Liver Antioxidants in Relation to Beak Morphology, Gizzard Size and Diet in the Common Eider Somateria mollissima


Abstract
1. Introduction
2. Materials and Methods
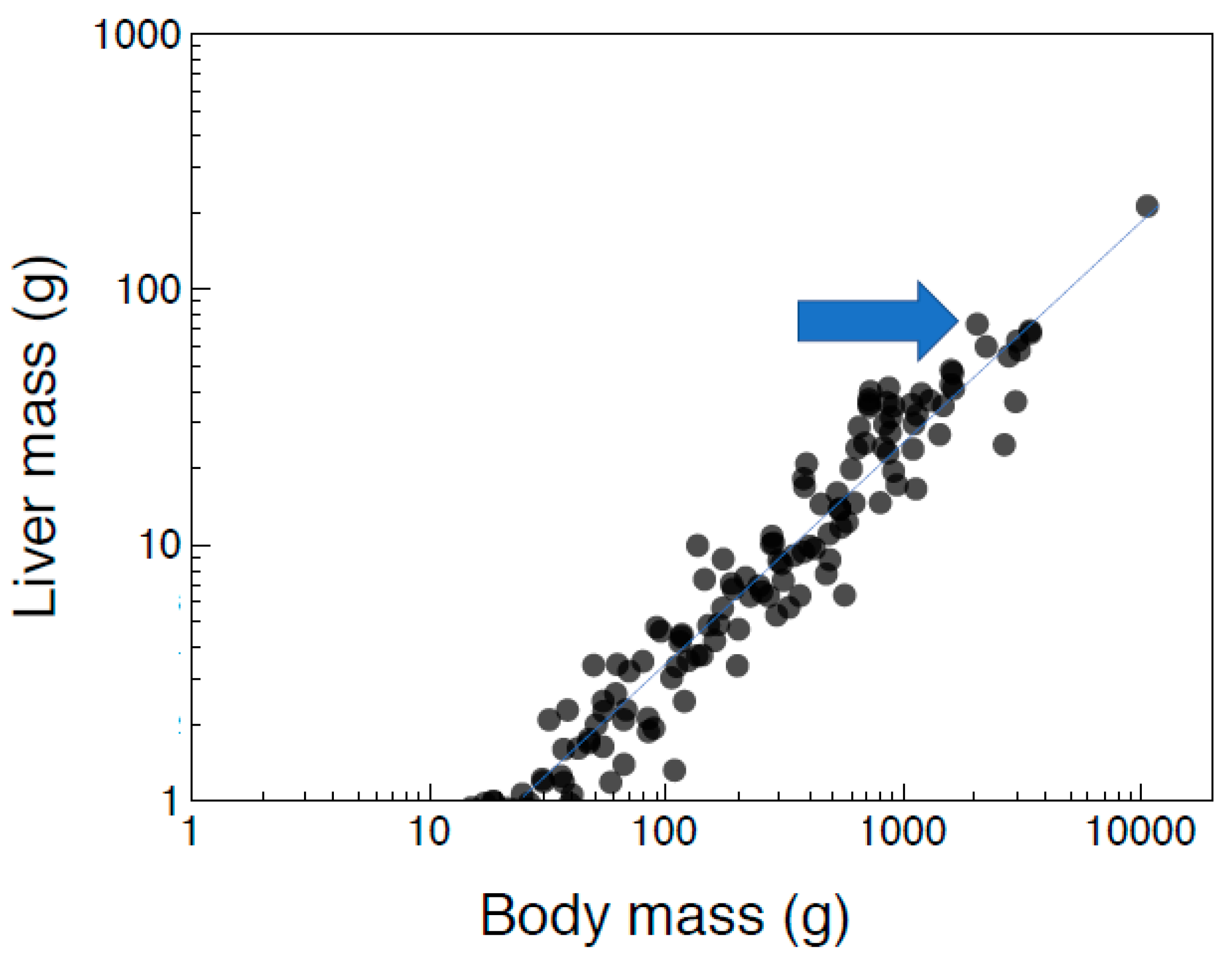
2.1. Liver Mass and Body Mass in Different Species of Birds
2.2. Study Sites and Study Samples
2.3. Sample Extraction and HPLC Analysis
2.4. Statistical Analyses
3. Results
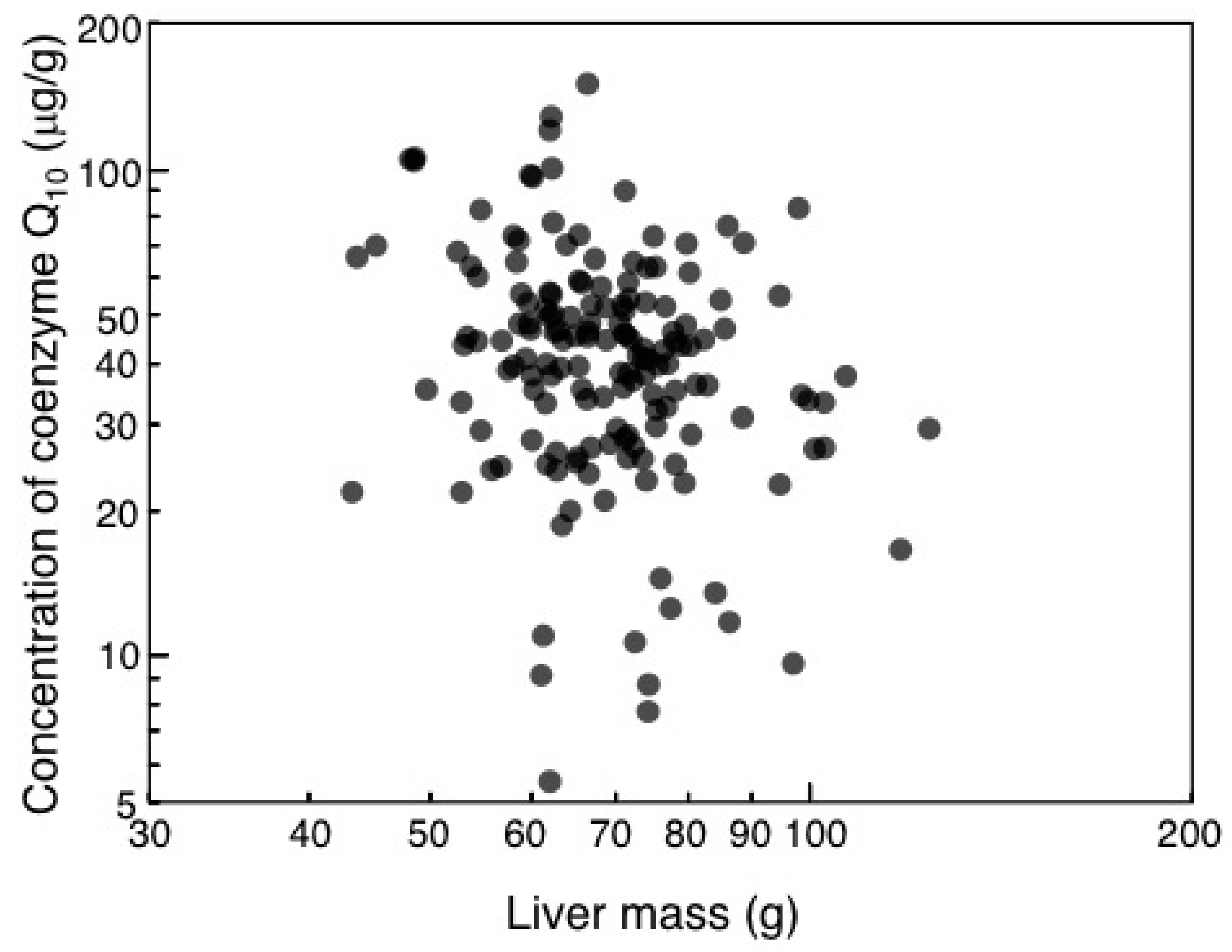
3.1. Interspecific Allometry of Liver Antioxidants
3.2. Liver Antioxidants and Ecology of Common Eiders
3.3. Antioxidants, Date and Age
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Halliwell, B.; Gutteridge, J.M. Free Radicals in Biology and Medicine; Oxford University Press: Oxford, UK, 2007. [Google Scholar]
- Surai, P.F. Natural Antioxidants in Avian Nutrition and Reproduction; Nottingham University Press: Nottingham, UK, 2002. [Google Scholar]
- Klasing, K.C. Comparative Avian Nutrition; Cab International: London, UK, 2000. [Google Scholar]
- Meijer, T.; Drent, R. Re-examination of the capital and income dichotomy in breeding birds. Ibis 1999, 141, 399–414. [Google Scholar] [CrossRef]
- Blums, P.; Nichols, J.; Hines, J.; Lindberg, M.; Mednis, A. Individual quality, survival variation and patterns of phenotypic selection on body condition and timing of nesting in birds. Oecologia 2005, 143, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Rubolini, D.; Romano, M.; Bonisoli Alquati, A.; Saino, N. Early maternal, genetic and environmental components of antioxidant protection, morphology and immunity of yellow-legged gull (Larus michahellis) chicks. Eur. J. Evol. Biol. 2006, 19, 1571–1584. [Google Scholar] [CrossRef] [PubMed]
- Pappas, A.C.; Karadas, F.; Surai, P.F.; Wood, N.A.R.; Cassey, P.; Bortolotti, G.R.; Speake, B.K. Interspecies variation in yolk selenium concentrations among eggs of free-living birds: The effect of phylogeny. J. Trace Elem. Med. Biol. 2006, 20, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Karadas, F.; Pappas, A.C.; Surai, P.F.; Speake, B.K. Embryonic development within carotenoid-enriched eggs influences the post-hatch carotenoid status of the chicken. Comp. Biochem. Physiol. Part B 2005, 244–251. [Google Scholar] [CrossRef]
