
Arginine-Mediated Liver Immune Regulation and Antioxidant Defense in Largemouth Bass (Micropterus salmoides): Multi-Omics Insights into Metabolic Remodeling During Nocardia seriolae Infection


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish Farming
2.2. Infection Experiment and Sample Collection
2.3. Histological Analysis
2.4. RNA Extraction, Reverse Transcription, and Quantitative PCR (qPCR)
2.5. Transcriptome Analysis
2.6. Untargeted Metabolome Analysis
2.7. Joint Analysis of Transcriptome and Metabolome
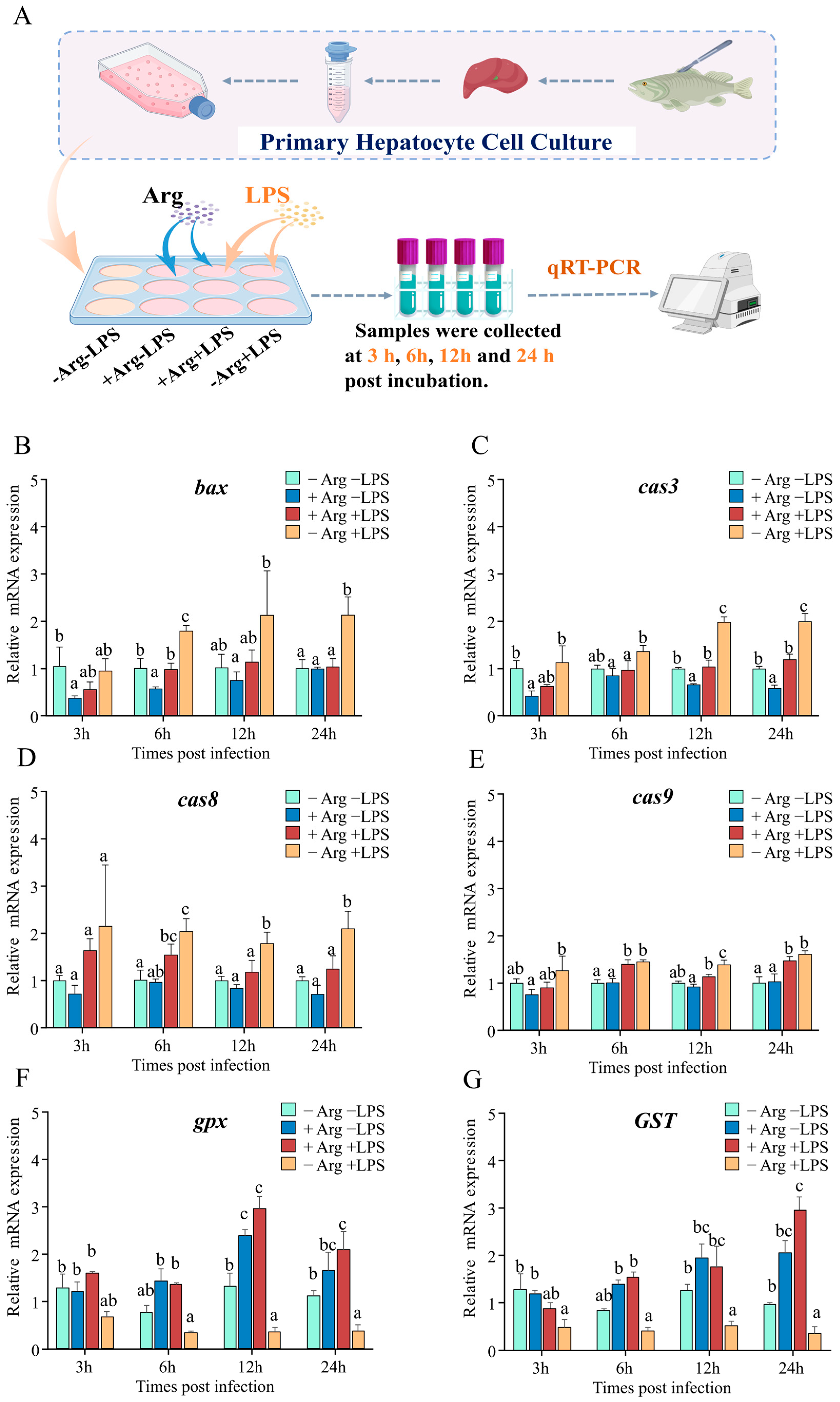
2.8. Isolation and Culture of Hepatocytes
3. Results
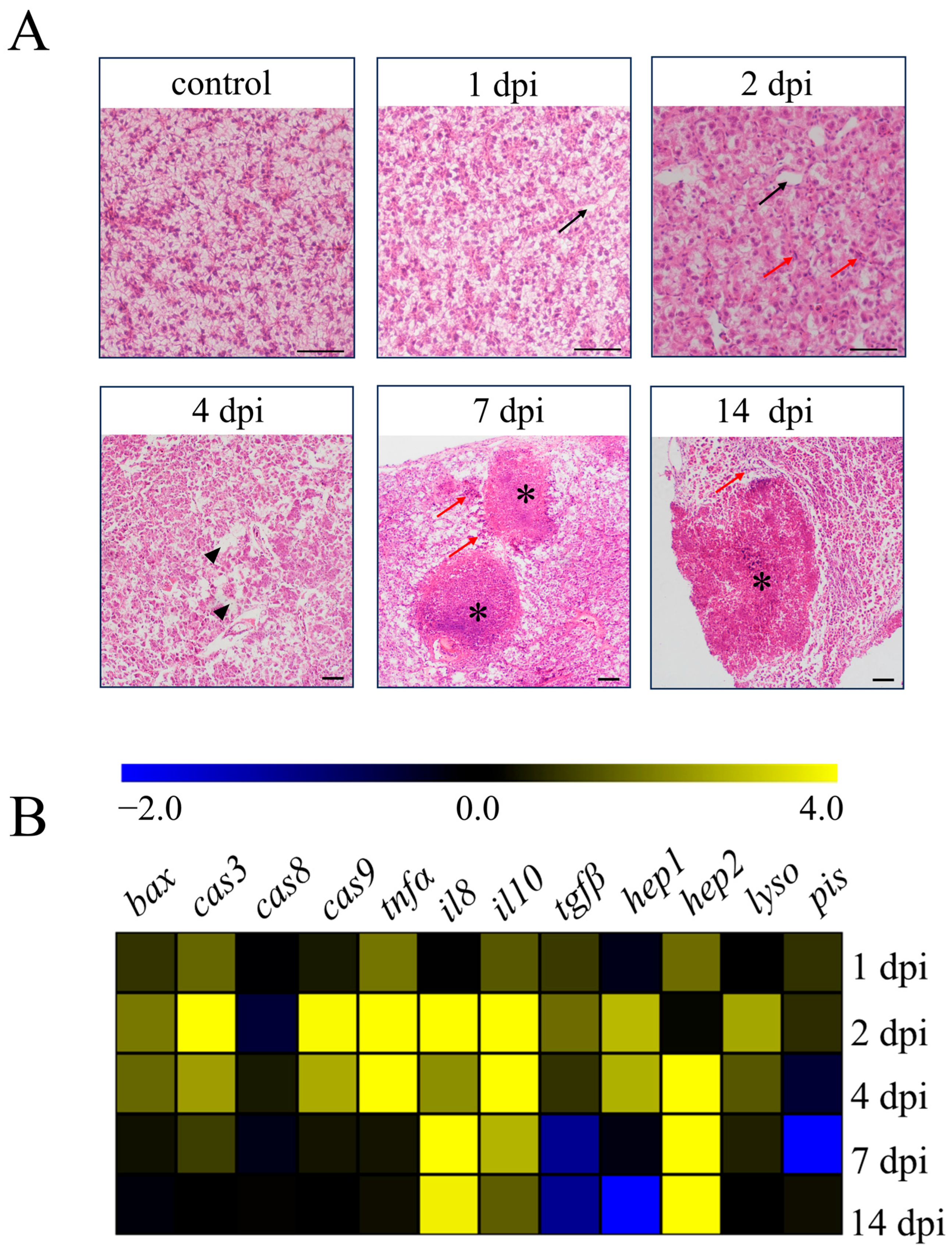
3.1. Pathological Changes and Gene Expression of Liver
3.2. DEG Analyses in the Liver Following N. seriolae Infection
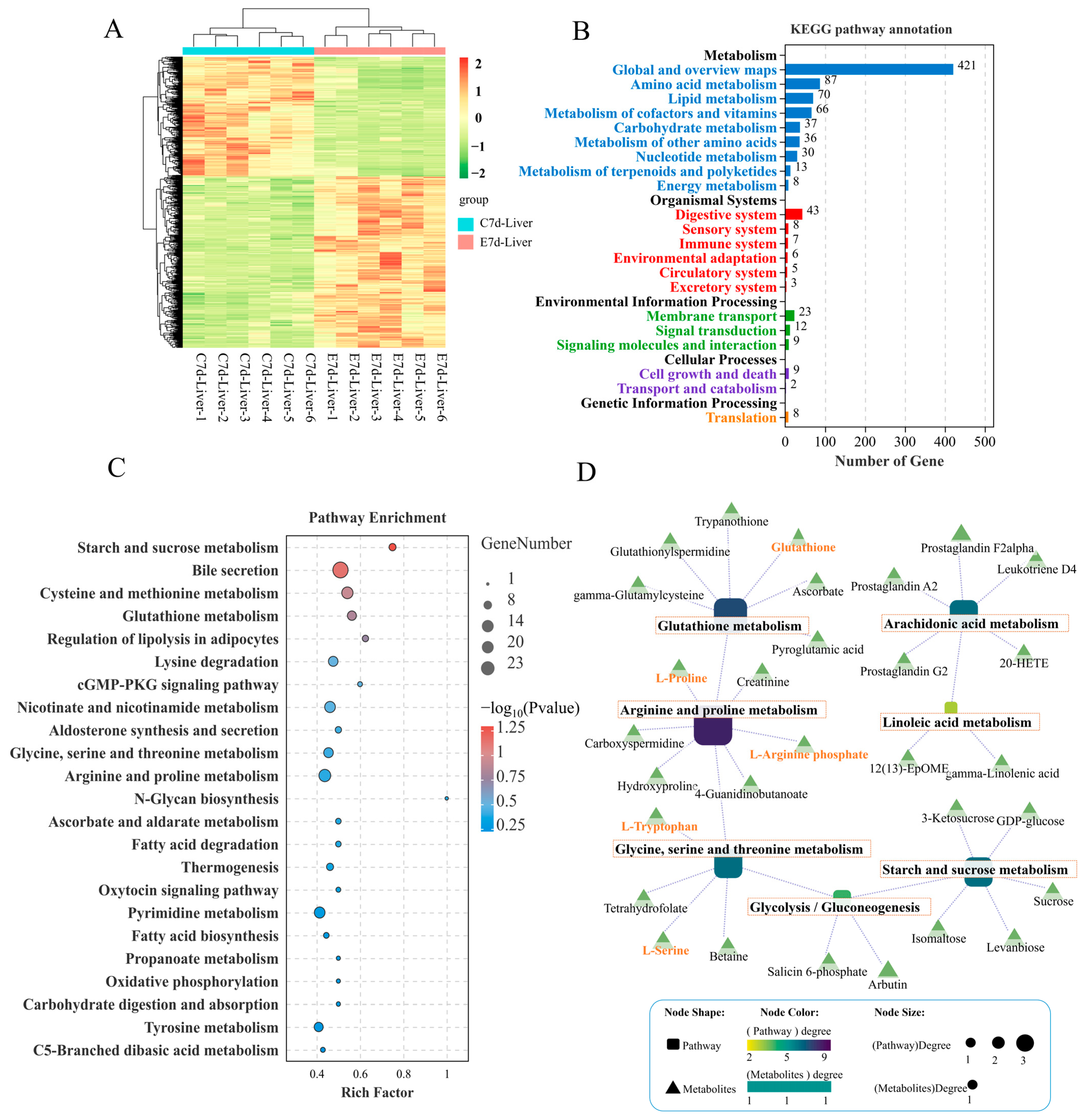
3.3. Change of Metabolome Profiles in the Liver Following N. seriolae Infection
3.4. Joint Analysis of Transcriptome and Metabolome
3.5. Amino Acid-Related Metabolic Changes in the Liver
3.6. Lipid-Related Metabolic Changes in the Liver
3.7. Carbohydrate-Related Metabolic Changes in the Liver
3.8. Arginine Suppresses Apoptosis Genes and Enhances Antioxidant Gene Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| DEGs | Differentially expressed genes | gpx | glutathione peroxidase |
| DMs | Differentially expressed metabolites | GST | glutathione-S-transferase |
| KEGG | Kyoto Encyclopedia of Genes and Genomes | CYP1A1 | cytochrome p450 family 1 subfamily a1 |
| GO | Gene Ontology | DHKTD1 | 2-oxoadipate dehydrogenase E1 component |
| GSEA | Gene Set Enrichment Analysis | ALDH | aldehyde dehydrogenase (NAD+) |
| OPLS-DA | orthogonal partial least squares discriminant analysis | HNMT | histamine n-methyltransferase |
| VIP | variable importance in the projection | ahcY | adenosylhomocysteinase |
| DMEM | Dulbecco’s Modified Eagle Medium | glyA | glycine hydroxymethyltransferase |
| PS | penicillin-streptomycin | serB | phosphoserine phosphatase |
| FBS | fetal bovine serum | GLDC | glycine dehydrogenase |
| LPS | lipopolysaccharide | TDO2 | tryptophan 2,3-dioxygenase |
| H&E | hematoxylin and eosin | TST | thiosulfate/3-mercaptopyruvate sulfurtransferase |
| qPCR | quantitative polymerase chain reaction | DAO | d-amino-acid oxidase |
| LC-MS | liquid chromatography-mass spectrometry | PGAM | 2,3-bisphosphoglycerate-dependent phosphoglycerate mutase |
| GSH | glutathione | Lipid metabolism-related genes/protein | |
| Arg | arginine | CYP2J | cytochrome P450 family 2 subfamily J |
| Immune and antioxidant-related genes | PLA2G | secretory phospholipase A2 | |
| bax | bcl-2-associated x protein | CYP7A1 | cholesterol 7alpha-monooxygenase |
| cas3/8/9 | caspase-3/8/9 | UGT | glucuronosyltransferase |
| tnfa | tumor necrosis factor-alpha | ACOT1_2_4 | acyl-coenzyme A thioesterase 1/2/4 |
| il8/il10 | interleukin-8/10 | ELOVL6 | elongation of very long chain fatty acids protein 6 |
| tgfβ | transforming growth factor beta | FADS2 | acyl- CoA 6-desaturase |
| hepl/hep2 | hepcidin-1/2 | Carbohydrate metabolism-related genes/protein | |
| lyso | lysozyme | ACSS1_2 | acetyl- CoA synthetase |
| pis | piscidin | pgm | phosphoglucomutase |
| sod1 | superoxide dismutase 1 | MINPP1 | multiple inositol-polyphosphate phosphatase |
| nod1 | nucleotide-binding oligomerization domain-containing protein 1 | LDH | l-lactate dehydrogenase |
| odc1 | ornithine decarboxylase 1 | PYG | glycogen phosphorylase |
| arg2 | arginase 2 | UGP2 | UTP-glucose-1-phosphate uridylyltransferase |
| c7b | complement component 7b | GCK | glucokinase |
| ass1 | argininosuccinate synthase 1 | glmS | methylaspartate mutase sigma subunit |
| asl | argininosuccinate lyase | AMY | alpha-amylase |
| myd88 | myeloid differentiation primary response 88 | GAA | lysosomal alpha-glucosidase |
References
