Effect of Natural Food Antioxidants against LDL and DNA Oxidative Changes



Abstract
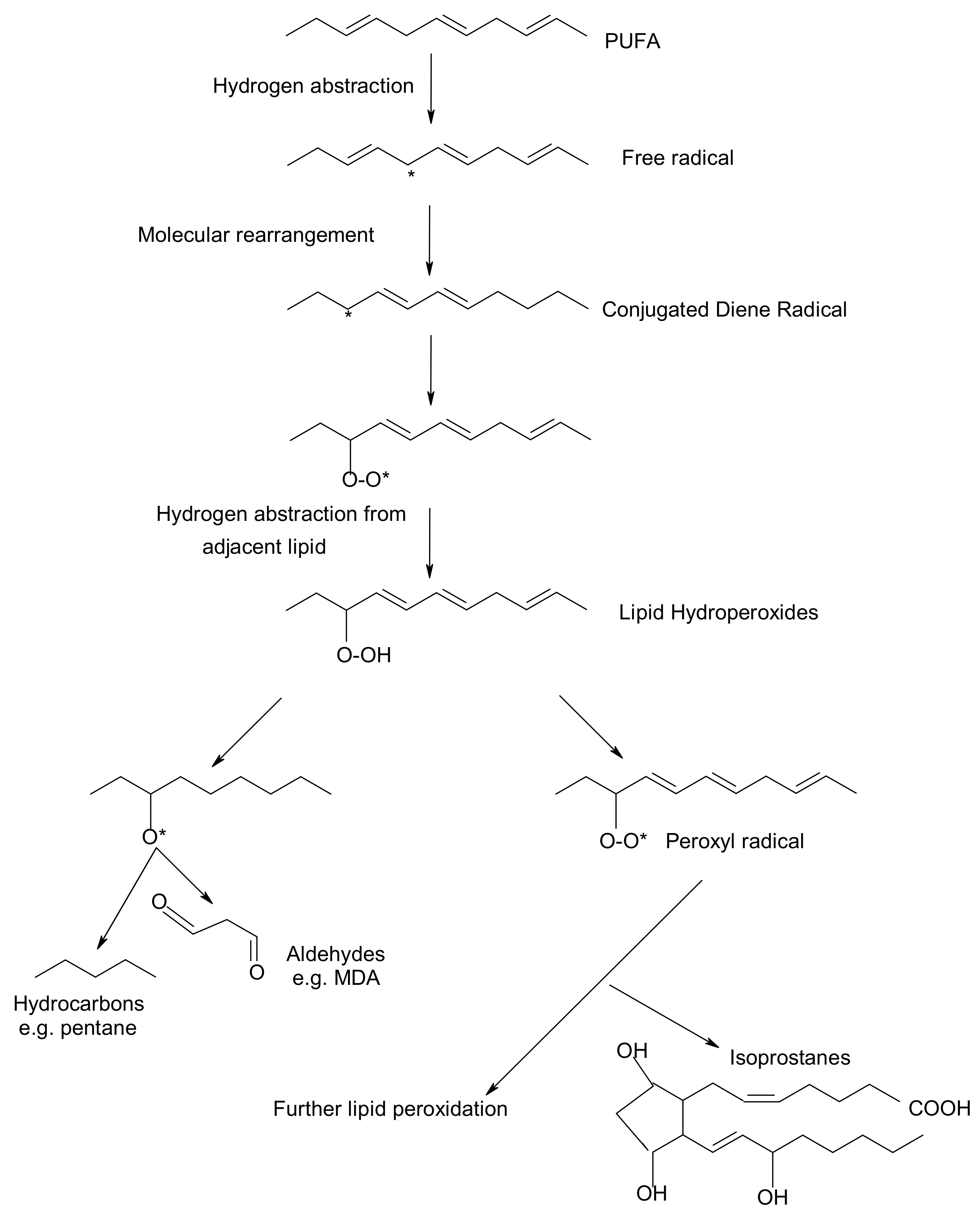
1. Introduction to Lipid Peroxidation and Antioxidants
2. Low-Density Lipoprotein (LDL) Oxidative Damage and Antioxidation
Link of LDL Oxidation to Atherogenesis and Common Monitoring Methods
3. Effect of Certain Natural Antioxidants against LDL Damage/Atherosclerosis
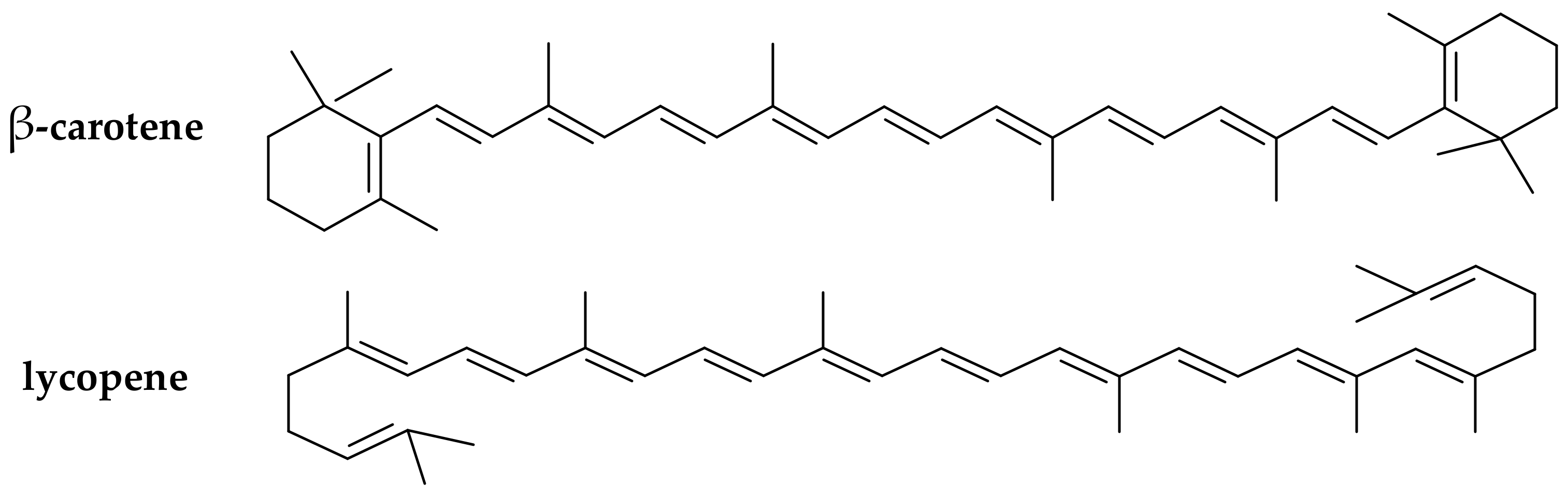
3.1. Effect of Carotenoids
3.2. Effect of Vitamin C
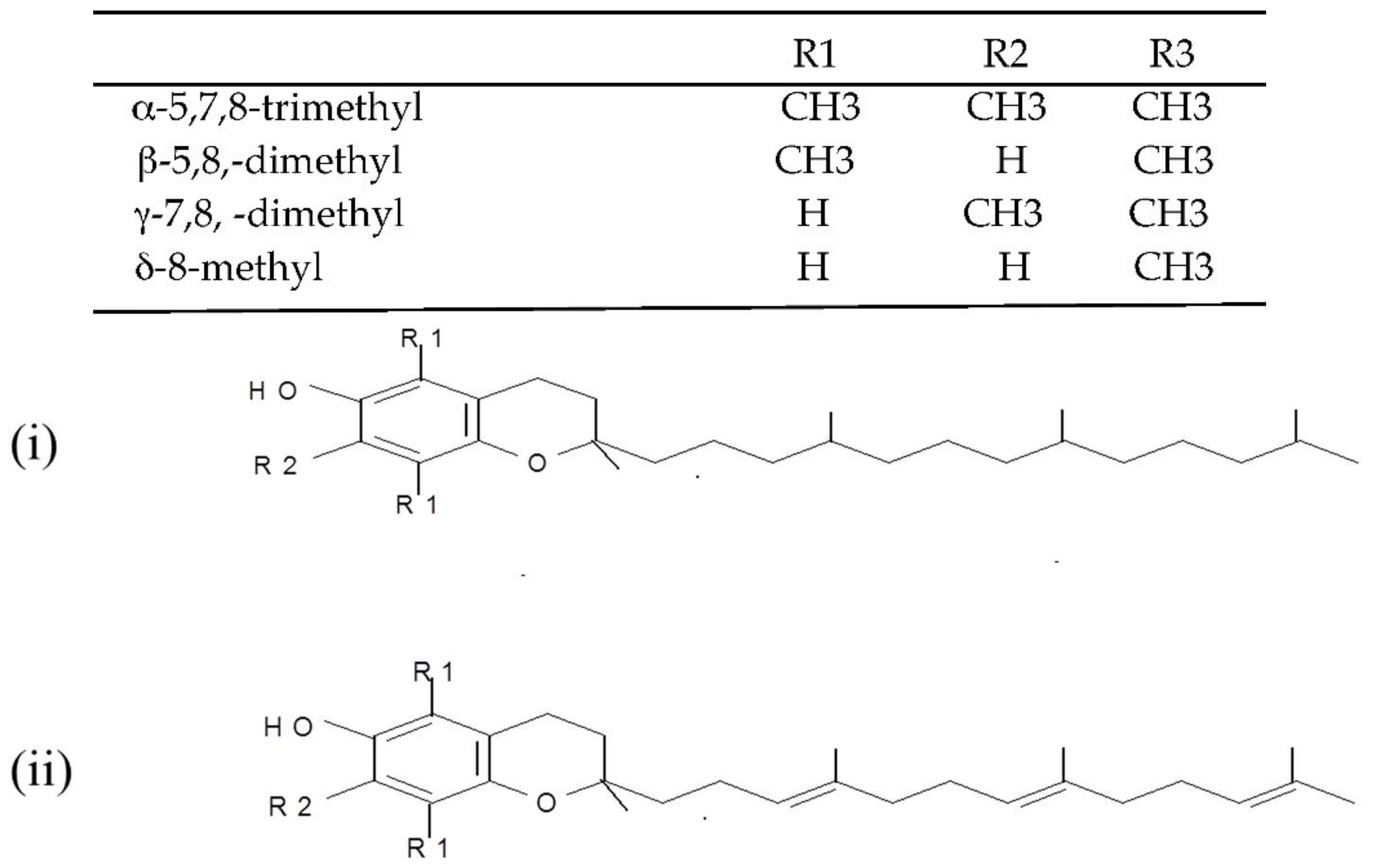
3.3. Effect of Vitamin E
3.4. Effect of Flavonoids, Phenolic Acids and Antioxidant Mixtures
4. DNA Oxidative Damage and Antioxidation
4.1. Link of DNA Oxidative Damage to Carcinogenesis and Common Monitoring Methods
4.2. Effects of Natural Antioxidants against DNA Damage/Carcinogenesis
4.2.1. Effect of Carotenoids
4.2.2. Effect of Vitamin C
4.2.3. Effect of Vitamin E
4.2.4. Effect of Flavonoids and Phenolic Acids
4.2.5. Effect of Antioxidant Mixtures
5. Main Conclusions/Future Work
Funding
Conflicts of Interest
References
- Pinchuk, I.; Shoval, Y.; Lichtenberg, D. Evaluation of antioxidants: Scope, limitations and relevance of assays. Chem. Phys. Lipids 2012, 165, 638–647. [Google Scholar] [CrossRef] [PubMed]
- Muller, F.-L.; Lustgarten, M.-S.; Jang, Y.; Richardson, A.; Van Remmen, H. Trends in oxidative aging theories. Free Radic. Biol. Med. 2007, 43, 477–503. [Google Scholar] [CrossRef] [PubMed]
- Dimakou, C.; Kiokias, S.; Tsaprouni, I.; Oreopoulou, V. Effect of processing and storage parameters on oxidative deterioration of oil-in-water emulsions. Food Biophys. 2007, 2, 38–45. [Google Scholar] [CrossRef]
- Lien, P.-H.; Hua, H.; Chuong, P.-H. Free radicals, antioxidants in disease and health. Int. J. Biomed. Sci. 2008, 4, 89–96. [Google Scholar]
- Preiser, J.-C. Oxidative stress. J. Parenter. Enter. Nutr. 2015, 36, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Lo, C.-J.; Edwards, T.-J.; Baran, S.-P. Radicals: Reactive intermediates with translational potential. J. Am. Chem. Soc. 2016, 138, 12692–12714. [Google Scholar] [CrossRef] [PubMed]
- Dizdaroglu, M.; Jaruga, P. Mechanisms of free radical-induced damage to DNA. Free Radic. Res. 2012, 46, 382–419. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; Yin, H. Role of lipid peroxidation derived 4-hydroxynonenal (4-HNE) in cancer: Focusing on mitochondria. Redox Biol. 2015, 4, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Kovacic, P.; Somanathan, R. Nanoparticles: Toxicity, radicals, electron transfer, and antioxidants. Methods Mol. Biol. 2013, 1028, 15–35. [Google Scholar] [PubMed]
- Dominguez, M.-G.; Castelao, E.-J. Role of lipid peroxidation and oxidative stress in the association between thyroid diseases and breast cancer. Crit. Rev. Oncol. Hematol. 2008, 68, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, A.; Chattopadhyay, R.; Mitra, S.; Sheila, E.; Crowe, S.-E. Oxidative Stress: An Essential Factor in the Pathogenesis of Gastrointestinal Mucosal Diseases. Physiol. Rev. 2014, 94, 329–354. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-J.; Kim, E.-H.; Hahn, K.-B. Oxidative stress in inflammation-based gastrointestinal tract diseases: Challenges and opportunities. J. Gastroenterol. Hepatol. 2012, 27, 1004–1010. [Google Scholar] [CrossRef] [PubMed]
- Van’t Erve, T.-J.; Lih, F.B.; Jelsema, C.; Deterding, L.-J.; Eling, T.-E.; Mason, R.-P.; Kadiiska, M.-B. Reinterpreting the best biomarker of oxidative stress: The 8-iso-prostaglandin F2α/prostaglandin F2α ratio shows complex origins of lipid peroxidation biomarkers in animal models. Free Radic. Biol. Med. 2016, 95, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Barocas, D.-A.; Motel Motley, S.; Cookson, M.-S.; Chang, S.-S.; Penson, D.; Dai, Q.; Milne, G.; Roberts, L.-J.; Morrow, J.; Concepcion, R.-S.; et al. Oxidative stress measured by urine F2-isoprostane level is associated with prostate cancer. J. Urol. 2011, 85, 2102–2107. [Google Scholar] [CrossRef] [PubMed]
- Weber, D.; Milkovic, L.; Bennett, S.-J.; Griffiths, H.-R.; Zarkovic, N.; Grune, T. Measurement of HNE-protein adducts in human plasma and serum by ELISA—Comparison of two primary antibodies. Redox Biol. 2013, 1, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Bennett, L.; Rojas, S.; Seefeldt, T. Role of antioxidants in the prevention of cancer. J. Exp. Clin. Med. 2012, 4, 215–222. [Google Scholar] [CrossRef]
