
Integrated Transcriptome and Metabolome Analyses Reveal Complex Oxidative Damage Mechanisms in Rice Seedling Roots Under Different Carbonate Stresses

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Design
2.2. Measurement of Rice Seedling Growth Index
2.3. Assessment of Root Physiological Indices in Rice Seedlings
2.3.1. Plasma Membrane Integrity (MI) and Root Vigor Measurement
2.3.2. Measurement of Reactive Oxygen Species (ROS) and Lipid Peroxidation
2.3.3. Antioxidant Enzyme Activity Determination
2.3.4. Measurement of Plant Auxin (IAA) Content
2.3.5. Measurement of Reduced Ascorbic Acid (AsA) Content
2.3.6. Measurement of Glutathione (GSH and GSSG) Contents
2.3.7. Measurement of Flavonoid Content
2.4. Transcriptomic Analysis
2.5. Metabolomics Analysis
2.6. RNA Extraction and Real-Time Quantitative PCR (qRT-PCR)
2.7. Statistical Analysis
3. Results
3.1. Effect of Different Carbonate Stresses on Rice Seedling Growth and Root Injury
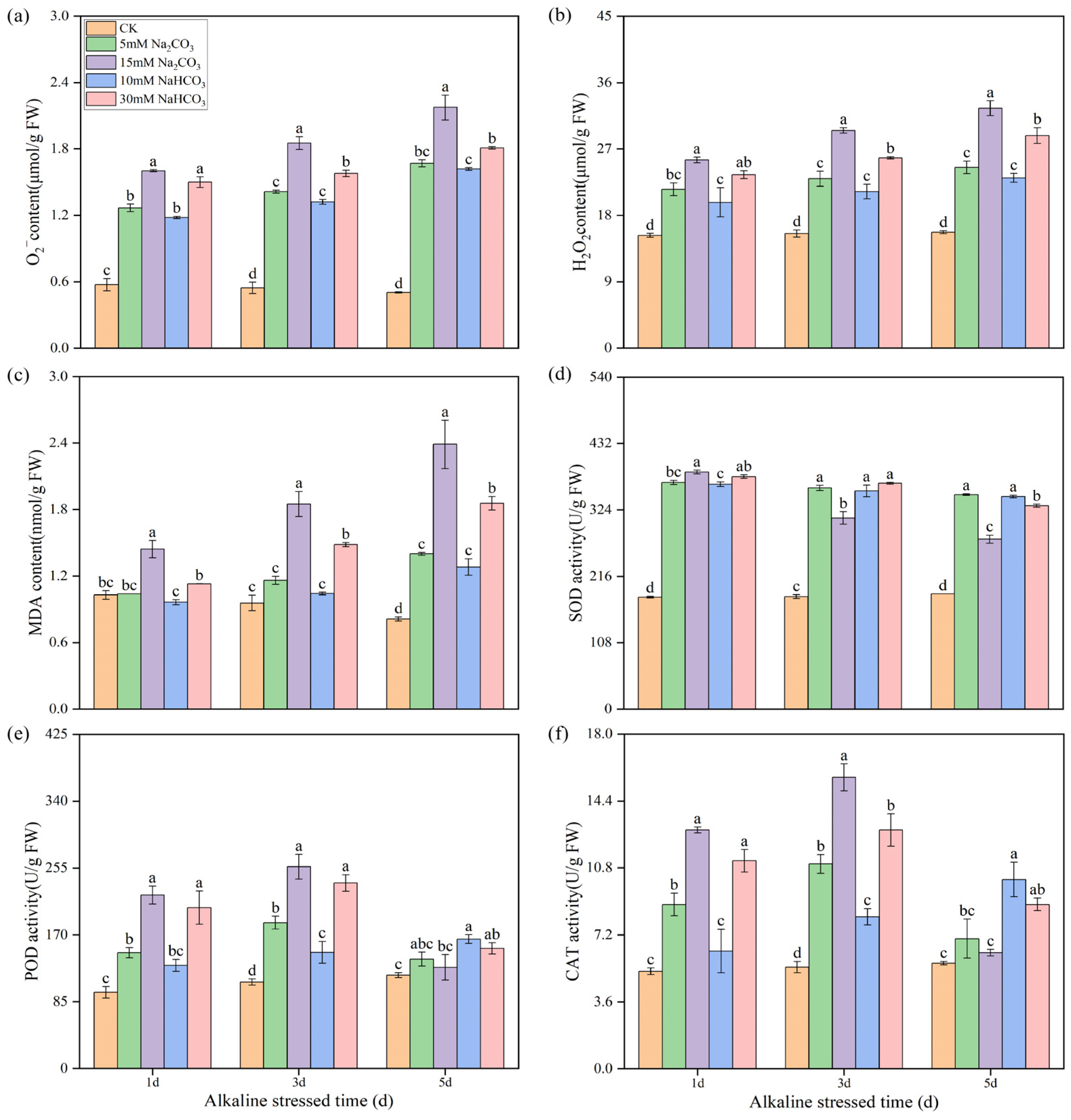
3.2. Effects of Different Carbonate Stresses on Reactive Oxygen Species Accumulation and Antioxidant Enzyme Activity in Rice Roots
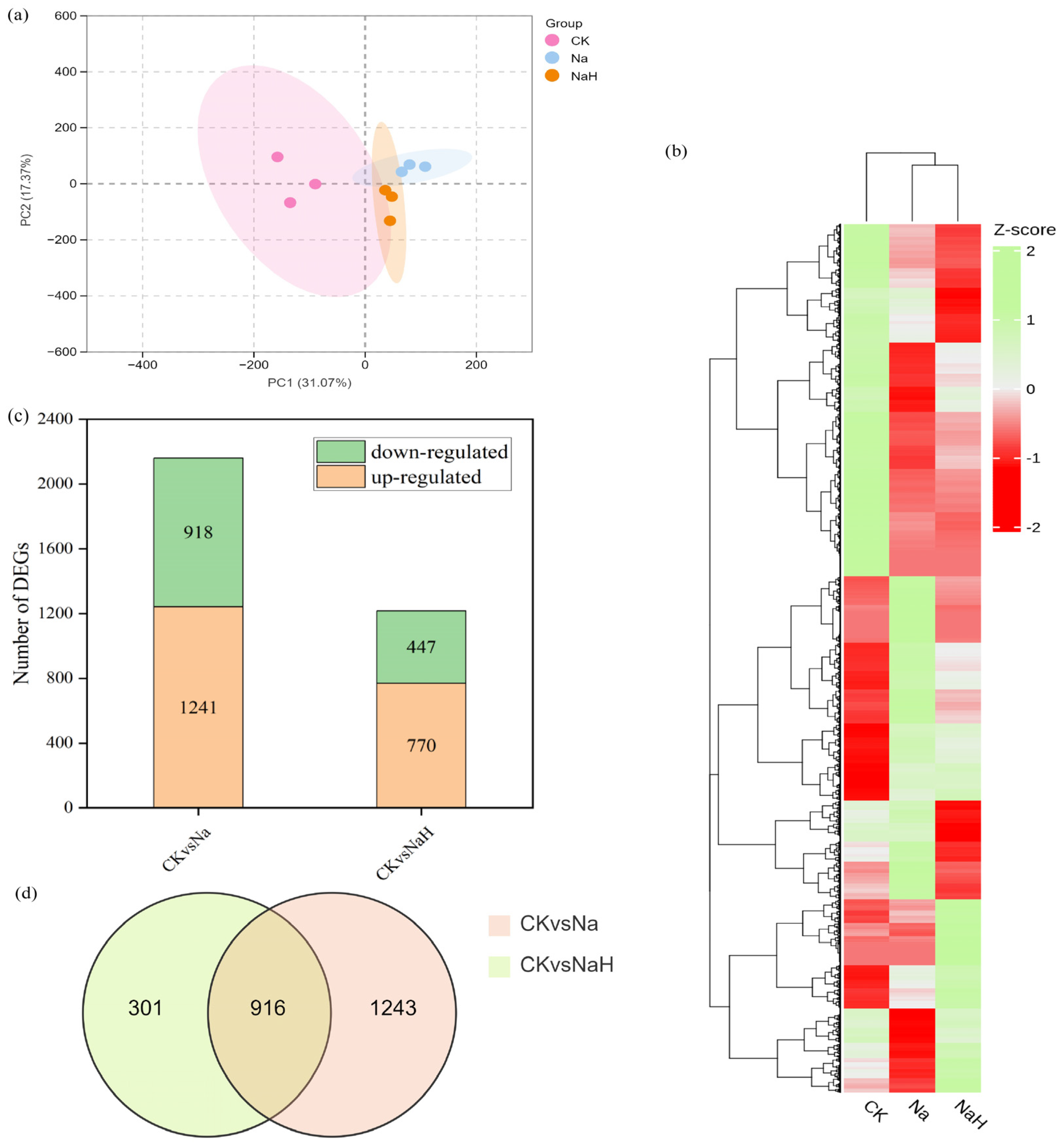
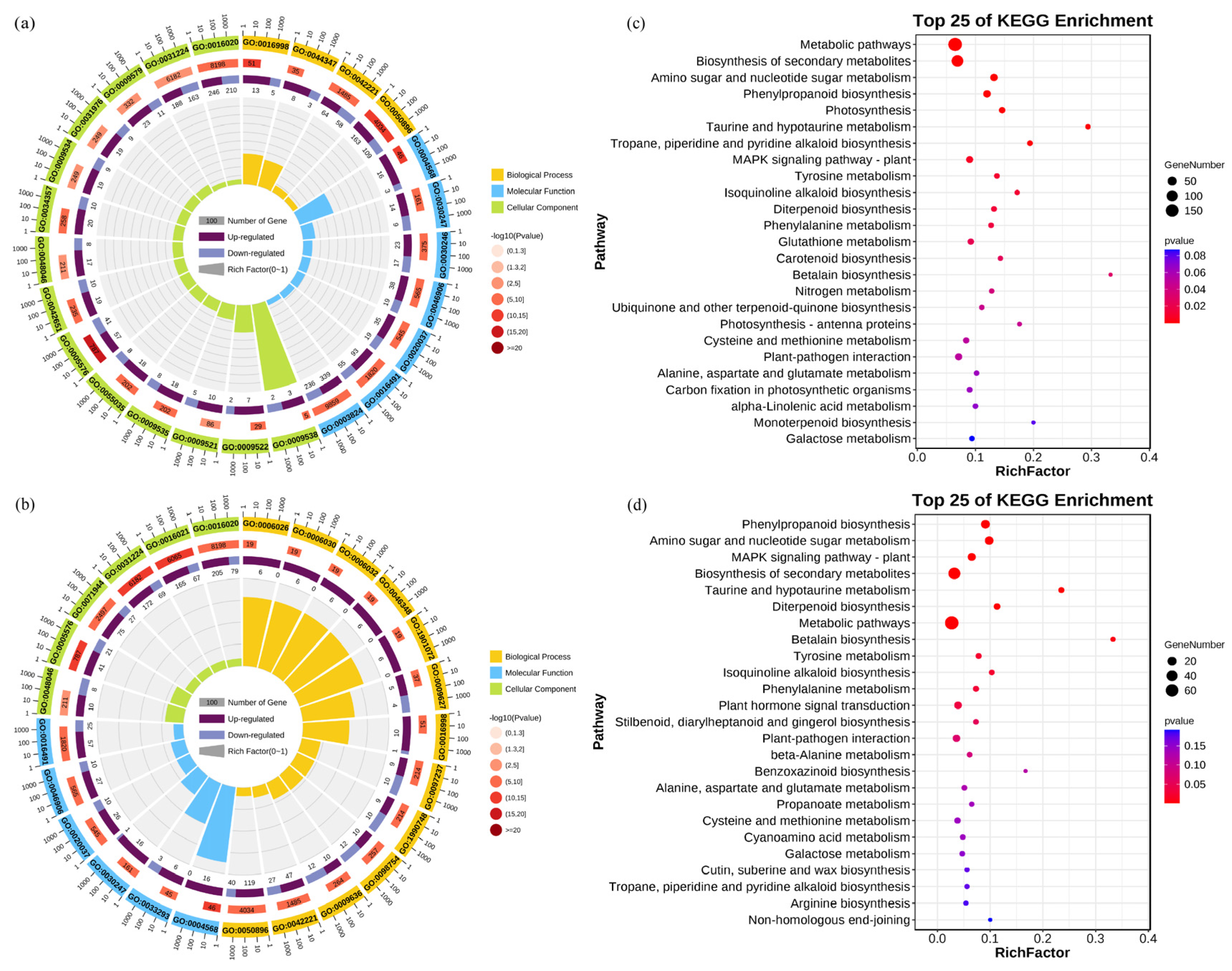