- Trefts, E.; Gannon, M.; Wasserman, D.H. The liver. Curr. Biol. 2017, 27, R1147–R1151. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, E.; Baird, A.W.; O’Farrelly, C. Microanatomy of the liver immune system. In Seminars in Immunopathology; Springer: Berlin, Germany, 2009; Volume 31, pp. 333–343. [Google Scholar]
- Kubes, P.; Jenne, C. Immune responses in the liver. Annu. Rev. Immunol. 2018, 36, 247–277. [Google Scholar] [CrossRef]
- Möller, A.-M.; Korytář, T.; Kollner, B.; Schmidt-Posthaus, H.; Segner, H. The teleostean liver as an immunological organ: Intrahepatic immune cells (IHICs) in healthy and benzo[a]pyrene challenged rainbow trout (Oncorhynchus mykiss). Dev. Comp. Immunol. 2014, 46, 518–529. [Google Scholar] [CrossRef]
- Castro, R.; Abós, B.; Pignatelli, J.; von Gersdorff Jørgensen, L.; Gonzalez Granja, A.; Buchmann, K.; Tafalla, C. Early immune responses in rainbow trout liver upon viral hemorrhagic septicemia virus (VHSV) infection. PLoS ONE 2014, 9, e111084. [Google Scholar] [CrossRef]
- Falfushynska, H.; Kasianchuk, N.; Siemens, E.; Henao, E.; Rzymski, P. A review of common cyanotoxins and their effects on fish. Toxics 2023, 11, 118. [Google Scholar] [CrossRef]
- Singh, G.; Sharma, S. Heavy metal contamination in fish: Sources, mechanisms and consequences. Aquat. Sci. 2024, 86, 107. [Google Scholar] [CrossRef]
- Dong, F.; Sun, Y.-L.; Qian, Y.-X.; Chen, Q.; He, J.-L.; Wang, J.-T.; Han, T.; Zhang, X.-M.; Deng, Y.-T. Integrated analysis of transcriptome and metabolome reveals the regulatory mechanism of largemouth bass (Micropterus salmoides) in response to Nocardia seriolae infection. Fish Shellfish Immunol. 2024, 145, 109322. [Google Scholar] [CrossRef]
- Ning, X.; Peng, Y.; Tang, P.; Zhang, Y.; Wang, L.; Zhang, W.; Zhang, K.; Ji, J.; Yin, S. Integrated analysis of transcriptome and metabolome reveals distinct responses of Pelteobagrus fulvidraco against Aeromonas veronii infection at invaded and recovering stage. Int. J. Mol. Sci. 2022, 23, 10121. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, H.; Zhang, J.; Liu, R.; Zhao, H.; Shan, S.; Yang, G. Identification of three inflammatory caspases in common carp (Cyprinus carpio L.) and its role in immune response against bacterial infection. Fish Shellfish Immunol. 2022, 131, 590–601. [Google Scholar]
- Wang, Z.; Zhai, W.; Liu, H. Megalobrama amblycephala IL-22 attenuates Aeromonas hydrophila induced inflammation, apoptosis and tissue injury by regulating the ROS/NLRP3 inflammasome axis. Front. Immunol. 2024, 15, 1447431. [Google Scholar] [CrossRef]
- Maekawa, S.; Yoshida, T.; Wang, P.C.; Chen, S.C. Current knowledge of nocardiosis in teleost fish. J. Fish Dis. 2018, 41, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Zhang, H.; Dong, Z.; Wang, W. Molecular identification of Nocardia seriolae and comparative analysis of spleen transcriptomes of hybrid snakehead (Channa maculata ♀ × Channa argus ♂) with nocardiosis disease. Front. Immunol. 2022, 13, 778915. [Google Scholar] [CrossRef] [PubMed]
- Labrie, L.; Ng, J.; Tan, Z.; Komar, C.; Ho, E.; Grisez, L. Nocardial infections in fish: An emerging problem in both freshwater and marine aquaculture systems in Asia. In Diseases in Asian Aquaculture VI; Fish Health Section, Asian Fisheries Society: Manila, Philippines, 2008; pp. 297–312. [Google Scholar]
- Elkesh, A.; Kantham, K.; Shinn, A.; Crumlish, M.; Richards, R. Systemic nocardiosis in a Mediterranean population of cultured meagre, Argyrosomus regius Asso (Perciformes: Sciaenidae). J. Fish Dis. 2013, 36, 141–149. [Google Scholar] [CrossRef]
- Liu, W.; Deng, Y.; Tan, A.; Zhao, F.; Chang, O.; Wang, F.; Lai, Y.; Huang, Z. Intracellular behavior of Nocardia seriolae and its apoptotic effect on RAW264.7 macrophages. Front. Cell. Infect. Microbiol. 2023, 13, 1138422. [Google Scholar] [CrossRef]
- Fishery Administration, Bureau of Fisheries, Ministry of Agriculture and Rural Affairs, P.R. China. China Fishery Statistical Yearbook; China Agriculture Press: Beijing, China, 2023. [Google Scholar]