- Kumar, S. The Importance of Antioxidant and their role in Pharmaceutical science. Asian J. Res. Chem. Pharmac. Sci. 2014, 1, 27–44. [Google Scholar]
- Rizzo, A.-M.; Berselli, P.; Zava, S.; Montorfano, G.; Negroni, M.; Corsetto, P.; Berra, B. Endogenous antioxidants and radical scavengers. Adv. Exp. Med. Biol. 2010, 698, 52–67. [Google Scholar] [PubMed]
- Takashima, M.; Horie, M.; Shichirini, M.; Hagihara, Y.; Yoshida, Y.; Niki, E. Assessment of antioxidant capacity for scavenging free radicals in vitro: A rational basis and practical application. Free Radic. Biol. Med. 2012, 52, 1242–1252. [Google Scholar] [CrossRef] [PubMed]
- Kiokias, S.; Varzakas, T.; Oreopoulou, V. In vitro activity of vitamins, flavonoids, and natural phenolic antioxidants against the oxidative deterioration of oil-based systems. Crit. Rev. Food Sci. Nutr. 2008, 48, 78–93. [Google Scholar] [CrossRef] [PubMed]
- Mamede, A.-C.; Tavares, S.-D.; Abrantes, A.-M.; Trindade, J.; Maia, J.-M.; Botelho, M.-F. The role of vitamins in cancer: A review. Nutr. Cancer 2011, 63, 479–494. [Google Scholar] [CrossRef] [PubMed]
- Roginsky, V.; Lissi, A.-E. Review of methods to determine chain-breaking antioxidant activity in food. Food Chem. 2005, 92, 235–254. [Google Scholar] [CrossRef]
- Puertollano, M.-A.; Puertollano, E.; de Cienfugos, G.-A.; de Pablo, M.-A. Dietary antioxidants: Immunity and host defense. Curr. Top. Med. Chem. 2011, 11, 1752–1766. [Google Scholar] [CrossRef] [PubMed]
- Traber, G.-M.; Jeffrey, A. Vitamin E, antioxidant and nothing more. Free Radic. Biol. Med. 2007, 43, 4–15. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-K.; Im, G.-J.; An, Y.-S.; Lee, S.-H.; Jung, H.-H.; Park, S.-Y. The effects of the antioxidant α-tocopherol succinate on cisplatin-induced ototoxicity in HEI-OC1 auditory cells. Int. J. Pediatr. Otorhinolaryngol. 2016, 86, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; De Camargo, C.-A. Tocopherols and tocotrienols in common and emerging dietary sources: Occurrence, applications, and health benefits. Int. J. Mol. Sci. 2016, 17, 1745. [Google Scholar] [CrossRef] [PubMed]
- Bjelakovic, G.; Nikolova, D.; Gluud, L.-L. Antioxidant supplements for prevention of mortality in healthy participants and patients with various diseases. JAMA Clin. Evid. Syn. 2013, 310, 178–179. [Google Scholar]
- Narra, M.-R.; Rajendar, K.; Rudra, R.; Rao, J.-V.; Begum, G. The role of vitamin C as antioxidant in protection of biochemical and haematological stress induced by chlorpyrifos in freshwater fish Clarias batrachus. Chemosphere 2015, 132, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Kiokias, S.; Proestos, C.; Varzakas, T.-A. Review of the structure, biosynthesis, absorption of carotenoids-analysis and properties of their common natural extract. Curr. Res. Nutr. Food Sci. 2016, 4, 25–37. [Google Scholar] [CrossRef]
- Meléndez-Martínez, A.-M.; Stinco, C.-M.; Brahm, P.-M.; Vicario, I.-M. Analysis of carotenoids and tocopherols in plant matrices and assessment of their in vitro antioxidant capacity. Methods Mol. Biol. 2014, 1153, 77–97. [Google Scholar] [PubMed]
- Pantavos, A.; Ruite, R.; Feskens, F.-E.; de Keyser, E.-C.; Hofman, A.; Stricker, H.-B.; Franco, O.-H.; Kiefte-de Jong, J.-C. Total dietary antioxidant capacity, individual antioxidant intake and breast cancer risk: The Rotterdam study. Int. J. Cancer 2015, 136, 2178–2186. [Google Scholar] [CrossRef] [PubMed]
- Beker, B.-Y.; Bakir, T.; Sonmezoglu, F.-I.; Apak, R. Antioxidant protective effect of flavonoids on linoleic acid peroxidation induced by copper (II)/ascorbic acid. Chem. Phys. Lipids 2011, 164, 732–739. [Google Scholar] [CrossRef] [PubMed]
- Mattia, C.-D.; Sacchetti, G.; Mastrocola, D.; Pittia, P. Effect of phenolic antioxidants on the dispersion state and chemical stability of olive oil ο/w emulsions. Food Res. Int. 2009, 42, 1163–1170. [Google Scholar] [CrossRef]
- Tsimogiannis, D.; Oreopoulou, V. Defining the role of flavonoid structure on cottonseed oil stabilization: Study of A- and C-ring substitution. J. Am. Oil Chem. Soc. 2007, 84, 129–136. [Google Scholar] [CrossRef]
- Kiokias, S.; Gordon, M. Properties of carotenoids in vitro and in vivo. Food Rev. Int. 2004, 20, 99–121. [Google Scholar] [CrossRef]