3.3. Transcriptome Profiling of Rice Seedling Roots Under Different Carbonate Stresses
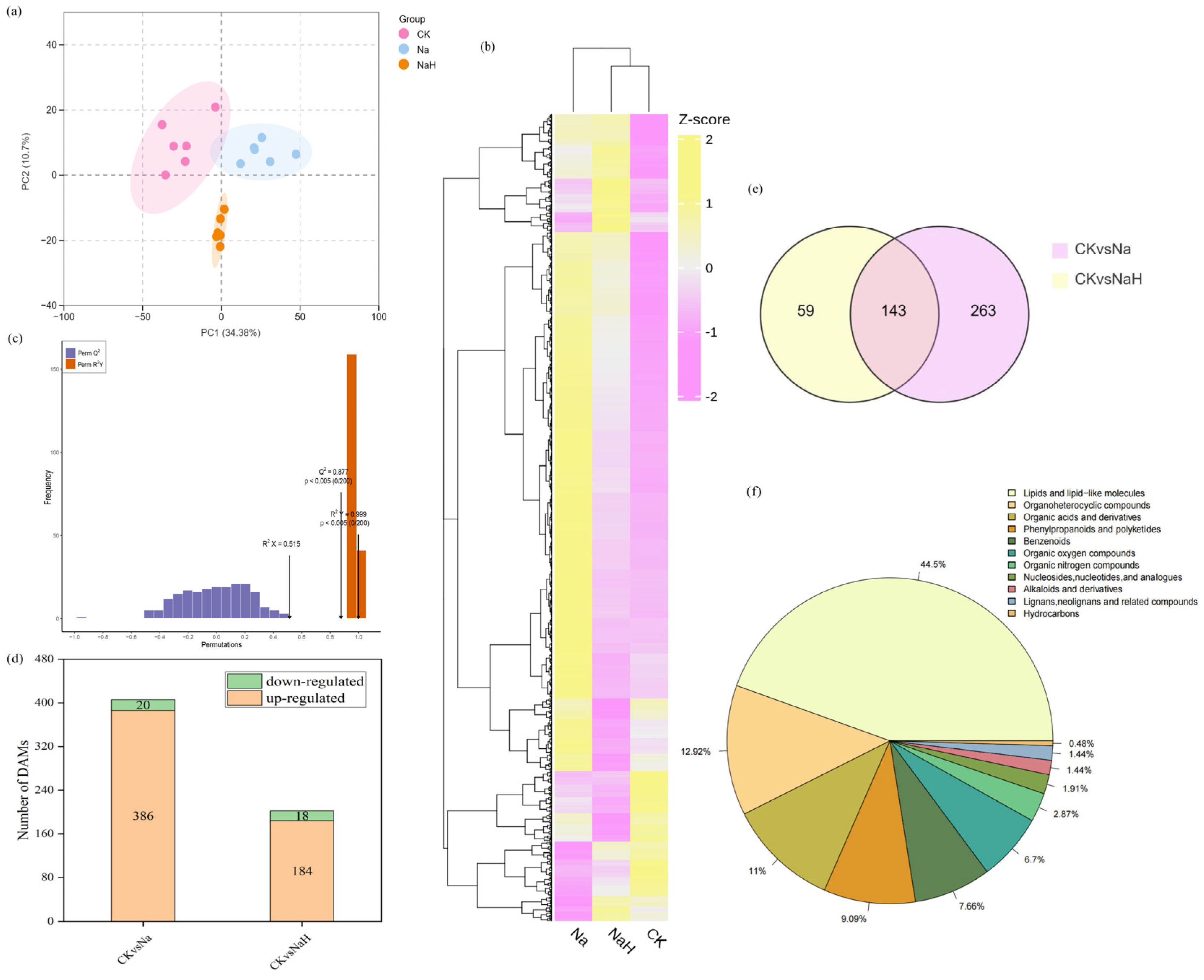
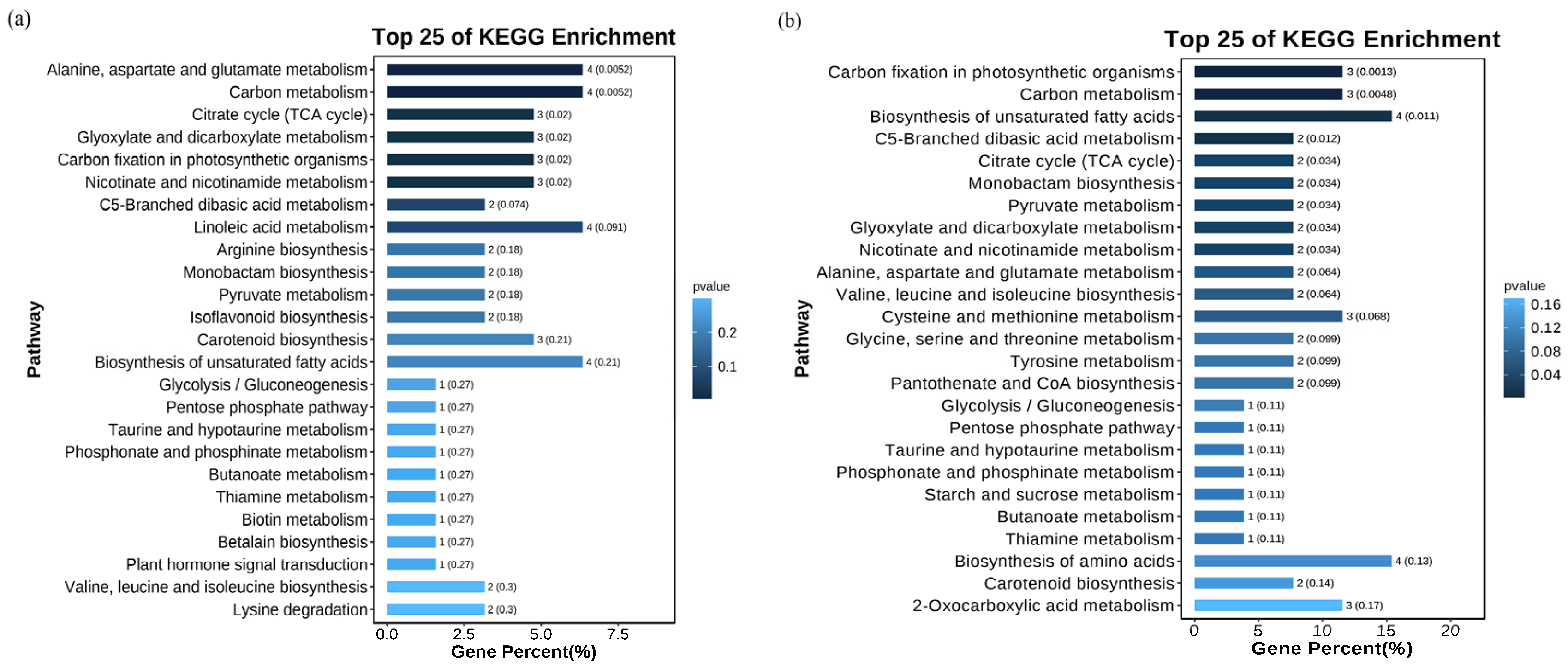
3.4. Metabolome Differences of Rice Seedling Roots Under Different Carbonate Stresses
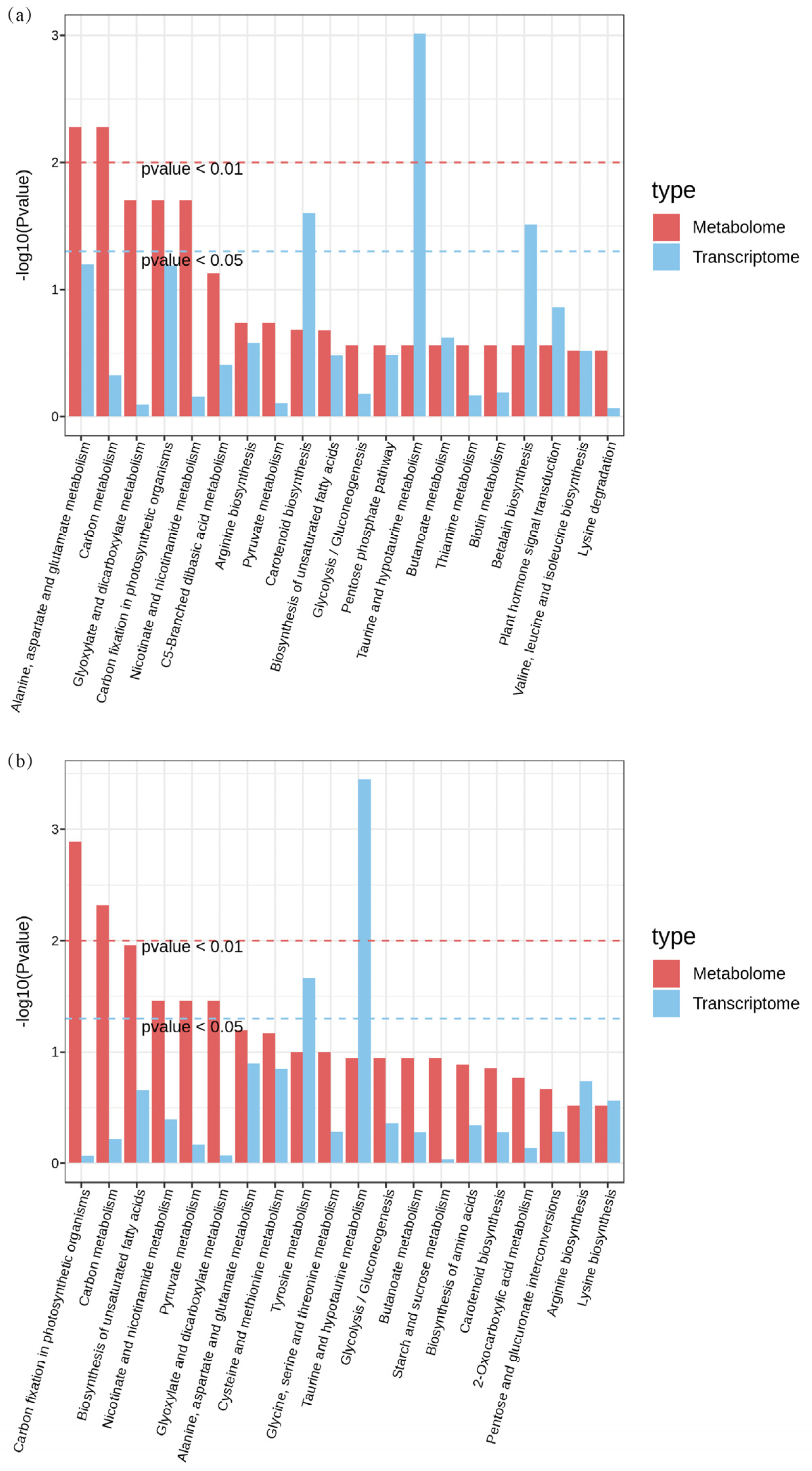
3.5. Integrative Analysis of Transcriptome and Metabolome of Rice Seedling Roots Under Different Carbonate Stresses
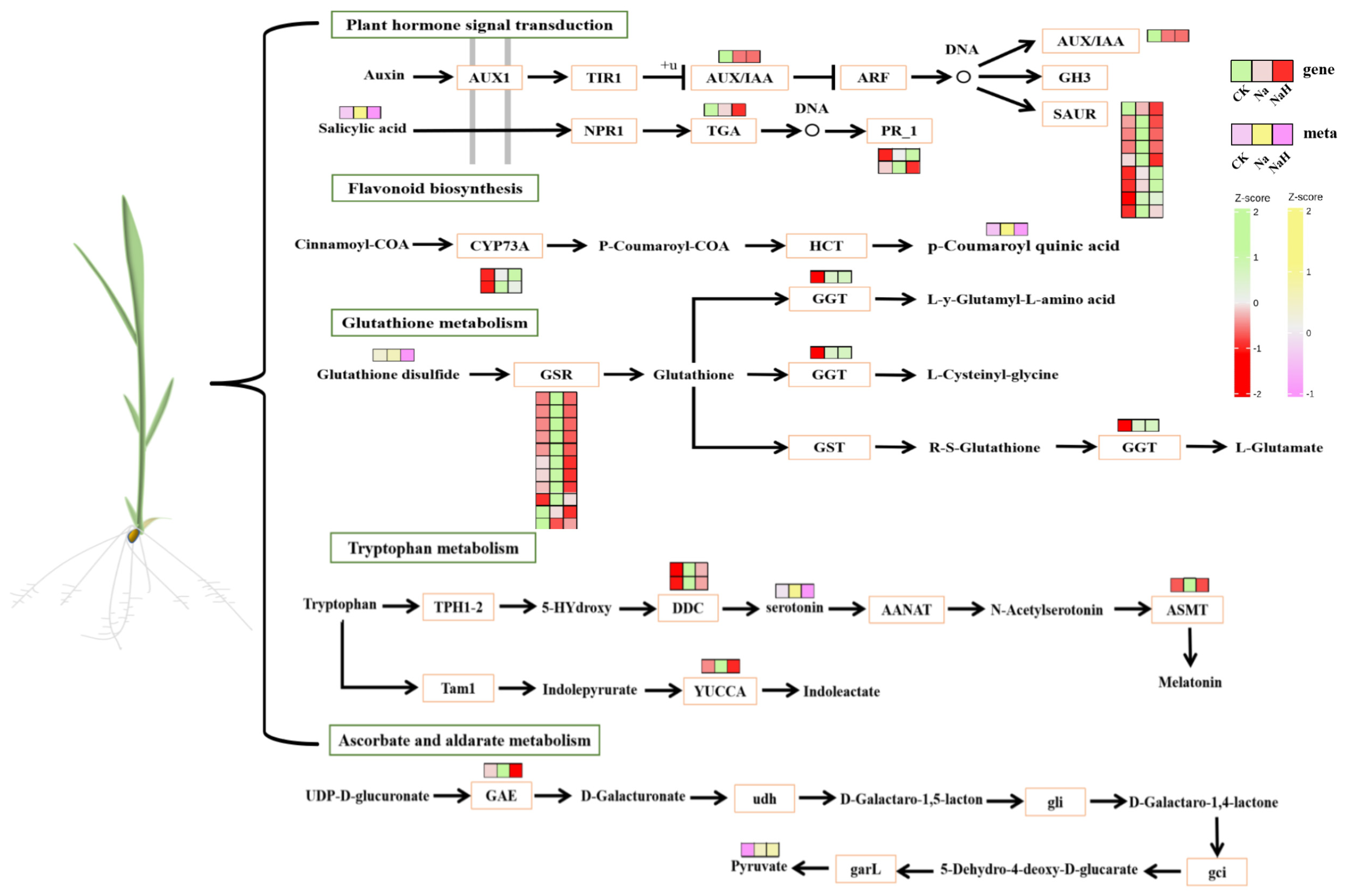
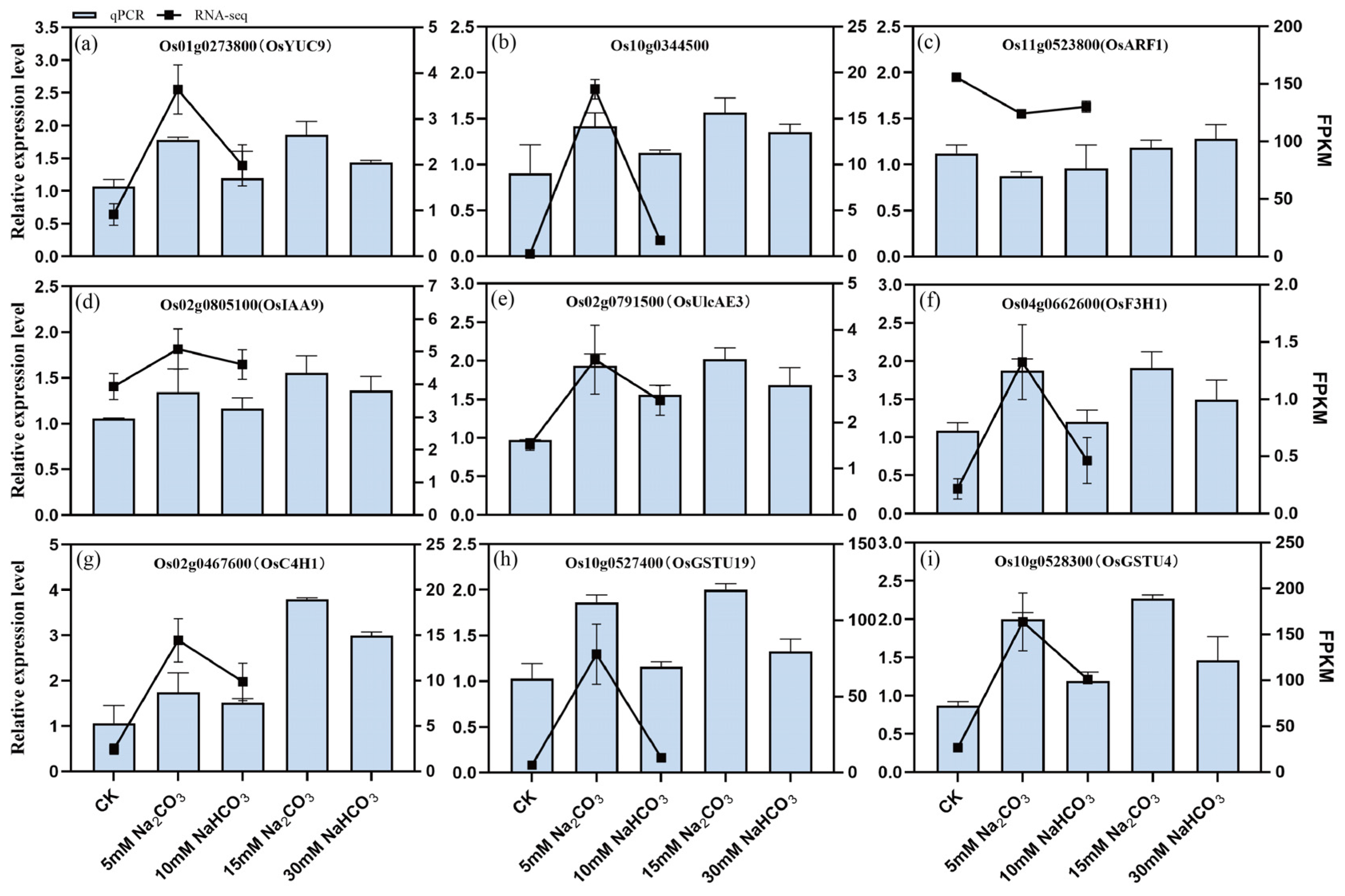
3.6. Verification of DAMs and DEGs Related to Auxin, Ascorbate, and Flavonoid Biosynthesis and Glutathione Metabolism in Rice Seedling Roots Under Different Carbonate Stresses