- Karadas, F.; Surai, P.F.; Grammenidis, E.; Sparks, N.H.; Acamovic, T. Supplementation of the maternal diet with tomato powder and marigold extract: Effects on the antioxidant system of the developing quail. Br. Poult. Sci. 2006, 200–208. [Google Scholar] [CrossRef]
- Karadas, F.; Surai, P.F.; Grammenidis, E.; Sparks, N.H.C. Effects of maternal dietary supplementation with three sources of carotenoids on the retinyl esters of egg yolk and developing quail liver. Comp. Biochem. Physiol. Part B 2006, 140, 430–435. [Google Scholar] [CrossRef]
- Surai, P.; Noble, R.C.; Speake, B. Tissue-specific differences in antioxidant distribution and susceptibility to lipid peroxidation during development of the chick embryo. Biochem. Biochys. Acta 1996, 1304, 1–10. [Google Scholar] [CrossRef]
- Blount, J.D.; Houston, D.C.; Surai, P.F.; Møller, A.P. Egg-laying capacity is limited by carotenoid pigment availability in wild gulls Larus fuscus. Biol. Lett. 2003, 271, S79–S81. [Google Scholar] [CrossRef]
- Biard, C.; Surai, P.F.; Møller, A.P. Effects of carotenoid availability during laying on reproduction in the blue tit. Oecologia 2005, 144, 32–44. [Google Scholar] [CrossRef]
- Surai, P.F. Selenium-vitamin E interactions: Does 1+1 equal more than 2? In Nutritional Biotechnology in the Feed and Food Industries Proceedings of Alltech’s 19th Annual Symposium; Lyons, T.P., Jacques, K.A., Eds.; Nottingham University Press: Nottingham, UK, 2003; pp. 59–76. [Google Scholar]
- Sunder, A.; Flachowsky, G. Influence of high vitamin E dosages on retinol and carotenoid concentration in body tissues and eggs of laying hens. Arch. Tierernähr. 2001, 55, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Surai, P.F.; Sparks, N.H.C. Designer eggs: from improvement of egg composition to functional food. Trends Food Sci. Technol. 2001, 12, 7–16. [Google Scholar] [CrossRef]
- Koutsos, E.A.; Clifford, A.J.; Calvert, C.C.; Klasing, K. Maternal carotenoid status modifies the incorporation of dietary carotenoids into immune tissues of growing chickens (Gallus gallus domesticus). J. Nutr. 2003, 133, 1132–1138. [Google Scholar] [CrossRef]
- Surai, P.F.; Speake, B.K.; Sparks, N.H.C. Carotenoids in avian nutrition and embryonic development: 1. Absorption, availability and levels in plasma and egg yolk. J. Poult. Sci. 2001, 38, 1–27. [Google Scholar] [CrossRef]