- Dashti, M.; Kulik, W.; Hoek, F.; Veerman, E.-C.; Peppelenbosch, M.-P.; Rezaee, F. A phospholipidomic analysis of all defined human plasma lipoproteins. Sci. Rep. 2011, 1, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Mohammad, P.-I. Trans fatty acids—A risk factor for cardiovascular disease. Pak. J. Med. Sci. 2014, 30, 194–197. [Google Scholar]
- Winklhofer-Rooba, B.; Faustmanna, G.; Roob, J.-M. Low-density lipoprotein oxidation biomarkers in human health and disease and effects of bioactive compounds. Free Radic. Biol. Med. 2017, 111, 38–86. [Google Scholar] [CrossRef] [PubMed]
- Cocate, P.-G.; Natali, A.-J.; Alfenas, R.-G.; de Oliveira, A. Carotenoid consumption is related to lower lipid oxidation and DNA damage in middle-aged men. Br. J. Nutr. 2015, 114, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Krauss, R.-M. Lipoprotein subfractions and cardiovascular disease risk. Curr. Opin. Lipidol. 2010, 21, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Amarowicz, R.; Pegg, R.-B. The potential protective effects of phenolic compounds against low-density lipoprotein oxidation. Curr. Pharm. Des. 2017, 23, 2754–2766. [Google Scholar] [CrossRef] [PubMed]
- Parthasaranthy, S.; Raghavamenon, A.; Raghavamenon, S.; Garelnabi, O.-M.; Santanam, N. Oxidized low-density lipoprotein. Methods Mol. Biol. 2010, 610, 403–417. [Google Scholar]
- Kiokias, S.; Gordon, M. Dietary supplementation with a natural carotenoid mixture decreases oxidative stress. Eur. J. Clin. Nutr. 2003, 57, 1135–1140. [Google Scholar] [CrossRef] [PubMed]
- Stocker, R.; Keaney, J.-F. Role of oxidative modifications in atherosclerosis. Physiol. Rev. 2004, 84, 1381–1478. [Google Scholar] [CrossRef] [PubMed]
- Kiokias, S. In vitro and in vivo antioxidant properties of natural carotenoid mixtures. Ph.D. Thesis, Faculty of Life Sciences, School of Food Biosciences, The University of Reading, Reading, UK, July 2002. [Google Scholar]
- Itabe, H. Oxidized low-density lipoprotein as a biomarker of in vivo oxidative stress: From atherosclerosis to periodontitis. J. Clin. Biochem. Nutr. 2012, 51, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ghaffari, A.-M.; Ghiasvand, T. Kinetic study of low density lipoprotein oxidation by copper. Indian J. Clin. Biochem. 2010, 25, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Pizzimenti, S.; Ciamporcero, E.; Daga, M.; Pettazoni, E.; Arcaro, A.; Cetrangolo, G.; Minelli, R.; Dianzani, C.; Lepore, A.; Gentile, F.; et al. Interaction of aldehydes derived from lipid peroxidation and membrane proteins. Front. Physiol. 2013, 4, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Mahfouz, M.-M.; Zhou, O.; Kummerow, F.-A. Effect of curcumin on LDL oxidation in vitro, and lipid peroxidation and antioxidant enzymes in cholesterol fed rabbits. Int. J. Vitam. Nutr. Res. 2011, 81, 378–391. [Google Scholar] [CrossRef] [PubMed]
- Fuhrman, B.; Volkova, N.; Rosenhalt, M.; Aviram, M. Lycopene synergistically inhibits LDL oxidation in combination with vitamin E, rosmarinic acid, carnosic acid, or garlic. Antioxid. Redox Signal. 2000, 2, 491–506. [Google Scholar] [CrossRef] [PubMed]
- Gammone, M.-A.; Riccioni, G.; D’Orazio, N. Marine carotenoids against oxidative stress: Effects on human health. Mar. Drugs 2015, 13, 6226–6246. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.-D.; Youn, Y.-K.; Shin, W.-G. Positive effects of astaxanthin on lipid profiles and oxidative stress in overweight subjects. Plant Foods Hum. Nutr. 2011, 66, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-W.; Hellsten, A.; Jacobsson, L.-S.; Blomqvist, H.-M.; Olsson, A.-G.; Yuan, X. Alpha-tocopherol and astaxanthin decrease macrophage infiltration, apoptosis and vulnerability in atheroma of hyperlipidaemic rabbits. J. Mol. Cell. Cardiol. 2004, 37, 969–978. [Google Scholar] [CrossRef] [PubMed]
- Ciccone, M.-M.; Cortese, M.-M.; Cortese, F.; Gesualdo, M.; Carbonara, S.; Zito, A.; Ricci, G. Dietary intake of carotenoids and their antioxidant and anti-inflammatory effects in cardiovascular care. Mediat. Inflamm. 2013, 1, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bub, A.; Waltz, B.; Abrahamse, Z.; Adam, S.; Wever, J.; Muller, H.-S.; Rechenmmer, G. Moderate intervention with carotenoid rich vegetable products reduces lipid peroxidation in men. Am. Soc. Nutr. Sci. 2000, 135, 2200–2206. [Google Scholar] [CrossRef] [PubMed]
