

The DJ-1-Binding Compound Exerts a Protective Effect in Both In Vitro and In Vivo Models of Sepsis-Induced Acute Kidney Injury

 ,
,  , , ,
, , ,
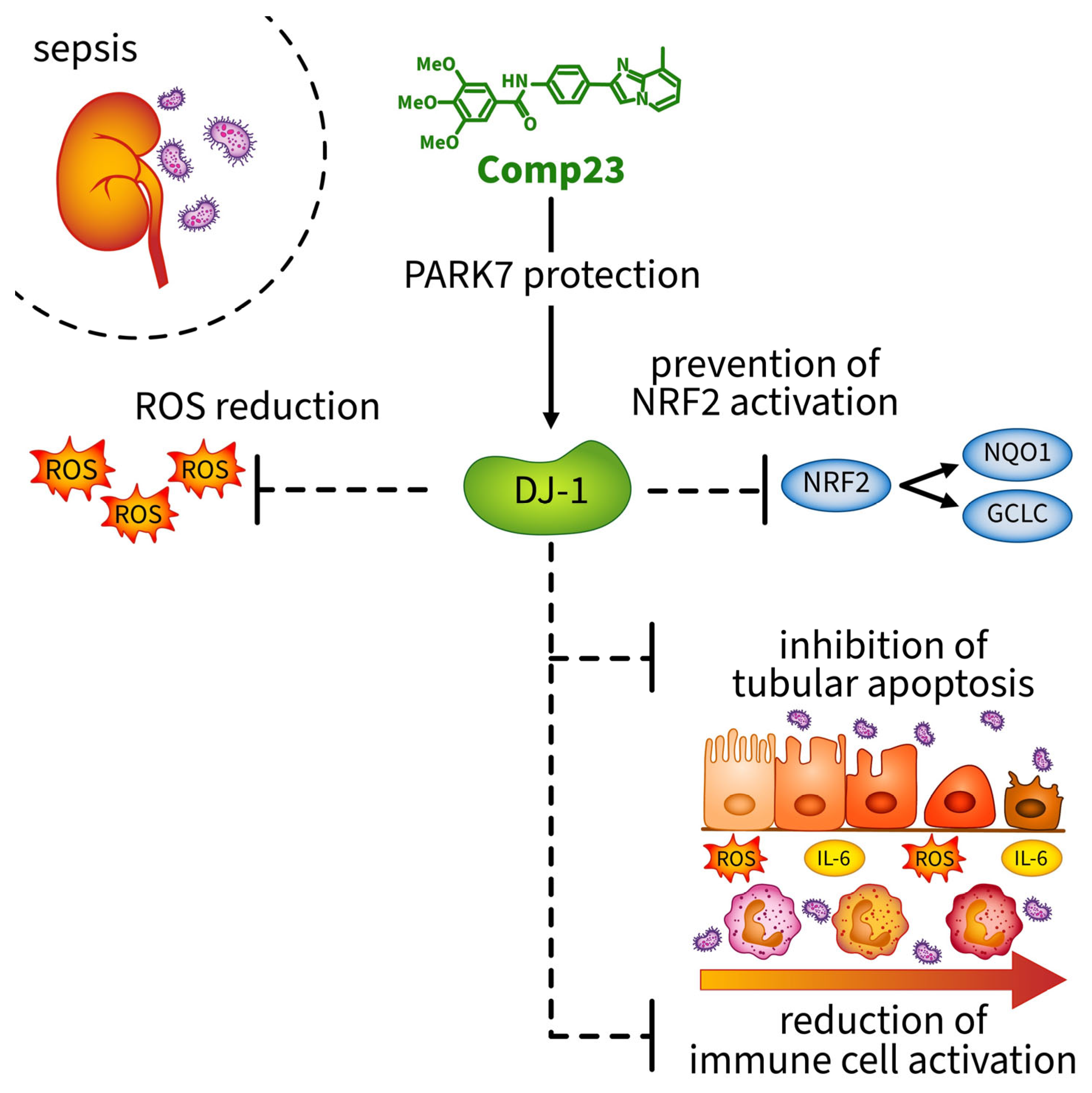
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Human Peripheral Blood Mononuclear Cells
2.3. Animal Model
2.4. Immunofluorescent Staining
2.5. Protein Isolation and Western Blot
2.6. RNA Isolation and cDNA Synthesis
2.7. Real-Time Polymerase Chain Reaction (PCR)
2.8. LDH Cytotoxicity Assay
2.9. MTT Cell Viability Assay
2.10. DCFDA Assay
2.11. Measurement of Renal Parameters
2.12. Statistical Analysis
3. Results
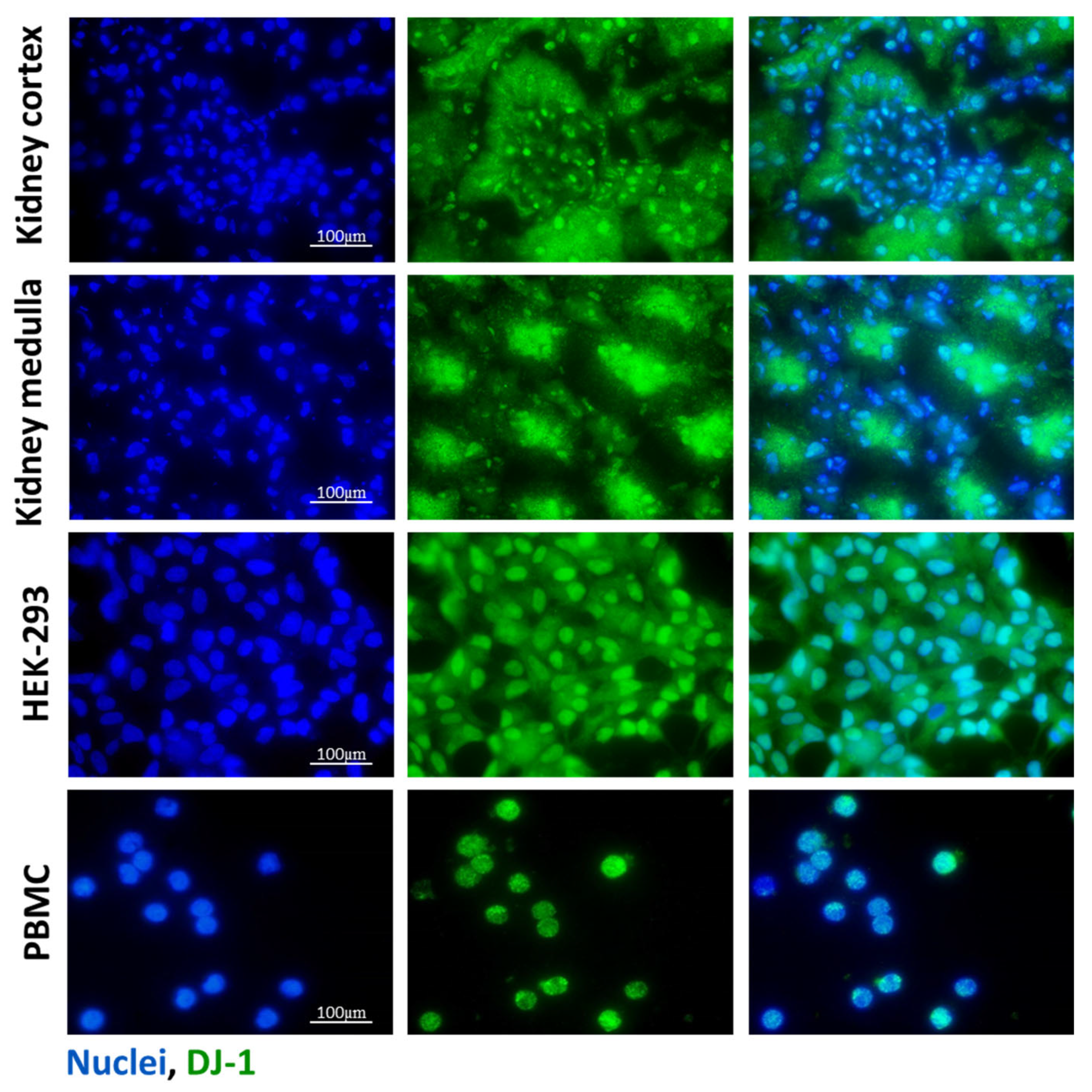
3.1. DJ-1 Expression in Kidney, HEK-293 Cells and PBMCs
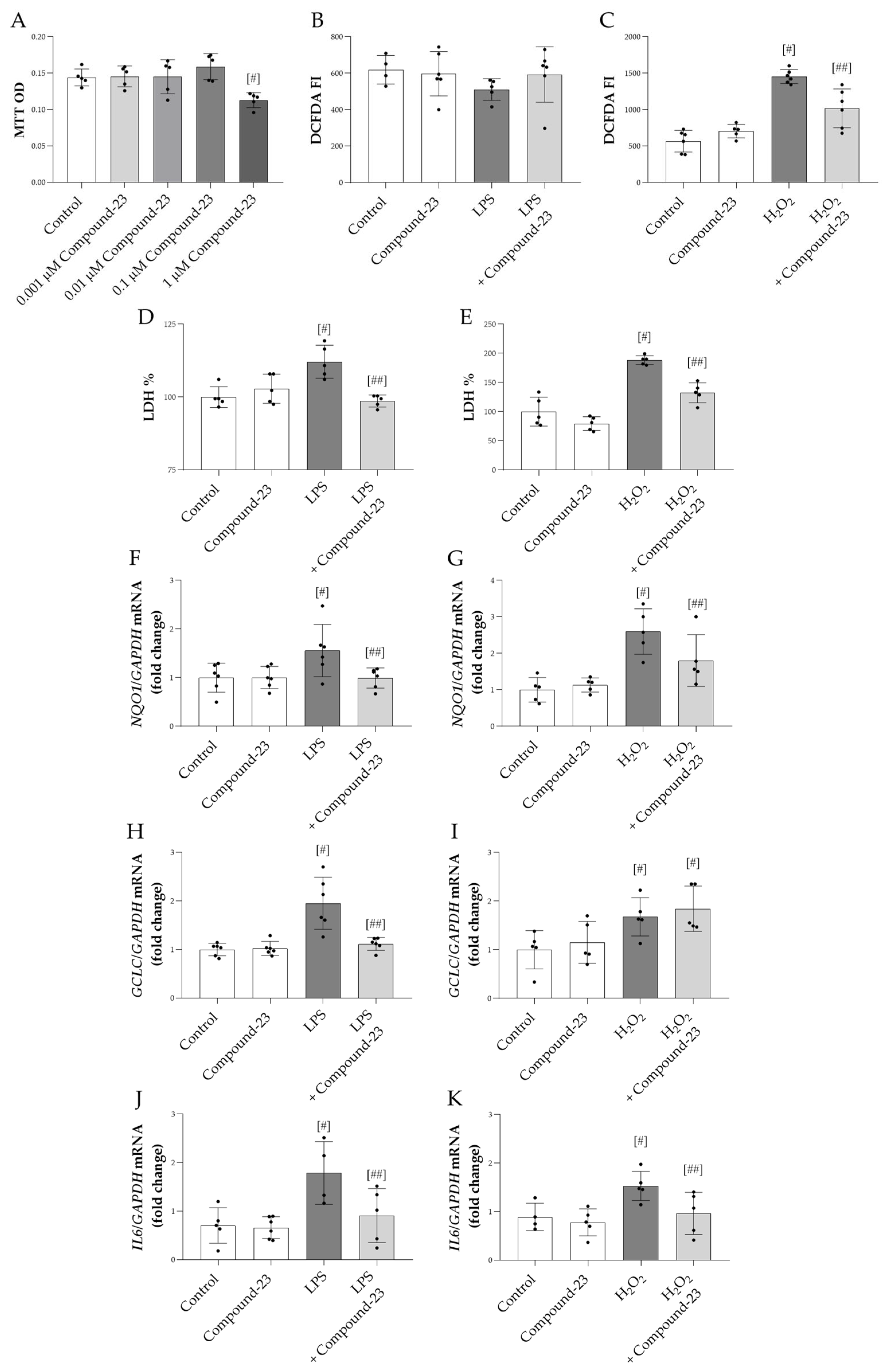
3.2. The Effect of DJ-1-Binding Compound-23 on the Oxidative Stress Response of Renal Cells
3.3. The Effect of DJ-1-Binding Compound-23 on the Proinflammatory Cytokine Production of PBMCs
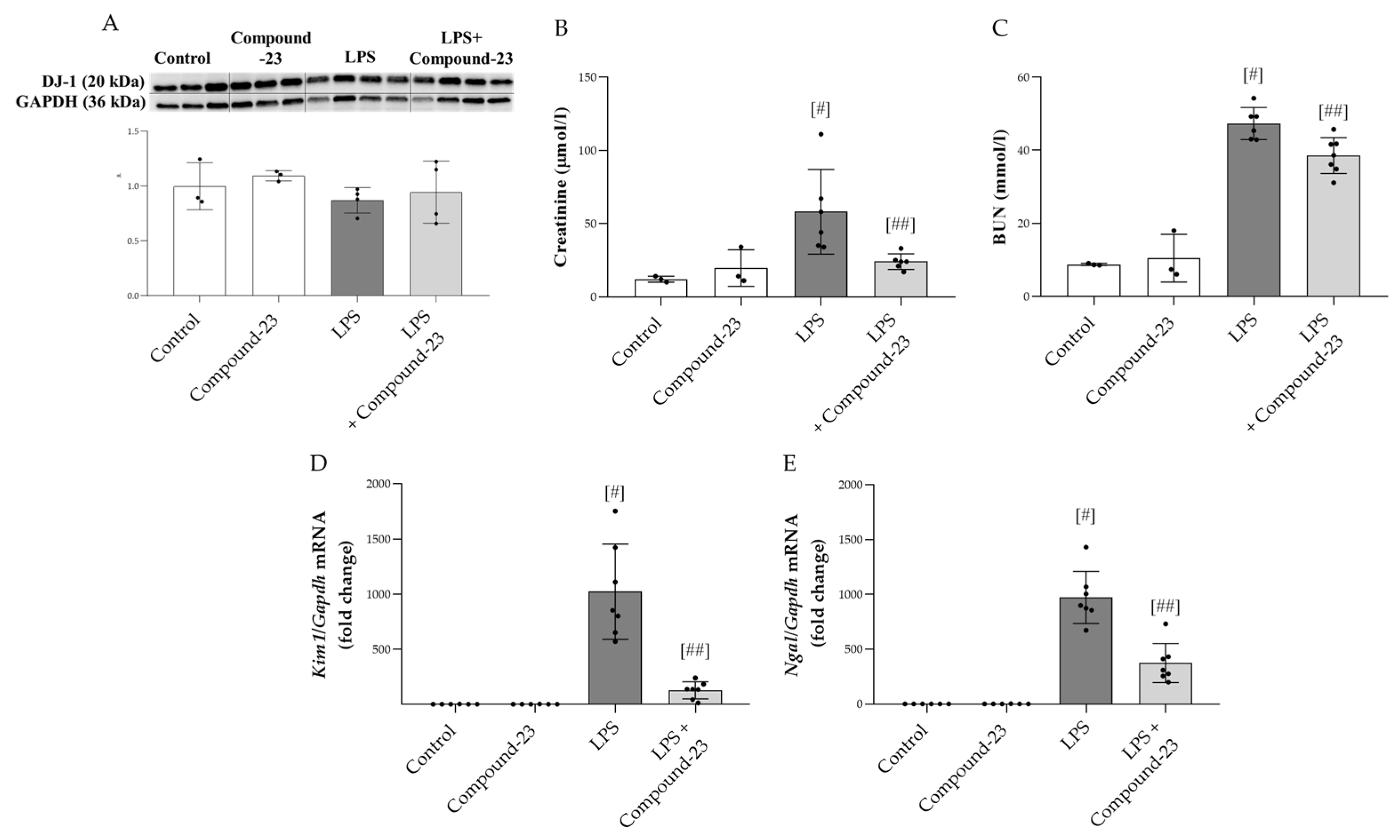
3.4. The Effect of DJ-1-Binding Compound-23 on LSP-Induced Acute Kidney Injury (AKI)
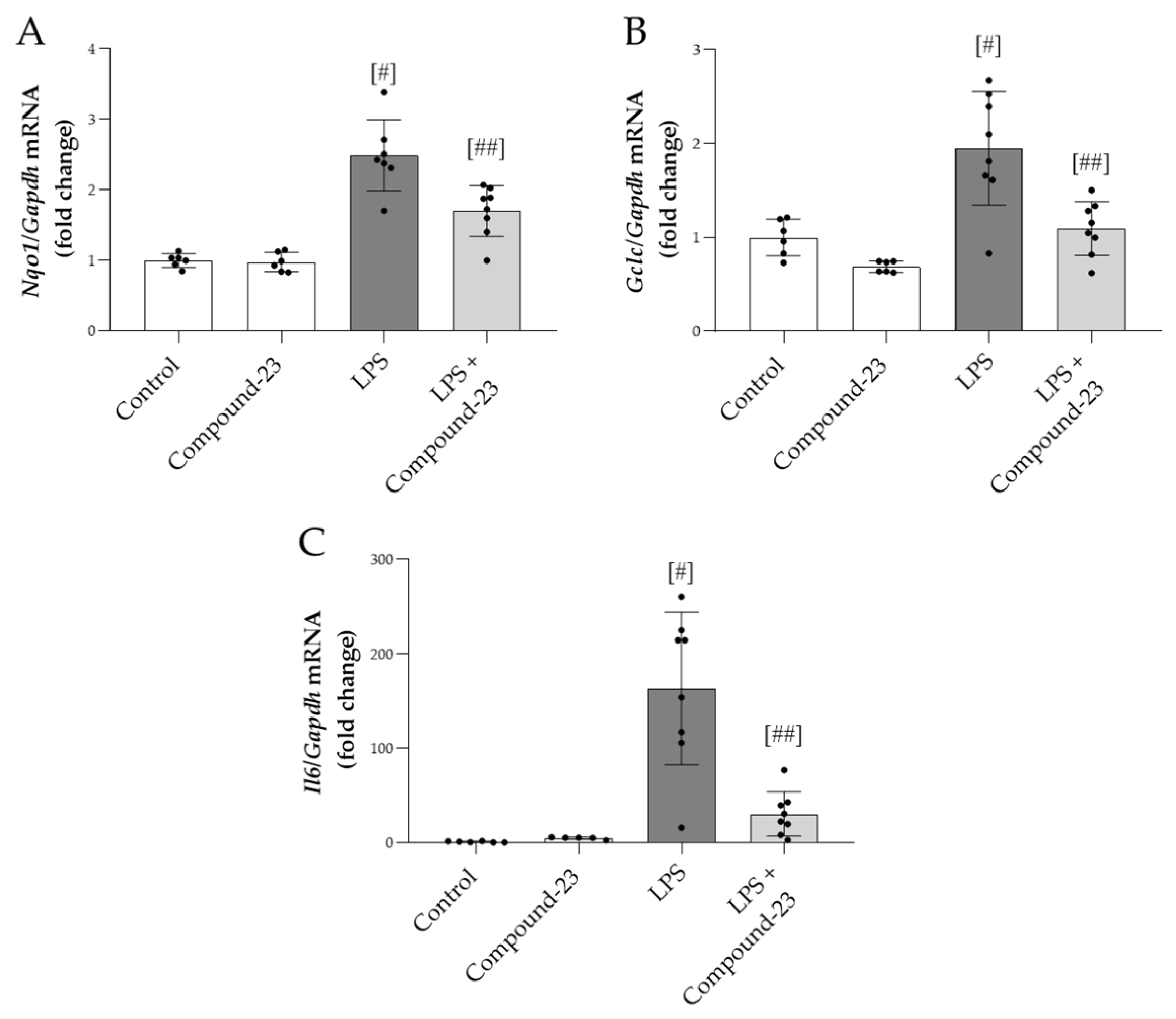
3.5. The Effect of DJ-1-Binding Compound-23 on the Renal Expression of Antioxidant and Proinflammatory Genes in the Mice Model of LPS-Induced Acute Kidney Injury (AKI)
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Peerapornratana, S.; Manrique-Caballero, C.L.; Gómez, H.; Kellum, J.A. Acute kidney injury from sepsis: Current concepts, epidemiology, pathophysiology, prevention and treatment. Kidney Int. 2019, 96, 1083–1099. [Google Scholar] [CrossRef] [PubMed]
- Coca, S.G.; Yusuf, B.; Shlipak, M.G.; Garg, A.X.; Parikh, C.R. Long-term risk of mortality and other adverse outcomes after acute kidney injury: A systematic review and meta-analysis. Am. J. Kidney Dis. 2009, 53, 961–973. [Google Scholar] [CrossRef] [PubMed]