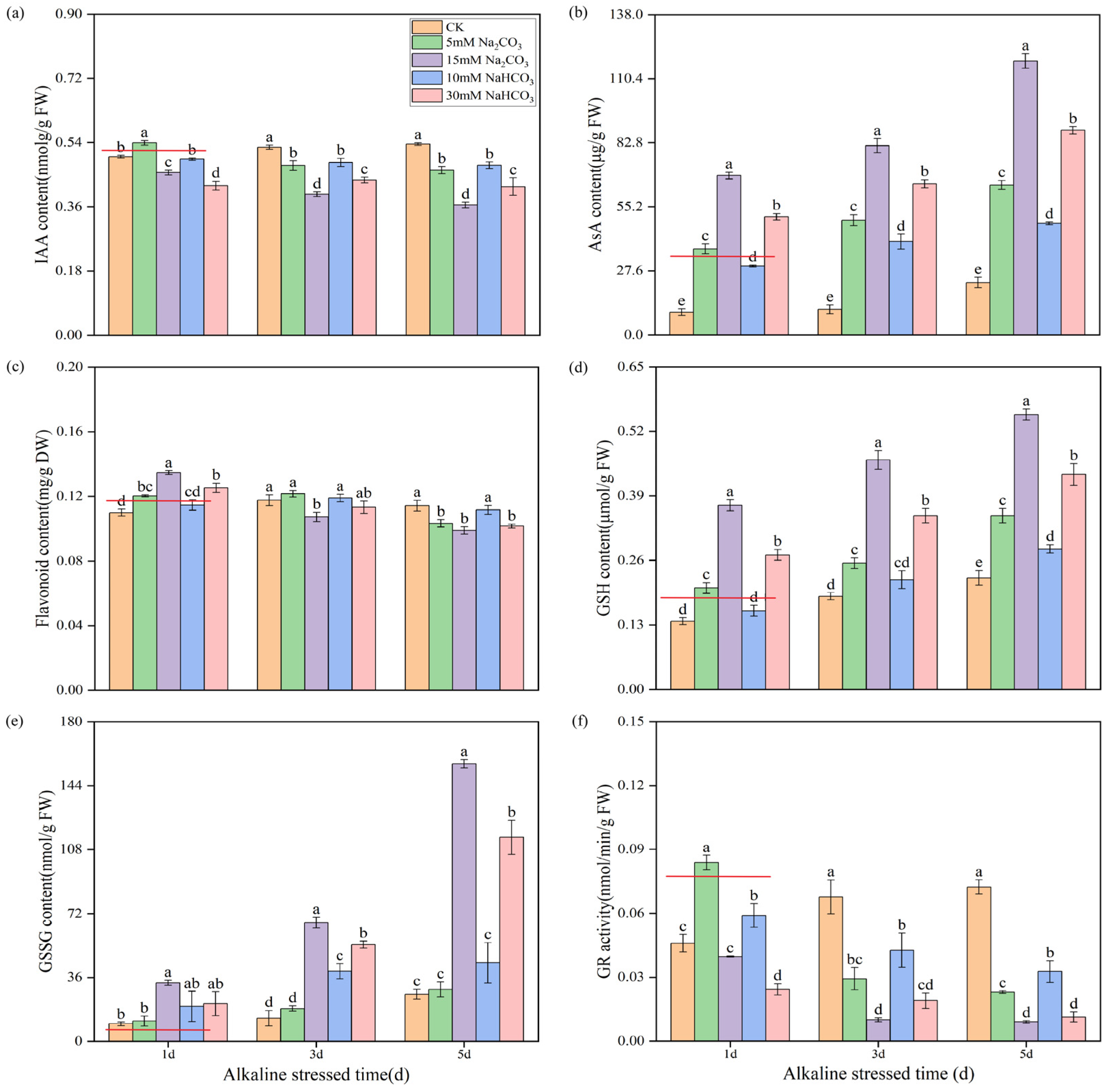
3.6.1. Effects of Different Carbonate Stresses on the Synthesis of Auxin and Antioxidants in Rice Seedling Roots
3.6.2. Effects of Different Carbonate Stresses on the Expression of Genes Related to Auxin, Ascorbate, and Flavonoid Biosynthesis and Glutathione Metabolism in Rice Seedling Roots
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, J.T.; Mu, C.S. Effects of saline and alkaline stresses on the germination, growth, photosynthesis, ionic balance and anti-oxidant system in an alkali-tolerant leguminous forage