- Zhang, W.; Zhou, K.; Huang, L.; Yang, N.; Lin, L.; Chen, L.; Yao, J.; Dong, M.; Shen, J.; Pan, X. Biological characteristics and pathogenicity comparison of Nocardia seriolae isolated from Micropterus salmoides and Channa argus. Front. Vet. Sci. 2024, 11, 1367066. [Google Scholar] [CrossRef]
- Lei, X.; Zhao, R.; Geng, Y.; Wang, K.; Yang, P.O.; Chen, D.; Huang, X.; Zuo, Z.; He, C.; Chen, Z. Nocardia seriolae: A serious threat to the largemouth bass Micropterus salmoides industry in Southwest China. Dis. Aquat. Org. 2020, 142, 13–21. [Google Scholar] [CrossRef]
- Horta, M.A.C.; Pimenta, R.J.G.; Almeida, D.A.; Rosolen, R.R.; Aono, A.H.; Jaire Filho, F.; de Oliveira, F.A.; Niederauer, G.F.; Ferreira, R.C.U.; Bajay, S.K. Transcriptomic analysis of genes: Expression and regulation. In Transcriptome Profiling; Academic Press: Cambridge, MA, USA, 2023; pp. 1–41. [Google Scholar]
- Byadgi, O.; Chen, C.-W.; Wang, P.-C.; Tsai, M.-A.; Chen, S.-C. De novo transcriptome analysis of differential functional gene expression in largemouth bass (Micropterus salmoides) after challenge with Nocardia seriolae. Int. J. Mol. Sci. 2016, 17, 1315. [Google Scholar] [CrossRef]
- Chen, J.; Li, Y.; Wang, W.; Xia, L.; Wang, Z.; Hou, S.; Huang, J.; Lu, Y. Transcriptome analysis of immune-related gene expression in hybrid snakehead (Channa maculata ♀ × Channa argus ♂) after challenge with Nocardia seriolae. Fish Shellfish Immunol. 2018, 81, 476–484. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.-X.; Ma, Y.-M.; Li, H.; Peng, X.-X. N-acetylglucosamine enhances survival ability of tilapias infected by Streptococcus iniae. Fish Shellfish Immunol. 2014, 40, 524–530. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, C.; Sun, Y.; Chen, S.; Zhou, J.; Han, T.; Wang, J.; Harpaz, S. An integrated analysis of transcriptome and metabolome reveals three BCAAs relieve lipid accumulation by inhibiting lipid synthesis and promoting lipid oxidation in the liver of largemouth bass (Micropterus salmoides). Aquaculture 2024, 581, 740384. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, L.; Zhou, E.; Li, J.; Wu, X.; Zhong, X.; Lv, H.; Yi, S.; Gao, Q.; Yang, Z. Integrated comparative transcriptome and weighted gene co-expression network analysis provide valuable insights into the response mechanisms of crayfish (Procambarus clarkii) to copper stress. J. Hazard. Mater. 2023, 448, 130820. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-Y.; Liu, Y.; Jiang, W.-D.; Jiang, J.; Wu, P.; Zhao, J.; Kuang, S.-Y.; Tang, L.; Tang, W.-N.; Zhang, Y.-A. Co- and post-treatment with lysine protects primary fish enterocytes against Cu-induced oxidative damage. PLoS ONE 2016, 11, e0147408. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-Y.; Tang, L.; Hu, K.; Liu, Y.; Jiang, W.-D.; Jiang, J.; Wu, P.; Chen, G.-F.; Li, S.-H.; Kuang, S.-Y. Effect of dietary lysine on growth, intestinal enzymes activities and antioxidant status of sub-adult grass carp (Ctenopharyngodon idella). Fish Physiol. Biochem. 2014, 40, 659–671. [Google Scholar] [CrossRef]
- Wang, B.; Feng, L.; Chen, G.-F.; Jiang, W.-D.; Liu, Y.; Kuang, S.-Y.; Jiang, J.; Tang, L.; Wu, P.; Tang, W.-N. Jian carp (Cyprinus carpio var. Jian) intestinal immune responses, antioxidant status and tight junction protein mRNA expression are modulated via Nrf2 and PKC in response to dietary arginine deficiency. Fish Shellfish Immunol. 2016, 51, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Liu, Y.; Jiang, J.; Jiang, W.; Kuang, S.; Tang, L.; Tang, W.; Zhang, Y.-A.; Zhou, X.; Feng, L. Effect of dietary arginine on the immune response and gene expression in head kidney and spleen following infection of Jian carp with Aeromonas hydrophila. Fish Shellfish Immunol. 2015, 44, 195–202. [Google Scholar] [CrossRef]
- Siddik, M.A.; Howieson, J.; Islam, S.M.; Fotedar, R. Symbiotic feed supplementation improves antioxidant response and innate immunity of juvenile barramundi (Lates calcarifer) subjected to bacterial infection. Aquaculture 2022, 552, 737965. [Google Scholar] [CrossRef]