- He, F.F.; Wang, Y.M.; Chen, Y.Y.; Huang, W.; Li, Z.Q.; Zhang, C. Sepsis-induced AKI: From pathogenesis to therapeutic approaches. Front. Pharmacol. 2022, 13, 981578. [Google Scholar] [CrossRef]
- Pap, D.; Veres-Székely, A.; Szebeni, B.; Vannay, Á. PARK7/DJ-1 as a Therapeutic Target in Gut-Brain Axis Diseases. Int. J. Mol. Sci. 2022, 23, 6626. [Google Scholar] [CrossRef]
- Cao, J.; Chen, X.; Ying, M.; He, Q.; Yang, B. DJ-1 as a Therapeutic Target Against Cancer. Adv. Exp. Med. Biol. 2017, 1037, 203–222. [Google Scholar] [CrossRef]
- Shen, Z.-Y.; Sun, Q.; Xia, Z.-Y.; Meng, Q.-T.; Lei, S.-Q.; Zhao, B.; Tang, L.-H.; Xue, R.; Chen, R. Overexpression of DJ-1 reduces oxidative stress and attenuates hypoxia/reoxygenation injury in NRK-52E cells exposed to high glucose. Int. J. Mol. Med. 2016, 38, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhou, J.; Li, Y.; Yang, F.; Lian, X.; Liu, W. Overexpression of DJ-1 alleviates autosomal dominant polycystic kidney disease by regulating cell proliferation, apoptosis, and mitochondrial metabolism in vitro and in vivo. Ann. Transl. Med. 2020, 8, 1175. [Google Scholar] [CrossRef]
- De Miguel, C.; Kraus, A.C.; Saludes, M.A.; Konkalmatt, P.; Ruiz Domínguez, A.; Asico, L.D.; Latham, P.S.; Offen, D.; Jose, P.A.; Cuevas, S. ND-13, a DJ-1-Derived Peptide, Attenuates the Renal Expression of Fibrotic and Inflammatory Markers Associated with Unilateral Ureter Obstruction. Int. J. Mol. Sci. 2020, 21, 7048. [Google Scholar] [CrossRef]
- Raninga, P.V.; Di Trapani, G.; Tonissen, K.F. The Multifaceted Roles of DJ-1 as an Antioxidant. Adv. Exp. Med. Biol. 2017, 1037, 67–87. [Google Scholar] [CrossRef]