- Surai, P.F.; Speake, B.K.; Sparks, N.H.C. Carotenoids in avian nutrition and embryonic development: 2. Antioxidant properties and discrimination in embryonic tissues. J. Poult. Sci. 2001, 38, 117–145. [Google Scholar]
- Saino, N.; Ferrari, R.P.; Romano, M.; Martinelli, R.; Møller, A.P. Experimental manipulation of egg carotenoids affects immunity of barn swallow nestlings. Proc. R. Soc. Lond. B 2003, 270, 2485–2489. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.F.; Chang, S.J.; Hsu, A.L. Effects of supplemental vitamin E during the laying period on the reproductive performance of Taiwan native chickens. Br. Poult. Sci. 2004, 45, 807–814. [Google Scholar] [CrossRef]
- Lenaz, G.; Fato, R.; Formiggini, G.; Genova, M.L. The role of Coenzymes Q in mitochondrial electron transport. Mitochondrion 2007, 7, S8–S33. [Google Scholar] [CrossRef]
- Gopi, M.; Purushothaman, M.R.; Chandrasekaran, D. Effect of dietary coenzyme Q10 supplementation on the growth rate, carcass characters and cost effectiveness of broiler fed with three energy levels. Springer Plus 2014, 3, 518. [Google Scholar] [CrossRef]
- Bentinger, M.; Brismar, K.; Dallner, G. The antioxidant role of coenzyme Q. Mitochondrion 2007, 7, S41–S50. [Google Scholar] [CrossRef]
- Geng, A.L.; Guo, Y.M.; Yang, Y. Reduction of ascites mortality in broilers by coenzyme Q10. Poult. Sci. 2010, 83, 1587–1593. [Google Scholar] [CrossRef] [PubMed]
- Forstmark-Andre, P.; Lee, C.P.; Dallner, G.; Ernster, L. Lipid peroxidation and changes in the ubiquinone content and the respiratory chain enzymes of submitochondrial particles. Free Radic. Biol. Med. 1997, 22, 391–400. [Google Scholar] [CrossRef]
- Tang, P.H.; Miles, M.V.; Degrauw, A.; Hershey, A.; Pesce, A. HPLC analysis of reduced and oxidized coenzyme Q10 in human plasma. Clin. Chem. 2001, 47, 256–265. [Google Scholar] [PubMed]
- Navas, P.; Villalba, J.M.; Cabo, R.D. The importance of plasma membrane coenzyme Q in aging and stress responses. Mitochondrion 2007, 7S, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Krizman, B.J.; Prosek, M.; Smidovnik, A.; Wondra, A.G.; Glaser, R.; Zelenko, B.; Volk, M. Poultry products with increased content of CoQ10 prepared from chickens fed with supplemental CoQ10. In Trends in Vital Food and Control Engineering; Eissa, A.A., Ed.; InTech: Rijeka, Croatia, 2012; Chapter 9; pp. 165–186. [Google Scholar]