- Li, X.; Han, T.; Zheng, S.; Wu, G. Nutrition and functions of amino acids in aquatic crustaceans. Adv. Exp. Med. Biol. 2021, 1285, 169–198. [Google Scholar]
- Liang, H.; Ji, K.; Ge, X.; Ren, M.; Liu, B.; Xi, B.; Pan, L. Effects of dietary arginine on antioxidant status and immunity involved in AMPK-NO signaling pathway in juvenile blunt snout bream. Fish Shellfish Immunol. 2018, 78, 69–78. [Google Scholar] [CrossRef]
- Evoy, D.; Lieberman, M.D.; Fahey III, T.J.; Daly, J.M. Immunonutrition: The role of arginine. Nutrition 1998, 14, 611–617. [Google Scholar] [CrossRef]
- Eddy, F.; Tibbs, P. Effects of nitric oxide synthase inhibitors and a substrate, L-arginine, on the cardiac function of juvenile salmonid fish. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2003, 135, 137–144. [Google Scholar] [CrossRef]
- Council, N.R. Nutrient Requirements of Fish and Shrimp; National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Tan, B.; Yin, Y.; Kong, X.; Li, P.; Li, X.; Gao, H.; Li, X.; Huang, R.; Wu, G. L-Arginine stimulates proliferation and prevents endotoxin-induced death of intestinal cells. Amino Acids 2010, 38, 1227–1235. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Shi, D.; Zhou, X.-Q.; Hu, Y.; Feng, L.; Liu, Y.; Jiang, W.-D.; Zhao, Y. In vitro and in vivo protective effect of arginine against lipopolysaccharide-induced inflammatory response in the intestine of juvenile Jian carp (Cyprinus carpio var. Jian). Fish Shellfish Immunol. 2015, 42, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Petrovic, V.; Buzadzic, B.; Korac, A.; Vasilijevic, A.; Jankovic, A.; Micunovic, K.; Korac, B. Antioxidative defence alterations in skeletal muscle during prolonged acclimation to cold: Role of L-arginine/NO-producing pathway. J. Exp. Biol. 2008, 211, 114–120. [Google Scholar] [CrossRef]
- Wang, B.; Liu, Y.; Feng, L.; Jiang, W.-D.; Kuang, S.-Y.; Jiang, J.; Li, S.-H.; Tang, L.; Zhou, X.-Q. Effects of dietary arginine supplementation on growth performance, flesh quality, muscle antioxidant capacity and antioxidant-related signalling molecule expression in young grass carp (Ctenopharyngodon idella). Food Chem. 2015, 167, 91–99. [Google Scholar] [CrossRef]
- Qian, Y.-X.; Zhou, F.-F.; Chen, Q.; Dong, F.; Xu, H.-Y.; Sun, Y.-L.; Wang, J.-T.; Han, T. Arginine alleviates LPS-induced leukocytes inflammation and apoptosis via adjusted NODs signaling. Fish Shellfish Immunol. 2024, 154, 109985. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Wang, C.; Lou, C.; Yang, Z.; Shi, J.; Niu, N. Plasma metabolomic analysis reveals the metabolic characteristics and potential diagnostic biomarkers of spinal tuberculosis. Heliyon 2024, 10, e27940. [Google Scholar] [CrossRef]
- Gong, C.; Guo, G.; Pan, B.; Gao, C.; Zhu, X.; Liu, J.; Wang, S.; Diao, W. Global transcription and metabolic profiles of five tissues in pepper fruits. Sci. Data 2024, 11, 1129. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, T.; Shen, X.; Liu, J.; Zhao, D.; Sun, Y.; Wang, L.; Liu, Y.; Gong, X.; Liu, Y. Serum metabolomics for early diagnosis of esophageal squamous cell carcinoma by UHPLC-QTOF/MS. Metabolomics 2016, 12, 116. [Google Scholar] [CrossRef]
- Thevenot, E.A.; Roux, A.; Xu, Y.; Ezan, E.; Junot, C. Analysis of the human adult urinary metabolome variations with age, body mass index, and gender by implementing a comprehensive workflow for univariate and OPLS statistical analyses. J. Proteome Res. 2015, 14, 3322–3335. [Google Scholar] [CrossRef] [PubMed]
- Starai, V.; Escalante-Semerena, J. Acetyl-coenzyme A synthetase (AMP forming). Cell. Mol. Life Sci. 2004, 61, 2020–2030. [Google Scholar] [CrossRef]
- Zhao, X.; Li, L.; Li, C.; Liu, E.; Zhu, H.; Ling, Q. Heat stress-induced endoplasmic reticulum stress promotes liver apoptosis in largemouth bass (Micropterus salmoides). Aquaculture 2022, 546, 737401. [Google Scholar] [CrossRef]