- Shendelman, S.; Jonason, A.; Martinat, C.; Leete, T.; Abeliovich, A. DJ-1 is a redox-dependent molecular chaperone that inhibits alpha-synuclein aggregate formation. PLoS Biol. 2004, 2, e362. [Google Scholar] [CrossRef]
- Lippai, R.; Veres-Székely, A.; Sziksz, E.; Iwakura, Y.; Pap, D.; Rokonay, R.; Szebeni, B.; Lotz, G.; Béres, N.J.; Cseh, Á.; et al. Immunomodulatory role of Parkinson’s disease 7 in inflammatory bowel disease. Sci. Rep. 2021, 11, 14582. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Ishimori, C.; Takahashi-Niki, K.; Taira, T.; Kim, Y.C.; Maita, H.; Maita, C.; Ariga, H.; Iguchi-Ariga, S.M. DJ-1 binds to mitochondrial complex I and maintains its activity. Biochem. Biophys. Res. Commun. 2009, 390, 667–672. [Google Scholar] [CrossRef] [PubMed]
- Clements, C.M.; McNally, R.S.; Conti, B.J.; Mak, T.W.; Ting, J.P. DJ-1, a cancer- and Parkinson’s disease-associated protein, stabilizes the antioxidant transcriptional master regulator Nrf2. Proc. Natl. Acad. Sci. USA 2006, 103, 15091–15096. [Google Scholar] [CrossRef]
- Dolgacheva, L.P.; Berezhnov, A.V.; Fedotova, E.I.; Zinchenko, V.P.; Abramov, A.Y. Role of DJ-1 in the mechanism of pathogenesis of Parkinson’s disease. J. Bioenerg. Biomembr. 2019, 51, 175–188. [Google Scholar] [CrossRef]
- He, F.; Ru, X.; Wen, T. NRF2, a Transcription Factor for Stress Response and Beyond. Int. J. Mol. Sci. 2020, 21, 4777. [Google Scholar] [CrossRef] [PubMed]
- Kiss, R.; Zhu, M.; Jójárt, B.; Czajlik, A.; Solti, K.; Fórizs, B.; Nagy, É.; Zsila, F.; Beke-Somfai, T.; Tóth, G. Structural features of human DJ-1 in distinct Cys106 oxidative states and their relevance to its loss of function in disease. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 2619–2629. [Google Scholar] [CrossRef]