- Upritchard, J.-E.; Sutherland, W.-H.; Mann, J.-I. Effect of supplementation with tomato juice, vitamin E, and vitamin C on LDL oxidation and products of inflammatory activity in type 2 diabetes. Diabetes Care 2000, 23, 733–738. [Google Scholar] [CrossRef] [PubMed]
- Kiokias, S.; Varzakas, T.; Arvanitoyannis, I.; Labropoulos, A. Lipid oxidation. In Advances in Food Biochemistry; Yildiz, F., Ed.; CRC Press: New York, NY, USA, 2009; pp. 384–403. [Google Scholar]
- Wen, Y. The effect of pharmacological supplementation with Vit-C on LDL oxidation. Br. J. Clin. Pharmacol. 1997, 44, 94–97. [Google Scholar] [CrossRef] [PubMed]
- Hillstrom, R.-J.; Yacapin-Ammons, A.-K.; Lynch, S.-M. Vitamin C inhibits lipid oxidation in human HDL. J. Nutr. 2003, 133, 3047–3051. [Google Scholar] [CrossRef] [PubMed]
- Shariat, S.-Z.; Mostafavi, S.-A.; Khakpour, F. Antioxidant effects of vitamins C and E on the low-density lipoprotein oxidation mediated by myeloperoxidase. Iran. Biomed. J. 2013, 17, 22–28. [Google Scholar]
- Jacobsson, L.-S.; Yuan, X.-M.; Zieden, B.; Olsson, A.-G. Effects of α-tocopherol and astaxanthin on LDL oxidation and atherosclerosis in WHHL rabbits. Atherosclerosis 2004, 173, 231–237. [Google Scholar] [PubMed]
- Parameswari, C.-S.; Vijayageetha, B.; Vijayakumar, R.; Parameswari, C.-S.; Vijayageetha, B.; Vijayakumar, R. Effect of supplementation of vitamin E, vitamin C and reduced glutathione on copper ion induced lipoprotein oxidation in renal diseased patients—An in vitro study. Indian J. Clin. Biochem. 2006, 21, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Carr, C.-A.; Zhu, B.-Z.; Frei, B. Potential Antiatherogenic Mechanisms of Ascorbate (Vitamin C) and α-Tocopherol (Vitamin E). Circ. Res. 2018, 87, 349–354. [Google Scholar] [CrossRef]
- Dotan, Y.; Lichtenberg, D.; Pinchuk, I. No evidence supports vitamin E indiscriminate supplementation. Biofactors 2009, 35, 469–473. [Google Scholar] [CrossRef] [PubMed]
- Niki, E. Assessment of antioxidant capacity in vitro and in vivo. Free Radic. Biol. Med. 2010, 49, 503–515. [Google Scholar] [CrossRef] [PubMed]
- Niki, E. Do free radicals play causal role in atherosclerosis? Low density lipoprotein oxidation and vitamin E. J. Clin. Biochem. Nutr. 2011, 48, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Naderi, G.-A.; Seddigheh, A.; Sarraf-Zadegan, N. Antioxidant effect of flavonoids on the susceptibility of LDL oxidation. Mol. Cell. Biochem. 2003, 246, 193–196. [Google Scholar] [CrossRef] [PubMed]
- Carmeli, E.; Fogelman, Y. Antioxidant effect of polyphenolic glabridin on LDL oxidation. Toxicol. Ind. Health 2009, 25, 321–324. [Google Scholar] [CrossRef] [PubMed]
- Lam, R.-Y.; Woo, A.-Y.; Leung, P.-S.; Cheng, C.-H. Antioxidant actions of phenolic compounds found in dietary plants on low-density lipoprotein and erythrocytes in vitro. J. Am. Coll. Nutr. 2007, 26, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Costa-Mugica, A.; Elsa Batista, A.; Diadelis Mondejar, G.; Soto-López, Y.; Brito-Navarro, V.; Maria Vázquez, A.; Brömme, D.; Zaldívar-Muñoz, C.; Vidal-Novoa, A.; de Oliveira, A.; et al. Inhibition of LDL-oxidation and antioxidant properties related to polyphenol content of hydrophilic fractions from seaweed Halimeda Incrassata. Braz. J. Pharm. Sci. 2012, 48. [Google Scholar] [CrossRef]
- Singh, N.; Singh, S.-K.; Bhargava, V. Status of LDL oxidation and antioxidant potential of LDL in type II diabetes. Biomed. Res. 2010, 21, 416–418. [Google Scholar]
- Aviram, M.; Dornfel, M.; Kaplan, M.; Coleman, R.; Gaitini, D.; Nitecki, S.; Hofman, A.; Rosenblat, M.; Volkova, N.; Presser, D.; et al. Pomegranate juice flavonoids inhibit low-density lipoprotein oxidation and cardiovascular diseases: Studies in atherosclerotic mice and in humans. Drugs Exp. Clin. Res. 2002, 28, 49–62. [Google Scholar] [PubMed]
- Carru, C.; Pasciu, V.; Sotgia, S.; Zinellu, A.; Nicoli, M.-C.; Deiana, L. The oxidative state of LDL is the major determinant of anti/prooxidant effect of coffee on Cu2+ catalysed peroxidation. Open Biochem. J. 2018, 12, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.-F.; Liu, R.-H. Novel low-density lipoprotein (LDL) oxidation model: Antioxidant capacity for the inhibition of LDL oxidation. J. Agric. Food Chem. 2004, 52, 6818–6823. [Google Scholar] [CrossRef] [PubMed]