- Campos-Sanchez, J.C.; Esteban, M.A. Review of inflammation in fish and value of the zebrafish model. J. Fish Dis. 2021, 44, 123–139. [Google Scholar] [CrossRef]
- Zhang, X.-T.; Yu, Y.-Y.; Xu, H.-Y.; Huang, Z.-Y.; Liu, X.; Cao, J.-F.; Meng, K.-F.; Wu, Z.-B.; Han, G.-K.; Zhan, M.-T. Prevailing role of mucosal Igs and B cells in teleost skin immune responses to bacterial infection. J. Immunol. 2021, 206, 1088–1101. [Google Scholar] [CrossRef]
- Iyer, S.S.; Cheng, G. Role of interleukin 10 transcriptional regulation in inflammation and autoimmune disease. Crit. Rev. Immunol. 2012, 32, 23–63. [Google Scholar] [CrossRef] [PubMed]
- Boackle, S.A. Complement and autoimmunity. Biomed. Pharmacother. 2003, 57, 269–273. [Google Scholar] [CrossRef]
- Sunyer, J.O.; Boshra, H.; Lorenzo, G.; Parra, D.; Freedman, B.; Bosch, N. Evolution of complement as an effector system in innate and adaptive immunity. Immunol. Res. 2003, 27, 549–564. [Google Scholar] [CrossRef]
- Boshra, H.; Li, J.; Sunyer, J. Recent advances on the complement system of teleost fish. Fish Shellfish Immunol. 2006, 20, 239–262. [Google Scholar] [CrossRef]
- Li, M.-F.; Zhang, H.-Q. An overview of complement systems in teleosts. Dev. Comp. Immunol. 2022, 137, 104520. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Ding, M.; Li, Y.; Zhong, X.; Liu, S.; Guo, Z.; Yin, X.; Fu, S.; Ye, J. The complement component 1 q (C1q) in Nile tilapia (Oreochromis niloticus): Functional characterization in host defense against bacterial infection and effect on cytokine response in macrophages. Dev. Comp. Immunol. 2018, 87, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Wang, G.-H.; Yue, B.; Wang, J.-J.; Gu, Q.-Q.; Zhou, S.; Zhang, M.; Hu, Y.-H. A novel C1q domain containing protein in black rockfish (Sebastes schlegelii) serves as a pattern recognition receptor with immunoregulatory properties and possesses binding activity to heat-aggregated IgG. Fish Shellfish Immunol. 2019, 87, 73–81. [Google Scholar] [CrossRef]
- Wang, L.; Fan, C.; Xu, W.; Zhang, Y.; Dong, Z.; Xiang, J.; Chen, S. Characterization and functional analysis of a novel C1q-domain-containing protein in Japanese flounder (Paralichthys olivaceus). Dev. Comp. Immunol. 2017, 67, 322–332. [Google Scholar] [CrossRef]
- Wu, M.; Jia, B.-B.; Li, M.-F. Complement C3 and activated fragment C3a are involved in complement activation and anti-bacterial immunity. Front. Immunol. 2022, 13, 813173. [Google Scholar] [CrossRef]
- Ma, Z.-Y.; Liang, J.-X.; Li, W.-S.; Sun, Y.; Wu, C.-S.; Hu, Y.-Z.; Li, J.; Zhang, Y.-A.; Zhang, X.-J. Complement C3a enhances the phagocytic activity of B cells through C3aR in a fish. Front. Immunol. 2022, 13, 873982. [Google Scholar] [CrossRef]
- Troisi, R.; Balasco, N.; Autiero, I.; Sica, F.; Vitagliano, L. New insight into the traditional model of the coagulation cascade and its regulation: Illustrated review of a three-dimensional view. Res. Pract. Thromb. Haemost. 2023, 7, 100023. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Kong, J.; Huang, J.; Zhou, L.; Jiang, Y.; Miao, R.; Yin, F. Integration of metabolomic and transcriptomic analyses to characterize the influence of the gill metabolism of Nibea albiflora on the response to Cryptocaryon irritans infection. Vet. Parasitol. 2021, 298, 109533. [Google Scholar] [CrossRef]
- Yang, L.; Chu, Z.; Liu, M.; Zou, Q.; Li, J.; Liu, Q.; Wang, Y.; Wang, T.; Xiang, J.; Wang, B. Amino acid metabolism in immune cells: Essential regulators of the effector functions, and promising opportunities to enhance cancer immunotherapy. J. Hematol. Oncol. 2023, 16, 59. [Google Scholar] [CrossRef]
- Wu, G.; Bazer, F.W.; Davis, T.A.; Kim, S.W.; Li, P.; Marc Rhoads, J.; Carey Satterfield, M.; Smith, S.B.; Spencer, T.E.; Yin, Y. Arginine metabolism and nutrition in growth, health and disease. Amino Acids 2009, 37, 153–168. [Google Scholar] [CrossRef]