- Wilson, M.A. The role of cysteine oxidation in DJ-1 function and dysfunction. Antioxid. Redox Signal 2011, 15, 111–122. [Google Scholar] [CrossRef]
- Ito, G.; Ariga, H.; Nakagawa, Y.; Iwatsubo, T. Roles of distinct cysteine residues in S-nitrosylation and dimerization of DJ-1. Biochem. Biophys. Res. Commun. 2006, 339, 667–672. [Google Scholar] [CrossRef]
- Kitamura, Y.; Watanabe, S.; Taguchi, M.; Takagi, K.; Kawata, T.; Takahashi-Niki, K.; Yasui, H.; Maita, H.; Iguchi-Ariga, S.M.; Ariga, H. Neuroprotective effect of a new DJ-1-binding compound against neurodegeneration in Parkinson’s disease and stroke model rats. Mol. Neurodegener. 2011, 6, 48. [Google Scholar] [CrossRef]
- Takahashi-Niki, K.; Inafune, A.; Michitani, N.; Hatakeyama, Y.; Suzuki, K.; Sasaki, M.; Kitamura, Y.; Niki, T.; Iguchi-Ariga, S.M.; Ariga, H. DJ-1-dependent protective activity of DJ-1-binding compound no. 23 against neuronal cell death in MPTP-treated mouse model of Parkinson’s disease. J. Pharmacol. Sci. 2015, 127, 305–310. [Google Scholar] [CrossRef]
- Veres-Székely, A.; Bernáth, M.; Pap, D.; Rokonay, R.; Szebeni, B.; Takács, I.M.; Lippai, R.; Cseh, Á.; Szabó, A.J.; Vannay, Á. PARK7 Diminishes Oxidative Stress-Induced Mucosal Damage in Celiac Disease. Oxid. Med. Cell Longev. 2020, 2020, 4787202. [Google Scholar] [CrossRef]
- Amatullah, H.; Maron-Gutierrez, T.; Shan, Y.; Gupta, S.; Tsoporis, J.N.; Varkouhi, A.K.; Teixeira Monteiro, A.P.; He, X.; Yin, J.; Marshall, J.C.; et al. Protective function of DJ-1/PARK7 in lipopolysaccharide and ventilator-induced acute lung injury. Redox Biol. 2021, 38, 101796. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, A.; Theilig, F.; Schweda, F.; Höcherl, K. Acute endotoxemia in mice induces downregulation of megalin and cubilin in the kidney. Kidney Int. 2012, 82, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Niu, Y.; Zhang, X.; Zhang, Y.; Yu, Y.; Huang, J.; Li, J.; Yu, C. Recombinant α-Klotho Protein Alleviated Acute Cardiorenal Injury in a Mouse Model of Lipopolysaccharide-Induced Septic Cardiorenal Syndrome Type 5. Anal Cell Pathol. 2019, 2019, 5853426. [Google Scholar] [CrossRef]
- Miao, S.; Lv, C.; Liu, Y.; Zhao, J.; Li, T.; Wang, C.; Xu, Y.; Wang, X.; Xiao, X.; Zhang, H. Pharmacologic Blockade of 15-PGDH Protects Against Acute Renal Injury Induced by LPS in Mice. Front. Physiol. 2020, 11, 138. [Google Scholar] [CrossRef] [PubMed]
- Mir, S.M.; Ravuri, H.G.; Pradhan, R.K.; Narra, S.; Kumar, J.M.; Kuncha, M.; Kanjilal, S.; Sistla, R. Ferulic acid protects lipopolysaccharide-induced acute kidney injury by suppressing inflammatory events and upregulating antioxidant defenses in Balb/c mice. Biomed. Pharmacother. 2018, 100, 304–315. [Google Scholar] [CrossRef]
- Zhang, S.; Li, R.; Dong, W.; Yang, H.; Zhang, L.; Chen, Y.; Wang, W.; Li, C.; Wu, Y.; Ye, Z.; et al. RIPK3 mediates renal tubular epithelial cell apoptosis in endotoxin-induced acute kidney injury. Mol. Med. Rep. 2019, 20, 1613–1620. [Google Scholar] [CrossRef]
- Inada, M.; Izawa, G.; Kobayashi, W.; Ozawa, M. 293 cells express both epithelial as well as mesenchymal cell adhesion molecules. Int. J. Mol. Med. 2016, 37, 1521–1527. [Google Scholar] [CrossRef]
- Gill, R.; Tsung, A.; Billiar, T. Linking oxidative stress to inflammation: Toll-like receptors. Free Radic. Biol. Med. 2010, 48, 1121–1132. [Google Scholar] [CrossRef]
