMed]
- Levine, M.; Ebenuwa, I.; Violet, P.-C. Chapter 17—Vitamin C. In Essential and Toxic Trace Elements and Vitamins in Human Health; Prasad, A.S., Brewer, G.J., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 241–262. [Google Scholar] [CrossRef]
- Kini, R.D.; Tripathi, Y.B.; Raghuveer, C.V.; Pai, S.; Ramaswamy, C.; Kamath, P.R. Role of vitamin C as an antioxidant in cadmium chloride induced testicular damage. Int. J. Appl. Biol. Pharm. 2011, 2, 484–488. [Google Scholar]
- O’Hara, L.; Smith, L.B. Androgen receptor roles in spermatogenesis and infertility. Best Pract. Res. Clin. Endocrinol. Metab. 2015, 29, 595–605. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.-J.; Hardy, M.P.; Inigo, I.V.; Huhtaniemi, I.; Bardin, C.W.; Moo-Young, A.J. Effects of androgen on androgen receptor expression in rat testicular and epididymal cells: A quantitative immunohistochemical study. Biol. Reprod. 2000, 63, 368–376. [Google Scholar] [CrossRef]
- Birgner, C.; Kindlundh-Högberg, A.M.; Alsiö, J.; Lindblom, J.; Schiöth, H.B.; Bergström, L. The anabolic androgenic steroid nandrolone decanoate affects mRNA expression of dopaminergic but not serotonergic receptors. Brain Res. 2008, 1240, 221–228. [Google Scholar] [CrossRef]
- Rebourcet, D.; O’Shaughnessy, P.J.; Smith, L.B. The expanded roles of Sertoli cells: Lessons from Sertoli cell ablation models. Curr. Opin. Endocr. Metab. Res. 2019, 6, 42–48. [Google Scholar] [CrossRef]
- Watanabe, N. Decreased number of sperms and Sertoli cells in mature rats exposed to diesel exhaust as fetuses. Toxicol. Lett. 2005, 155, 51–58. [Google Scholar] [CrossRef]
- Bahey, N.G.; Soliman, G.M.; El-Deeb, T.A.; El-Drieny, E.A. Influence of insulin and testosterone on diabetic rat ventral prostate: Histological, morphometric and immunohistochemical study. J. Microsc. Ultrastruct. 2014, 2, 151–160. [Google Scholar]
- Fritz, I.B. Somatic cell-germ cell relationships in mammalian testes during development and spermatogenesis. Ciba Found. Symp. 1994, 182, 271–274. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Control | Vehicle Control | Vit-C | BOL | BOL+Vit-C | p-Value |
|---|---|---|---|---|---|---|
| Initial body weight (g) | 165.33 ± 2.31 | 166.00 ± 1.76 | 168.75 ± 1.50 | 165.50 ± 1.43 | 168.50 ±1.50 | 0.465 |
| Final body weight (g) | 204.93 d ± 4.47 | 205.25 d ± 1.20 | 220.25 c ± 1.59 | 247.00 b ± 1.84 | 275.75 a ± 5.89 | <0.001 |
| Body weight gain (g) | 39.60 c ± 4.86 | 39.25 c ± 1.98 | 51.50 c ± 2.64 | 81.50 b ± 2.50 | 107.25 a ± 7.15 | <0.001 |
| Gonadosomatic index (%) | 1.00a ± 0.05 | 0.89ab ± 0.05 | 1.01 a ± 0.05 | 0.73c ± 0.03 | 0.79 bc ± 0.02 | <0.001 |
| Sperm count (sp.cc/mL×125×104) | 37.67 a ± 1.31 | 38.00 a ± 1.08 | 40.00 a ± 1.22 | 16.00b ± 1.47 | 17.33b± 1.25 | <0.001 |
| Sperm motility (%) | 87.00 a ± 1.08 | 86.33 a ± 1.43 | 89.33 a ± 2.09 | 60.00 c ± 2.04 | 79.67 b ± 1.84 | <0.001 |
| Sperm abnormalities (%) | 17.24 c ± 0.32 | 17.26 c ± 0.27 | 15.58 c ± 0.50 | 40.82a ± 0.47 | 24.26 b ± 1.31 | <0.001 |
| Groups | Control | Vehicle Control | Vit-C | BOL | BOL+Vit-C | p-Value |
|---|---|---|---|---|---|---|
| Serum TES (ng/mL) | 2.87 b ± 0.31 | 3.65 b ± 0.23 | 3.88 b ± 0.75 | 7.19 a ± 0.08 | 6.12 a ± 0.79 | <0.001 |
| Testicular TES (ng/g tissue) | 406.40a ± 9.86 | 400.00a ± 13.76 | 400.00 a ± 14.71 | 147.20 b ± 5.99 | 171.20b ± 8.16 | <0.001 |
| LH (mIU/mL) | 3.43 a ± 0.08 | 3.51 a ± 0.06 | 3.53 a ± 0.06 | 2.28 b ± 0.32 | 2.67 b ± 0.15 | <0.001 |
| FSH (ng/mL) | 8.31 a ± 0.39 | 8.35 a ± 0.23 | 8.45 a ± 0.29 | 4.54 b ± 0.50 | 5.63 b ± 0.38 | <0.001 |