Lathyrus quinquenervius. Soil Sci. Plant Nutr. 2009, 55, 685–697. [Google Scholar] [CrossRef]
- Zhang, J.L.; Flowers, T.J.; Wang, S.M. Mechanisms of sodium uptake by roots of higher plants. Plant Soil 2010, 326, 45–60. [Google Scholar] [CrossRef]
- Jiang, C.J.; Liang, Z.W.; Xie, X.Z. Priming for saline-alkaline tolerance in rice: Current knowledge and future challenges. Rice Sci. 2023, 30, 417–425. [Google Scholar]
- Liu, L.L.; Wang, B. Protection of halophytes and their uses for cultivation of saline- alkali soil in China. Biology 2021, 10, 353. [Google Scholar] [CrossRef]
- Huang, L.H.; Liu, Y.; Ferreira, J.F.S.; Wang, M.M.; Na, J.; Huang, J.X.; Liang, Z.W. Long-term combined effects of tillage and rice cultivation with phosphogypsum or farmyard manure on the concentration of salts, minerals, and heavy metals of saline-sodic paddy fields in Northeast China. Soil Tillage Res. 2022, 215, 105222. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, X.L.; Zhang, R.X.; Yuan, H.Y.; Wang, M.M.; Yang, H.Y.; Ma, H.Y.; Liu, D.; Jiang, C.J.; Liang, Z.W. Root Damage under Alkaline Stress Is Associated with Reactive Oxygen Species Accumulation in Rice (Oryza sativa L.). Front. Plant Sci. 2017, 8, 1580. [Google Scholar] [CrossRef]