- Nehls, G.; Ketzenberg, C. Do common Eiders Somateria mollissima exhaust their food resources? A study on natural mussel Mytilus edulis beds in the Wadden Sea. Dan. Rev. Game Biol. 2002, 16, 47–61. [Google Scholar]
- Laursen, K.; Frikke, J. Hunting from motorboats displaces Wadden Sea Eiders Somateria mollissima from their favoured feeding distribution. Wildl. Biol. 2008, 14, 423–433. [Google Scholar] [CrossRef]
- Laursen, K.; Asferg, K.; Frikke, J.; Sunde, P. Mussel fishery affects diet and reduces body condition of Eiders Somateria mollissima in the Wadden Sea. J. Sea Res. 2009, 62, 22–30. [Google Scholar] [CrossRef]
- Cramp, S. Handbook of the Birds of Europe, the Middle East and North Africa; Oxford University Press: Oxford, UK, 1977; Volume 1. [Google Scholar]
- Surai, P.F.; Speake, B.K.; Decrock, F.; Groscolas, R. Transfer of Vitamins E and A from yolk to embryo during development of the King Penguin. Physiol. Biochem. Zool. 2001, 74, 928–936. [Google Scholar] [CrossRef]
- Karadas, F.; Surai, P.F.; Sparks, N.H.C. Changes in broiler chick tissue concentrations of lipid-soluble antioxidants immediately post-hatch. Comp. Biochem. Physiol. A 2011, 160, 68–71. [Google Scholar] [CrossRef]
- Mattila, P.; Kumpulainen, J. Coenzymes Q9 and Q10: Contents in foods and dietary intake. J. Food Comp. Anal. 2001, 14, 409–417. [Google Scholar] [CrossRef]
- McClave, J.T.; Sincich, T. Statistics, 9th ed.; Prentice-Hall: Upper Saddle River, NJ, USA, 2003. [Google Scholar]
- SAS. JMP Version 10.2; SAS Institute, Inc.: Cary, NC, USA, 2012. [Google Scholar]
- Hespenheide, H.A. Ecological inferences from morphological data. Ann. Rev. Ecol. Syst. 1973, 4, 213–229. [Google Scholar] [CrossRef]
- Lederer, R.J. Bill size, food size, and jaw forces of insectivorous birds. Auk 1973, 92, 385–387. [Google Scholar] [CrossRef]
- Wilson, D.S. The adequacy of body size as a niche difference. Am. Nat. 1975, 109, 769–784. [Google Scholar] [CrossRef]