- Kaur, V.; Kumar, M.; Kumar, A.; Kaur, S. Pharmacotherapeutic potential of phytochemicals: Implications in cancer chemoprevention and future perspectives. Biomed. Pharmacother. 2017, 97, 564–586. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Izakovic, M.; Mazur, M.; Rhodes, J.-C.; Telser, J. Oxidative damage and cancer incidence. Mol. Cell. Biochem. 2004, 266, 37–56. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Kaur, P.; Sandhir, R.; Kiran, R. Protective effects of vitamin E against atrazine-induced genotoxicity in rats. Mutat. Res. 2008, 654, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Friedberg, E.-C.; Aguilera, A.; Gellert, M.; Hanawalt, P.-C.; Hays, J.-B.; Lehmann, A.-R.; Lindahl, T.; Lowndes, N.; Sarasin, A.; Wood, R.-D. DNA repair: From molecular mechanism to human disease. DNA Repair. 2006, 13, 986–996. [Google Scholar] [CrossRef]
- Woodbine, L.; Brunton, H.; Goodarzi, A.-A.; Shibata, A.; Jeggo, P. Endogenously induced DNA double strand breaks arise in heterochromatic DNA regions and require ataxia telangiectasia mutated and Artemis for their repair. Nucleic Acids Res. 2011, 39, 6986–6997. [Google Scholar] [CrossRef] [PubMed]
- Shaposhnikov, S.; Thomsen, P.D.; Collins, A.R. Combining fluorescent in situ hybridization with the comet assay for targeted examination of DNA damage and repair. Methods Mol. Biol. 2011, 682, 115–132. [Google Scholar] [PubMed]
- Singh, S.-K.; Szulik, M.-W.; Ganguly, M.; Khutsishvili, M.; Stone, P.-M.; Marky, A.-L. Characterization of DNA with an 8-oxoguanine modification. Nucleic Acids Res. 2011, 39, 6789–6801. [Google Scholar] [CrossRef] [PubMed]
- Slyskova, J.; Langie, A.-S.; Collins, A.-R.; Vodicka, P. Functional evaluation of DNA repair in human biopsies and their relation to other cellular biomarkers. Front. Genet. 2014, 5, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Haghdoost, S.; Sjölander, L.; Czene, S.; Harms-Ringdahl, M. The nucleotide pool is a significant target for oxidative stress. Free Radic. Biol. Med. 2006, 41, 620–626. [Google Scholar] [CrossRef] [PubMed]
- Damas, J.; Carneiro, J.; Conalves, J.; Stewart, J.-B.; Samuels, D.-C.; Amorim, A. Mitochondrial DNA deletions are associated with non-B DNA conformations. Nucleic Acids Res. 2012, 40, 7606–7621. [Google Scholar] [CrossRef] [PubMed]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, C. 8-hydroxy-2′-deoxyguanosine (8-OHdG): A critical biomarker of oxidative stress and carcinogenesis. J. Environ. Sci. Health. Part C Environ. Carcinog. Ecotoxicol. Rev. 2009, 27, 120–139. [Google Scholar] [CrossRef] [PubMed]
- Toyokuni, S. Reactive oxygen species-induced molecular damage and its application in pathology. Pathol. Int. 1999, 49, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.-R.; Azqueta, A. DNA repair as a biomarker in human biomonitoring studies: Further applications of the comet assay. Mutat. Res. 2012, 736, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Nandhakumar, S.; Parasuraman, S.; Shanmugam, M.-M.; Ramachandra, R.; Parkash, C.; Vishnu, B. Evaluation of DNA damage using single-cell gel electrophoresis (Comet Assay). J. Pharmacol. Pharmacother. 2011, 2, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Ersson, C.; Möller, L. The effects on DNA migration by altering parameters in the comet assay protocol, including agarose density, electrophoresis conditions as well as durations of enzyme and alkaline treatments. Mutagenesis 2011, 26, 689–695. [Google Scholar] [CrossRef] [PubMed]
- Azqueta, A. Comet assay to measure DNA repair: Approach and applications. Front. Genet. 2014, 5, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Hofer, T.; Karlsson, H.-L.; Möller, L. DNA oxidative damage and strand breaks in young healthy individuals: A gender difference and the role of life style factors. Free Radic. Res. 2006, 40, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Astley, S.-B.; Hughes, D.-A.; Wright, A.-J.; Elliott, R.-M.; Southon, S. DNA damage and susceptibility to oxidative damage in lymphocytes: Effects of carotenoids in vitro and in vivo. Br. J. Nutr. 2004, 91, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Herrero-Barbudo, C.; Soldevilla, B.; Sacristán, B.-P.; Herrera, M.; Granado-Lorencio, F.; Domínguez, G. Modulation of DNA-induced damage and repair capacity in humans after dietary intervention with lutein-enriched fermented milk. PLoS ONE 2013, 11, 8. [Google Scholar] [CrossRef] [PubMed]