- Hoseini, S.M.; Ahmad Khan, M.; Yousefi, M.; Costas, B. Roles of arginine in fish nutrition and health: Insights for future researches. Rev. Aquac. 2020, 12, 2091–2108. [Google Scholar] [CrossRef]
- Wang, Q.; Xu, Z.; Ai, Q. Arginine metabolism and its functions in growth, nutrient utilization, and immunonutrition of fish. Anim. Nutr. 2021, 7, 716–727. [Google Scholar] [CrossRef]
- Cheng, Z.; Gatlin III, D.M.; Buentello, A. Dietary supplementation of arginine and/or glutamine influences growth performance, immune responses and intestinal morphology of hybrid striped bass (Morone chrysops × Morone saxatilis). Aquaculture 2012, 362, 39–43. [Google Scholar] [CrossRef]
- Pohlenz, C.; Buentello, A.; Mwangi, W.; Gatlin III, D.M. Arginine and glutamine supplementation to culture media improves the performance of various channel catfish immune cells. Fish Shellfish Immunol. 2012, 32, 762–768. [Google Scholar] [CrossRef]
- Chen, S.; Xia, Y.; He, F.; Fu, J.; Xin, Z.; Deng, B.; He, L.; Zhou, X.; Ren, W. Serine supports IL-1β production in macrophages through mTOR signaling. Front. Immunol. 2020, 11, 1866. [Google Scholar] [CrossRef]
- Rodriguez, A.E.; Ducker, G.S.; Billingham, L.K.; Martinez, C.A.; Mainolfi, N.; Suri, V.; Friedman, A.; Manfredi, M.G.; Weinberg, S.E.; Rabinowitz, J.D. Serine metabolism supports macrophage IL-1β production. Cell Metab. 2019, 29, 1003–1011. [Google Scholar] [CrossRef]
- Matthaiou, C.M.; Goutzourelas, N.; Stagos, D.; Sarafoglou, E.; Jamurtas, A.; Koulocheri, S.D.; Haroutounian, S.A.; Tsatsakis, A.M.; Kouretas, D. Pomegranate juice consumption increases GSH levels and reduces lipid and protein oxidation in human blood. Food Chem. Toxicol. 2014, 73, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Bansal, V.; Ochoa, J.B. Arginine availability, arginase, and the immune response. Curr. Opin. Clin. Nutr. Metab. Care 2003, 6, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Morris, S.M., Jr. Enzymes of arginine metabolism. J. Nutr. 2004, 134, 2743S–2747S. [Google Scholar] [CrossRef]
- Wu, G.; Morris, S.M., Jr. Arginine metabolism: Nitric oxide and beyond. Biochem. J. 1998, 336, 336. [Google Scholar] [CrossRef]
- Liang, M.; Wang, Z.; Li, H.; Cai, L.; Pan, J.; He, H.; Wu, Q.; Tang, Y.; Ma, J.; Yang, L. L-Arginine induces antioxidant response to prevent oxidative stress via stimulation of glutathione synthesis and activation of Nrf2 pathway. Food Chem. Toxicol. 2018, 115, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.A.; Ilies, M.A. The phospholipase A2 superfamily: Structure, isozymes, catalysis, physiologic and pathologic roles. Int. J. Mol. Sci. 2023, 24, 1353. [Google Scholar] [CrossRef]
- Cathcart, M.-C.; Lysaght, J.; Pidgeon, G.P. Eicosanoid signalling pathways in the development and progression of colorectal cancer: Novel approaches for prevention/intervention. Cancer Metastasis Rev. 2011, 30, 363–385. [Google Scholar] [CrossRef]
- Dennis, E.A.; Cao, J.; Hsu, Y.-H.; Magrioti, V.; Kokotos, G. Phospholipase A2 enzymes: Physical structure, biological function, disease implication, chemical inhibition, and therapeutic intervention. Chem. Rev. 2011, 111, 6130–6185. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Sato, H.; Taketomi, Y. Updating phospholipase A2 biology. Biomolecules 2020, 10, 1457. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Dong, R.; Chen, Z.; Hu, D.; Fu, M.; Tang, Y.; Wang, D.W.; Xu, X.; Tu, L. Endothelium-specific CYP2J2 overexpression attenuates age-related insulin resistance. Aging Cell 2018, 17, e12718. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, X.A.; Wang, D.W. The roles of CYP450 epoxygenases and metabolites, epoxyeicosatrienoic acids, in cardiovascular and malignant diseases. Adv. Drug Deliv. Rev. 2011, 63, 597–609. [Google Scholar] [CrossRef]
- Dai, M.; Wu, L.; Wang, P.; Wen, Z.; Xu, X.; Wang, D.W. CYP2J2 and its metabolites EETs attenuate insulin resistance via regulating macrophage polarization in adipose tissue. Sci. Rep. 2017, 7, 46743. [Google Scholar] [CrossRef]