| E2 (pg/mL) | 59.77 a ± 3.20 | 58.10 a ± 3.34 | 57.67 a ± 3.14 | 28.20 b ± 1.28 | 35.13b ± 1.82 | <0.001 |
| ACP (U/L) | 15.67 c ± 1.65 | 16.67c ± 1.43 | 16.33 c ± 1.25 | 47.33 a ± 2.72 | 25.67 b ± 2.46 | <0.001 |
| SDH (ng/mL) | 1.38 c ± 0.07 | 1.39 c ± 0.19 | 1.38 c ± 0.10 | 5.00 a ± 0.14 | 2.36 b ± 0.26 | <0.001 |
| Lesion | Control | Vehicle Control | Vit-C | BOL | Vit-C+BOL | p-Value |
|---|---|---|---|---|---|---|
| ST/10X | 15.40 b ± 0.69 | 15.90 b ± 0.67 | 15.50 b ± 0.72 | 24.60 a ± 0.70 | 23.00 a ± 0.61 | <0.001 |
| Diameter of ST | 275.09 a ± 6.22 | 275.58 a ± 6.07 | 279.05 a ± 5.80 | 175.23 b ± 11.33 | 184.41 b ± 10.81 | <0.001 |
| Height of germinal epithelium/ST | 83.18 a ± 1.68 | 83.54 a ± 1.58 | 83.81 a ± 2.09 | 44.04 b ± 3.08 | 46.26 b ± 2.26 | <0.001 |
| ST with vacuolated germinal epithelium | 0.00 c ± 0.00 | 0.00 c ± 0.00 | 0.00 c ± 0.00 | 5.66 a ± 0.73 | 4.05 b± 0.23 | <0.001 |
| ST with detached germinal epithelium | 0.12 c ± 0.12 | 0.13 c ± 0.13 | 0.13 c ± 0.13 | 18.10 a ± 1.22 | 14.54 b ± 1.96 | <0.001 |
| STs with depleted germ cells | 0.00 c ± 0.00 | 0.00 c ± 0.00 | 0.00 c ± 0.00 | 64.89 a ± 3.73 | 57.86 b ± 3.60 | <0.001 |
| ST with necrotic germinal epithelium | 0.00 c ± 0.00 | 0.00 c ± 0.00 | 0.00 c ± 0.00 | 58.65 a ± 2.39 | 50.07 b ± 1.67 | <0.001 |
| STs with complete loss of germ cells | 0.00 c ± 0.00 | 0.00 c ± 0.00 | 0.00 c ± 0.00 | 2.04 a ± 0.35 | 1.22 b ± 0.18 | <0.001 |
| STs with redundant basement membranes | 0.00 b ± 0.00 | 0.00 b ± 0.00 | 0.00 b ± 0.00 | 0.99 a ± 0.30 | 0.45 b ± 0.25 | <0.001 |
| Spermatid retention | 0.00 c ± 0.00 | 0.00 c ± 0.00 | 0.00 c ± 0.00 | 0.65 a ± 0.02 | 0.32 b ± 0.01 | <0.001 |
| Interstitial leukocytic infiltration | 0.00 c ± 0.00 | 0.00 c ± 0.00 | 0.00 c ± 0.00 | 4.10 a ± 0.80 | 2.05 b ± 0.40 | <0.001 |
| Interstitial edema | 0.00 c ± 0.00 | 0.00 c ± 0.00 | 0.00 c ± 0.00 | 8.00 a ± 0.73 | 4.00 b ± 0.37 | <0.001 |
| Interstitial congestion | 0.00 b ± 0.00 | 0.00 b ± 0.00 | 0.00 b ± 0.00 | 10.00 a ± 3.33 | 4.00 b ± 2.67 | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Behairy, A.; El-Sharkawy, N.I.; Saber, T.M.; Soliman, M.M.; Metwally, M.M.M.; Abd El-Rahman, G.I.; Abd-Elhakim, Y.M.; El Deib, M.M. The Modulatory Role of Vitamin C in Boldenone Undecylenate Induced Testicular Oxidative Damage and Androgen Receptor Dysregulation in Adult Male Rats. Antioxidants 2020, 9, 1053. https://doi.org/10.3390/antiox9111053
Behairy A, El-Sharkawy NI, Saber TM, Soliman MM, Metwally MMM, Abd El-Rahman GI, Abd-Elhakim YM, El Deib MM. The Modulatory Role of Vitamin C in Boldenone Undecylenate Induced Testicular Oxidative Damage and Androgen Receptor Dysregulation in Adult Male Rats. Antioxidants. 2020; 9(11):1053. https://doi.org/10.3390/antiox9111053
Chicago/Turabian StyleBehairy, Amany, Nabela I. El-Sharkawy, Taghred M. Saber, Mohamed Mohamed Soliman, Mohamed M.M. Metwally, Ghada I. Abd El-Rahman, Yasmina M. Abd-Elhakim, and Maha M. El Deib. 2020. "The Modulatory Role of Vitamin C in Boldenone Undecylenate Induced Testicular Oxidative Damage and Androgen Receptor Dysregulation in Adult Male Rats" Antioxidants 9, no. 11: 1053. https://doi.org/10.3390/antiox9111053
APA StyleBehairy, A., El-Sharkawy, N. I., Saber, T. M., Soliman, M. M., Metwally, M. M. M., Abd El-Rahman, G. I., Abd-Elhakim, Y. M., & El Deib, M. M. (2020). The Modulatory Role of Vitamin C in Boldenone Undecylenate Induced Testicular Oxidative Damage and Androgen Receptor Dysregulation in Adult Male Rats. Antioxidants, 9(11), 1053. https://doi.org/10.3390/antiox9111053