- Haegele, A.-D.; Gillete, C.; O’Neil, C.; Wolfe, P.; Heimendinger, J.; Sedlacek, S.; Thompson, H.-J. Plasma xanthophyll carotenoids correlate inversely with indices of oxidative DNA damage and lipid peroxidation. Cancer Epidemiol. Biomark. Prev. 2000, 9, 421–425. [Google Scholar]
- Barcelos, R.-M.; Grotto, D.; Serpeloni, J.-M.; Autures, L.-M.; Knasmuller, S.; Barbosa, F. Bixin and norbixin protect against DNA-damage and alterations of redox status induced by methylmercury exposure in vivo. Mol. Mutagen. 2012, 53, 535–541. [Google Scholar] [CrossRef] [PubMed]
- Noroozi, M.; Angerson, J.-W.; Lean, E.-J. Effects of flavonoids and vitamin C on oxidative DNA damage to human lymphocytes. Am. J. Clin. Nutr. 1998, 67, 1210–1218. [Google Scholar] [CrossRef] [PubMed]
- Kardivel, R.; Sundaram, K.; Mani, S.; Samuel, S.; Elango, N.; Panneerselvam, C. Supplementation of ascorbic acid and alpha-tocopherol prevents arsenic-induced protein oxidation and DNA damage induced by arsenic in rats. Hum. Exp. Toxicol. 2007, 26, 939–946. [Google Scholar]
- Kontek, R.; Kontek, B.; Grzegorczyk, K. Vitamin C modulates DNA damage induced by hydrogen peroxide in human colorectal adenocarcinoma cell lines (HT29) estimated by comet assay in vitro. Arch. Med. Sci. 2013, 9, 1006–1012. [Google Scholar] [CrossRef] [PubMed]
- Asgard, R. Effects of Antioxidants and Prooxidants on Oxidative Stress and DNA Damage Using the Comet Assay; Digital Comprehensive Summaries of Dissertations; Faculty of Pharmacy, University of Uppsala: Uppsala, Sweden, 2014; pp. 22–30. [Google Scholar]
- Pazdro, R.; Burgess, J.-R. Differential effects of α-tocopherol and N-acetyl-cysteine on advanced glycation end product-induced oxidative damage and neurite degeneration in SH-SY5Y cells. Biochim. Biophys. Acta 2012, 1822, 550–556. [Google Scholar] [CrossRef] [PubMed]
- Makpol, S.; Zainuddin, A.; Rahim, N.-A.; Yusof, Y.-A.; Ngah, W.-Z. Alpha-tocopherol modulates hydrogen peroxide-induced DNA damage and telomere shortening of human skin fibroblasts derived from differently aged individuals. Plant Med. 2010, 76, 869–875. [Google Scholar] [CrossRef] [PubMed]
- Fantappie, O.; Lodovici, M.; Fabrizio, P.; Marchettia, S.; Fabbroni, V.; Solazzo, M.; Lasagna, N.; Pantaleo, P.; Mazzanti, R. Vitamin E protects DNA from oxidative damage in human hepatocellular carcinoma cell lines. Free Radic. Res. 2004, 38, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Ragin, C.; Minor, A.; Farmer, P.; Garte, S.; Gonzales, C.; Kalina, I.; Pino, M.; Popov, T.; Palli, D.; Peluso, M.; et al. Pooled analysis of studies on DNA adducts and dietary vitamins. Mutat. Res. 2010, 705, 77–82. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, B.-F.; Veloso, C.-A.; Nogueira-Machado, J.-A.; Chaves, M.-M. High doses of in vitro beta-carotene, alpha-tocopherol and ascorbic acid induce oxidative stress and secretion of IL-6 in peripheral blood mononuclear cells from healthy donors. Curr. Aging Sci. 2012, 5, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Nderitu, A.-M.; Dykes, L.; Awika, M.-J.; Minnaar, A.; Duodu, G.-K. Phenolic composition and inhibitory effect against oxidative DNA damage of cooked cowpeas as affected by simulated in vitro gastrointestinal digestion. Food Chem. 2013, 141, 1763–1771. [Google Scholar] [CrossRef] [PubMed]
- Burkard, M.; Leischner, L.; Buschc, S.; Venturelli, J.-F. Dietary flavonoids and modulation of natural killer cells: Implications in malignant and viral diseases. J. Nutr. Biochem. 2017, 46, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Vazhappilly, C.-M.; Vasantha, H.-P. Apple flavonoids suppress carcinogen-induced DNA damage in normal human bronchial epithelial cells. Oxid. Med. Cell. Longev. 2017, 1, 1–12. [Google Scholar]
- Rusak, G.; Piantanida, I.; Masić, L.; Kapuralin, K.; Durgo, K.; Kopjar, N. Spectrophotometric analysis of flavonoid-DNA interactions and DNA damaging/protecting and cytotoxic potential of flavonoids in human peripheral blood lymphocytes. Chem. Biol. Interact. 2010, 188, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.-C.; Wang, Y.-H.; Liou, C.-C.; Lin, Y.-C.; Huang, W.-H.; Liu, Y.-C. Induction of oxidative DNA damage by flavonoids of propolis: Its mechanism and implication about antioxidant capacity. Chem. Res. Toxicol. 2012, 25, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Sevgi, K.; Tepe, B.; Sarikurkcu, C. Antioxidant and DNA damage protection potentials of selected phenolic acids. Food Chem. Toxicol. 2015, 77, 200–204. [Google Scholar] [CrossRef] [PubMed]