- Liu, J.; Li, G.; Xie, W.J.; Wang, L.; Zhang, R.; Huang, K.S.; Zhou, Q.S.; Chen, D.C. Lipopolysaccharide Stimulates Surfactant Protein-A in Human Renal Epithelial HK-2 Cells through Upregulating Toll-like Receptor 4 Dependent MEK1/2-ERK1/2-NF-κB Pathway. Chin. Med. J. 2017, 130, 1236–1243. [Google Scholar] [CrossRef]
- Cuevas, S.; Zhang, Y.; Yang, Y.; Escano, C.; Asico, L.; Jones, J.E.; Armando, I.; Jose, P.A. Role of renal DJ-1 in the pathogenesis of hypertension associated with increased reactive oxygen species production. Hypertension 2012, 59, 446–452. [Google Scholar] [CrossRef]
- Lu, S.C. Glutathione synthesis. Biochim. Biophys. Acta 2013, 1830, 3143–3153. [Google Scholar] [CrossRef] [PubMed]
- Ross, D.; Siegel, D. The diverse functionality of NQO1 and its roles in redox control. Redox Biol. 2021, 41, 101950. [Google Scholar] [CrossRef] [PubMed]
- Krejsa, C.M.; Franklin, C.C.; White, C.C.; Ledbetter, J.A.; Schieven, G.L.; Kavanagh, T.J. Rapid activation of glutamate cysteine ligase following oxidative stress. J. Biol. Chem. 2010, 285, 16116–16124. [Google Scholar] [CrossRef]
- Zhang, J.; Xu, M.; Zhou, W.; Li, D.; Zhang, H.; Chen, Y.; Ning, L.; Zhang, Y.; Li, S.; Yu, M.; et al. Deficiency in the anti-apoptotic protein DJ-1 promotes intestinal epithelial cell apoptosis and aggravates inflammatory bowel disease via p53. J. Biol. Chem. 2020, 295, 4237–4251. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Chen, C.; Yang, D.; Ding, J.; Wang, G.; Ren, H. DJ-1 inhibits microglial activation and protects dopaminergic neurons in vitro and in vivo through interacting with microglial p65. Cell Death Dis. 2021, 12, 715. [Google Scholar] [CrossRef]
- Morgan, M.J.; Liu, Z.-g. Crosstalk of reactive oxygen species and NF-κB signaling. Cell Res. 2011, 21, 103–115. [Google Scholar] [CrossRef]