- Pietrocola, F.; Galluzzi, L.; Bravo-San Pedro, J.M.; Madeo, F.; Kroemer, G. Acetyl coenzyme A: A central metabolite and second messenger. Cell Metab. 2015, 21, 805–821. [Google Scholar] [CrossRef]
- Alderton, G.K. Acetate nourishes stressed tumour cells. Nat. Rev. Cancer 2015, 15, 67. [Google Scholar] [CrossRef]
- Prochownik, E.V.; Wang, H. The metabolic fates of pyruvate in normal and neoplastic cells. Cells 2021, 10, 762. [Google Scholar] [CrossRef]
- Martinez-Reyes, I.; Chandel, N.S. Mitochondrial TCA cycle metabolites control physiology and disease. Nat. Commun. 2020, 11, 102. [Google Scholar] [CrossRef] [PubMed]
- Boger, R.H.; Bode-Boger, S.M.; Frolich, J.C. The L-arginine-nitric oxide pathway: Role in atherosclerosis and therapeutic implications. Atherosclerosis 1996, 127, 1–11. [Google Scholar] [CrossRef]
- Bronte, V.; Zanovello, P. Regulation of immune responses by L-arginine metabolism. Nat. Rev. Immunol. 2005, 5, 641–654. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Chen, J.-H.; Zhang, H.; Qiu, W.; Liu, Q.-H.; Peng, X.; Li, Y.-N.; Yang, H.-K. Alkali treatment affects in vitro digestibility and bile acid binding activity of rice protein due to varying its ratio of arginine to lysine. Food Chem. 2012, 132, 925–930. [Google Scholar] [CrossRef]
- Xaus, J.; Comalada, M.; Valledor, A.F.; Lloberas, J.; Lopez-Soriano, F.; Argilés, J.M.; Bogdan, C.; Celada, A. LPS induces apoptosis in macrophages mostly through the autocrine production of TNF-α. Blood J. Am. Soc. Hematol. 2000, 95, 3823–3831. [Google Scholar]
- Kundu, J.K.; Shin, Y.K.; Kim, S.H.; Surh, Y.-J. Resveratrol inhibits phorbol ester-induced expression of COX-2 and activation of NF-κB in mouse skin by blocking IκB kinase activity. Carcinogenesis 2006, 27, 1465–1474. [Google Scholar] [CrossRef] [PubMed]
- Mandrekar, P.; Szabo, G. Signalling pathways in alcohol-induced liver inflammation. J. Hepatol. 2009, 50, 1258–1266. [Google Scholar] [CrossRef]
- Fan, T.-J.; Han, L.-H.; Cong, R.-S.; Liang, J. Caspase family proteases and apoptosis. Acta Biochim. Biophys. Sin. 2005, 37, 719–727. [Google Scholar] [CrossRef]
- Sakamaki, K.; Imai, K.; Tomii, K.; Miller, D.J. Evolutionary analyses of caspase-8 and its paralogs: Deep origins of the apoptotic signaling pathways. Bioessays 2015, 37, 767–776. [Google Scholar] [CrossRef]
- Porter, A.G.; Janicke, R.U. Emerging roles of caspase-3 in apoptosis. Cell Death Differ. 1999, 6, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Feng, L.; Jiang, W.-D.; Wu, P.; Kuang, S.-Y.; Jiang, J.; Tang, L.; Tang, W.-N.; Zhang, Y.-A.; Liu, Y. Copper-induced tight junction mRNA expression changes, apoptosis and antioxidant responses via NF-κB, TOR and Nrf2 signaling molecules in the gills of fish: Preventive role of arginine. Aquat. Toxicol. 2015, 158, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Tang, G.; Xiong, C.; Han, S.; Yang, C.; He, K.; Liu, Q.; Luo, J.; Luo, W.; Wang, Y. Chronic chlorpyrifos exposure induces oxidative stress, apoptosis and immune dysfunction in largemouth bass (Micropterus salmoides). Environ. Pollut. 2021, 282, 117010. [Google Scholar] [CrossRef]
- Mu, W.; Wang, X.; Wu, X.; Li, X.; Dong, Y.; Geng, L.; Ma, L.; Ye, B. The optimal arginine requirement in diets for juvenile humpback grouper, Cromileptes altivelis. Aquaculture 2020, 514, 734509. [Google Scholar] [CrossRef]
- Fournier, V.; Gouillou-Coustans, M.; Métailler, R.; Vachot, C.; Moriceau, J.; Le Delliou, H.; Huelvan, C.; Desbruyères, E.; Kaushik, S. Excess dietary arginine affects urea excretion but does not improve N utilisation in rainbow trout Oncorhynchus mykiss and turbot Psetta maxima. Aquaculture 2003, 217, 559–576. [Google Scholar] [CrossRef]