- Herrel, A.; Podos, J.; Huber, S.K.; Hendry, A.P. Evolution of bite force in Darwin’s finches: A key role for head width. J. Evol. Biol. 2005, 18, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Karadas, F.; Pirgozliev, V.; Pappas, A.C.; Acamovic, T.; Bedford, M.R. Effects of different dietary phytase activities on the concentration of antioxidants in the liver of growing broilers. J. Anim. Physiol. Anim. Nutr. 2010, 94, 519–526. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Variable | Mean for Eider | SE | N | Mean for All Other Species | SE | N |
|---|---|---|---|---|---|---|
| Liver mass (g) | 70.117 | 1.065 | 162 | 12.494 | 1.617 | 179 |
| Total carotenoids | 4.295 | 0.142 | 162 | 44.134 | 5.408 | 36 |
| vitamin A | 1.539 | 0.113 | 162 | 1.751 | 0.202 | 36 |
| vitamin E | 2.900 | 0.201 | 162 | 120.327 | 12.500 | 36 |
| Coenzyme Q10 | 45.160 | 1.883 | 162 |
| Liver Mass | ||||||
|---|---|---|---|---|---|---|
| Liver mass | 1.000 | Body mass | ||||
| Body mass | 0.270 | 1.000 | Vitamin A | |||
| Vitamin A | 0.085 | 0.088 | 1.000 | Vitamin E | ||
| Vitamin E | 0.106 | 0.098 | 0.658 | 1.000 | Total carotenoids | |
| Total carotenoids | −0.013 | 0.121 | 0.123 | 0.525 | 1.000 | Coenzyme Q10 |
| Coenzyme Q10 | −0.268 | −0.096 | 0.072 | 0.291 | 0.391 | 1.000 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Møller, A.P.; Laursen, K.; Karadas, F. Liver Antioxidants in Relation to Beak Morphology, Gizzard Size and Diet in the Common Eider Somateria mollissima. Antioxidants 2019, 8, 31. https://doi.org/10.3390/antiox8020031
Møller AP, Laursen K, Karadas F. Liver Antioxidants in Relation to Beak Morphology, Gizzard Size and Diet in the Common Eider Somateria mollissima. Antioxidants. 2019; 8(2):31. https://doi.org/10.3390/antiox8020031
Chicago/Turabian StyleMøller, Anders Pape, Karsten Laursen, and Filiz Karadas. 2019. "Liver Antioxidants in Relation to Beak Morphology, Gizzard Size and Diet in the Common Eider Somateria mollissima" Antioxidants 8, no. 2: 31. https://doi.org/10.3390/antiox8020031
APA StyleMøller, A. P., Laursen, K., & Karadas, F. (2019). Liver Antioxidants in Relation to Beak Morphology, Gizzard Size and Diet in the Common Eider Somateria mollissima. Antioxidants, 8(2), 31. https://doi.org/10.3390/antiox8020031