- Vaidya, V.S.; Ferguson, M.A.; Bonventre, J.V. Biomarkers of acute kidney injury. Annu. Rev. Pharmacol. Toxicol. 2008, 48, 463–493. [Google Scholar] [CrossRef]
- Kellum, J.A.; Romagnani, P.; Ashuntantang, G.; Ronco, C.; Zarbock, A.; Anders, H.-J. Acute kidney injury. Nat. Rev. Dis. Primers 2021, 7, 52. [Google Scholar] [CrossRef]
- Song, J.; Yu, J.; Prayogo, G.W.; Cao, W.; Wu, Y.; Jia, Z.; Zhang, A. Understanding kidney injury molecule 1: A novel immune factor in kidney pathophysiology. Am. J. Transl. Res. 2019, 11, 1219–1229. [Google Scholar] [PubMed]
- Kokkoris, S.; Pipili, C.; Grapsa, E.; Kyprianou, T.; Nanas, S. Novel biomarkers of acute kidney injury in the general adult ICU: A review. Ren. Fail. 2013, 35, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Pap, D.; Szebeni, B.H.; Vannay, Á.; Veres-Székely, A. Benzamide Derivatives as Anti-Inflammatory Compounds and Uses Thereof. WO2021240187A1, 2 December 2021. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism | Gene | Primer Pairs |
|---|---|---|
| mouse | Gapdh | F: 5′-ATC TGA CGT GCC GCCTGGAGAAAC-3′ |
| R: 5′-CCCGGCATCGAAGGTGGAAGAGT-3′ | ||
| mouse | Kim1 | F: 5′-TCC AGG GAA GCC GCA GAA AAA-3′ |
| R: 5′-GGA AGG CAA CCA CGC TTA GAG ATG-3′ | ||
| mouse | Ngal | F: 5′-GCC AGG CCC AGG ACT CAA CTC A-3′ |
| R: 5′-GTA CCACCT GCC CCG GAA CTGBAT-3′ | ||
| mouse | Nqo1 | F: 5′-TGG CCG AAC ACA AGA AGC-3′ |
| R: 5′-TGA ATC GGC CAG AGA ATG AC-3′ | ||
| mouse | Gclc | F: 5′-GGA CTT TGA TGC GCC TCC TTC CTC TG-3′ |
| R: 5′-AAA CCC CAA CCA TCC GAC CCT CTG-3′ | ||
| mouse | Il6 | F: 5′-AAC CAC GGC CTT CCC TAC TTC A-3′ |
| R: 5′-TGC CAT TGC ACA ACT CTT TTC TCA-3′ | ||
| human | GAPDH | F: 5′-AGC AAT GCC TCC TGC ACC ACC AA-3′ |