- Luan, X.; Li, X.; He, J.; Xu, H.; Feng, W.; Chen, Q.; Han, T.; Wang, J. Effects of arginine family amino acids supplementation on growth, whole-body amino acid profiles, antioxidant capacity, and gene expression of juvenile largemouth bass (Micropterus salmoides). Aquaculture 2025, 594, 741312. [Google Scholar] [CrossRef]
- Cynober, L.; Le Boucher, J.; Vasson, M.-P. Arginine metabolism in mammals. J. Nutr. Biochem. 1995, 6, 402–413. [Google Scholar] [CrossRef]
- Rath, M.; Miller, I.; Kropf, P.; Closs, E.I.; Munder, M. Metabolism via arginase or nitric oxide synthase: Two competing arginine pathways in macrophages. Front. Immunol. 2014, 5, 532. [Google Scholar] [CrossRef]
- Iaccarino, D.; Uliano, E.; Agnisola, C. Effects of arginine and/or lysine diet supplementation on nitrogen excretion in zebrafish. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2009, 154, S36. [Google Scholar] [CrossRef]
- Berge, G.E.; Bakke-McKellep, A.M.; Lied, E. In vitro uptake and interaction between arginine and lysine in the intestine of Atlantic salmon (Salmo salar). Aquaculture 1999, 179, 181–193. [Google Scholar] [CrossRef]
- Saeij, J.P.; Verburg-van Kemenade, L.B.; van Muiswinkel, W.B.; Wiegertjes, G.F. Daily handling stress reduces resistance of carp to Trypanoplasma borreli: In vitro modulatory effects of cortisol on leukocyte function and apoptosis. Dev. Comp. Immunol. 2003, 27, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Radi, R. Nitric oxide, oxidants, and protein tyrosine nitration. Proc. Natl. Acad. Sci. 2004, 101, 4003–4008. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, K.; Kashiwagi, K. Modulation of cellular function by polyamines. Int. J. Biochem. Cell Biol. 2010, 42, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Andersen, S.M.; Holen, E.; Aksnes, A.; Rønnestad, I.; Zerrahn, J.-E.; Espe, M. Dietary arginine affects energy metabolism through polyamine turnover in juvenile Atlantic salmon (Salmo salar). Br. J. Nutr. 2013, 110, 1968–1977. [Google Scholar] [CrossRef]
- Banerjee, B.; Khrystoforova, I.; Polis, B.; Zvi, I.B.; Karasik, D. Acute hypoxia elevates arginase 2 and induces polyamine stress response in zebrafish via evolutionarily conserved mechanism. Cell. Mol. Life Sci. 2022, 79, 41. [Google Scholar] [CrossRef]
- McDonald, M.; Smith, C.; Walsh, P. The physiology and evolution of urea transport in fishes. J. Membr. Biol. 2006, 212, 93–107. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.-L.; Zhang, S.-L.; Zhou, F.-F.; Qian, Y.-X.; He, Y.; Zhang, R.-Z.; Dong, F.; Chen, Q.; Xu, H.-Y.; Wang, J.-T.; et al. Arginine-Mediated Liver Immune Regulation and Antioxidant Defense in Largemouth Bass (Micropterus salmoides): Multi-Omics Insights into Metabolic Remodeling During Nocardia seriolae Infection. Antioxidants 2025, 14, 681. https://doi.org/10.3390/antiox14060681
Sun Y-L, Zhang S-L, Zhou F-F, Qian Y-X, He Y, Zhang R-Z, Dong F, Chen Q, Xu H-Y, Wang J-T, et al. Arginine-Mediated Liver Immune Regulation and Antioxidant Defense in Largemouth Bass (Micropterus salmoides): Multi-Omics Insights into Metabolic Remodeling During Nocardia seriolae Infection. Antioxidants. 2025; 14(6):681. https://doi.org/10.3390/antiox14060681
Chicago/Turabian StyleSun, Yu-Long, Shuai-Liang Zhang, Feng-Feng Zhou, Yuan-Xin Qian, Yang He, Run-Zhe Zhang, Fen Dong, Qiang Chen, Han-Ying Xu, Ji-Teng Wang, and et al. 2025. "Arginine-Mediated Liver Immune Regulation and Antioxidant Defense in Largemouth Bass (Micropterus salmoides): Multi-Omics Insights into Metabolic Remodeling During Nocardia seriolae Infection" Antioxidants 14, no. 6: 681. https://doi.org/10.3390/antiox14060681
APA StyleSun, Y.-L., Zhang, S.-L., Zhou, F.-F., Qian, Y.-X., He, Y., Zhang, R.-Z., Dong, F., Chen, Q., Xu, H.-Y., Wang, J.-T., Deng, Y.-T., & Han, T. (2025). Arginine-Mediated Liver Immune Regulation and Antioxidant Defense in Largemouth Bass (Micropterus salmoides): Multi-Omics Insights into Metabolic Remodeling During Nocardia seriolae Infection. Antioxidants, 14(6), 681. https://doi.org/10.3390/antiox14060681