- Fabiani, R.; Rosignoli, P.; De Bartolomeno, A.; Fuccelii, R.; Servili, M.; Montedero, G.-F.; Morozzi, G. Oxidative DNA damage is prevented by extracts of olive oil, hydroxytyrosol, and other olive phenolic compounds in human blood mononuclear cells and HL60 cells. J. Nutr. 2008, 138, 1411–1416. [Google Scholar] [CrossRef] [PubMed]
- Lodovici, M.; Guglielmi, F.; Meoni, M.; Dolara, P. Effect of natural phenolic acids on DNA oxidation in vitro. Food Chem. Toxicol. 2001, 39, 1205–1210. [Google Scholar] [CrossRef]
- Cemeli, E.; Schmid, T.-E.; Anderson, D. Modulation by flavonoids of DNA damage induced by estrogen-like compounds. Environ. Mol. Mutagen. 2004, 44, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Min, K.; Ebeler, S.-E. Flavonoid effects on DNA oxidation at low concentrations relevant to physiological levels. Food Chem. Toxicol. 2008, 46, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Barcelos, G.-R.; Grotto, D.; Serpeloni, J.-M.; Angeli, J.-P.; Rocha, B.-A.; de Oliveira Souza, V.-C.; Vicentini, J.-T.; Emanuelli, T.; Bastos, J.-K.; Antunes, L.M.; et al. Protective properties of quercetin against DNA damage and oxidative stress induced by methylmercury in rats. Arch. Toxicol. 2011, 85, 1151–1157. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Somasagara, R.-R.; Hegde, M.; Nishana, M.; Tadi, S.-K.; Srivastava, M.; Choudhary, B.; Raghavan, S.-C. Quercetin, a natural flavonoid interacts with DNA, arrests cell cycle and causes tumor regression by activating mitochondrial pathway of apoptosis. Sci. Rep. Arch. 2016, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Rytter, E.; Johansson, C.; Vessby, B.; Sjödin, A.; Möller, L.; Åkesson, B.; Basu, S. Biomarkers of oxidative stress in overweight men are not influenced by a combination of antioxidants. Free Radic. Res. 2010, 44, 522–528. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Researchers/Study (per Chronological Order) | Experimental Conditions | Effect on LDL Oxidation |
|---|---|---|
| Kiokias and Gordon (2003) [44]/Clinical trial | Supplementation of 30 healthy volunteers with 30 mg active carotenoid/day (mixture of palm oil carotenes, lycopene, paprika, lutein, bixin) for 3 weeks. | Reduced ex vivo copper-induced LDL oxidation monitored by measurement of conjugated dienes at 233 nm. |
| Naderi et al. (2003) [68]/In vitro study | Model of LDL oxidation monitored by the change in 234-absorbance in presence of various flavonoids. | Genistein, morin and naringin exterted a stronger inhibitory activity than quercetin or apigenin. |
| Jacobson et al. (2004) [62]/Clinical trial | Supplementation of hyperlipedemic rabbits with 500 mg α-tocopherol/kg for 24 weeks. | Increased resistance to LDL oxidation observed in carotenoid treated rabbits (lag time of LDL oxidation in treatment group almost 2 times higher than in the placebo). |
| Lam et al. (2009) [69]/In vitro study | Model of lipid peroxidation in LDL induced by AAPH radical initiator. | Selected phenolic compounds from dietary sources (6-gingerol and rhapontin) were found to exhibit a strong inhibitory effect against LDL oxidation whereas barbaloin possessed weaker effects. |
| Carmeli and Fogelman (2009) [69]/Clinical trial | Supplementation of 10 healthy subjects for 6 months with a licorice-root extract rich in the isoflavone glabridin. | LDL oxidative stress was reduced by 20% in terms of TBARS in the treatment group compared to baseline. |
| Ghaffari and Ghiasvand (2010) [48]/In vitro study | Model of LDL oxidation induced by cupric ions. | α-tocopherol (in the range 0–100 μmol/L) reduced LDL oxidative deterioration. |
| Choi et al. (2011) [53]/Clinical trial | Supplementation of 27 overweight and obese adults with the carotenoid astaxanthin in a placebo-controlled study performed for 12 weeks. | The treatment group presented lower levels of LDL oxidative biomarkers compared to the placebo group. |
| Costa-Mugica et al. (2012) [71]/In vitro study | Model of heparin-precipitated LDL exposed to Cu2+ ions with AAPH as the free radical generator. | Lyophilized aqueous extracts and phenolic-rich fractions of seaweed (H. incrassata) significantly inhibited LDL oxidation. |