| R: 5′-GCG GCC ATC ACG CCA CAG TTT-3′ | ||
| human | NQO1 | F: 5′-CTG CTG CAG CGG CTT TGA AGA-3′ |
| R: 5′-GCC AGA ACA GAC TCG GCA GGA TAC-3′ | ||
| human | GCLC | F: 5′-AAA AGT CCG GTT GGT CCT GTC TGG-3′ |
| R: 5′-GGC TGT CCT GGT GTC CCT TCA ATC-3′ | ||
| human | IL6 | F: 5′-AAA GAT GGC TGA AAA AGA TGG AT-3′ |
| R: 5′-CTC TGG CTT GTT CCT CAC TAC TCT-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zrufkó, R.; Pajtók, C.; Szebeni, B.; Veres-Székely, A.; Bernáth, M.; Szász, C.; Bokrossy, P.; Szabó, A.J.; Vannay, Á.; Pap, D. The DJ-1-Binding Compound Exerts a Protective Effect in Both In Vitro and In Vivo Models of Sepsis-Induced Acute Kidney Injury. Antioxidants 2025, 14, 719. https://doi.org/10.3390/antiox14060719
Zrufkó R, Pajtók C, Szebeni B, Veres-Székely A, Bernáth M, Szász C, Bokrossy P, Szabó AJ, Vannay Á, Pap D. The DJ-1-Binding Compound Exerts a Protective Effect in Both In Vitro and In Vivo Models of Sepsis-Induced Acute Kidney Injury. Antioxidants. 2025; 14(6):719. https://doi.org/10.3390/antiox14060719
Chicago/Turabian StyleZrufkó, Réka, Csenge Pajtók, Beáta Szebeni, Apor Veres-Székely, Mária Bernáth, Csenge Szász, Péter Bokrossy, Attila J. Szabó, Ádám Vannay, and Domonkos Pap. 2025. "The DJ-1-Binding Compound Exerts a Protective Effect in Both In Vitro and In Vivo Models of Sepsis-Induced Acute Kidney Injury" Antioxidants 14, no. 6: 719. https://doi.org/10.3390/antiox14060719
APA StyleZrufkó, R., Pajtók, C., Szebeni, B., Veres-Székely, A., Bernáth, M., Szász, C., Bokrossy, P., Szabó, A. J., Vannay, Á., & Pap, D. (2025). The DJ-1-Binding Compound Exerts a Protective Effect in Both In Vitro and In Vivo Models of Sepsis-Induced Acute Kidney Injury. Antioxidants, 14(6), 719. https://doi.org/10.3390/antiox14060719