| Shariat et al. (2013) [61]/In vitro study | Model of LDL oxidation mediated by myeloperoxidase | Vitamin C inhibited LDL oxidation with a concentration dependent effect (50–200 mM). |
| Cocate et al. (2015) [40]/Clinical trial | Supplementation of 296 healthy middle-aged supplemented with a carotenoid mixture (β-cryptoxanthin, lycopene, lutein plus zeaxanthin, β-carotene and α-carotene). | The daily carotenoid intake was inversely associated (p < 0.05) with the plasma oxidised-LDL concentrations. |
| Researchers/Study (per chronological order) | Experimental Conditions | Effect on LDL Oxidation |
|---|---|---|
| Kiokias and Gordon (2003) [44]/Clinical trial | Supplementation of 30 healthy volunteers with 30 mg active carotenoid/day (mixture of α,β-carotene, lycopene, paprika, lutein, bixin) for 3 weeks. | Carotenoids caused a significant reduction (p < 0.05) of in vivo DNA oxidative damage in terms of 8-OH-dG as biomarker. |
| Astley et al. (2004) [93]/Clinical trial | Supplementation of healthy males with 15 mg/d lutein, β-carotene or lycopene (natural isolate capsules) for 4 weeks (3 independent clinical trials). | Carotenoids presented an antioxidant Character protection by scavenging DNA-damaging free radicals modulation of DNA repair. |
| Fantappiè et al. (2004) [103] /In vitro study | Model of oxidative DNA damage in the human hepatocellular carcinoma. | Vitamin E protected DNA from oxidative damage as evidenced by the concentration of TBARS and 8-OH-dG biomarkers after carotenoid treatment. |
| Fabiani et al. (2008) [112]/In vitro study | Model of oxidative DNA damage in human blood mononuclear cells and HL60 cells. | Extracts of olive oil, hydroxytyrosol, and other olive phenolic compounds exerted a strong inhibitory effect against DNA damage. |
| Rusac et al. (2010) [109]/In vitro study | Model of flavonoid-DNA interactions in human peripheral blood lymphocytes. | Certain flavonoids (luteolin, apigenin and kaempferol) were shown effective in protecting DNA from oxidative damage induced by hydrogen peroxide. |
| Barcelos et al. (2012) [96]/Clinical trial | Rats were treated orally with quercetin (0.5–50 mg/kg/bw/day), over 45 days. | Quercetin concentrations (5.0 and 50.0 mg/kg/bw/day) were found to protect against DNA damage. |
| Herrero-Barbudo et al. (2013) [94] /Clinical trial | Supplementation of 10 humans with lutein-enriched fermented milk (containing lutein and lutein esters at concentration 4–8 mg free lutein/100 mL). | A significant increase in serum lutein levels, was associated with an improved resistance to DNA damage. |
| Cocate et al. (2014) [40]/Clinical trial | Supplementation of 296 healthy middle-aged supplemented with a with carotenoid mixture (β-cryptoxanthin, lycopene, lutein plus zeaxanthin, β-carotene and α-carotene). | The daily carotenoid intake was inversely associated with the production of urinary 8-OH-dG as oxidative stress biomarker (p < 0.05). |
| Asgard (2014) [100]/Clinical trial | 47 type-2 diabetes subjects supplemented for 12 weeks with 16 capsules/day (mixture of β-carotene + α-tocopherol). | Dietary supplementation did not affect the levels of biomarkers of oxidative stress and inflammation, despite substantially increased plasma concentrations of antioxidants. |
| Sevgi et al. (2015) [111]/In vitro study | Model of plasmid DNA oxidative damage in the presence of hydrogen peroxide and ultraviolet (UV) light. | Tested phenolic acids (ferulic, caffeic, rosmarinic, and vanillic acids) inhibited DNA damage. |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiokias, S.; Proestos, C.; Oreopoulou, V. Effect of Natural Food Antioxidants against LDL and DNA Oxidative Changes. Antioxidants 2018, 7, 133. https://doi.org/10.3390/antiox7100133
Kiokias S, Proestos C, Oreopoulou V. Effect of Natural Food Antioxidants against LDL and DNA Oxidative Changes. Antioxidants. 2018; 7(10):133. https://doi.org/10.3390/antiox7100133
Chicago/Turabian StyleKiokias, Sotirios, Charalampos Proestos, and Vassilki Oreopoulou. 2018. "Effect of Natural Food Antioxidants against LDL and DNA Oxidative Changes" Antioxidants 7, no. 10: 133. https://doi.org/10.3390/antiox7100133
APA StyleKiokias, S., Proestos, C., & Oreopoulou, V. (2018). Effect of Natural Food Antioxidants against LDL and DNA Oxidative Changes. Antioxidants, 7(10), 133. https://doi.org/10.3390/antiox7100133