- Qian, G.T.; Wang, M.Y.; Wang, X.T.; Liu, K.; Li, Y.; Bu, Y.Y.; Li, L.X. Integrated Transcriptome and metabolome analysis of rice leaves response to high saline-alkali stress. Int. J. Mol. Sci. 2023, 24, 4062. [Google Scholar] [CrossRef]
- Ma, H.Y.; Liang, Z.W. Effects of different soil pH and soil extracts on the germination and seedling growth of Leymus chinensis. Chin. Bull. Bot. 2007, 24, 181–188. [Google Scholar]
- Lin, Y.J.; Ma, J.; Wu, N.; Qi, F.; Peng, Z.W.; Nie, D.D.; Yao, R.R.; Qi, X.; Slaski, J.; Yang, F.; et al. Transcriptome study of rice roots status under high alkaline stress at seedling stage. Agronomy 2022, 12, 925. [Google Scholar] [CrossRef]
- Nampei, M.; Jiadkong, K.; Chuamnakthong, S.; Wangsawang, T.; Sreewongchai, T.; Ueda, A. Different rhizospheric pH conditions affect nutrient accumulations in rice under salinity stress. Plants 2021, 10, 1295. [Google Scholar] [CrossRef]
- Li, Y.Y.; Huang, L.H.; Zhang, H.; Wang, M.M.; Liang, Z.W. Assessment of ammonia volatilization losses and nitrogen utilization during the rice growing season in alkaline salt-affected soils. Sustainability 2017, 9, 132. [Google Scholar] [CrossRef]
- Tian, Z.J.; Li, J.P.; Jia, X.Y.; Yang, F.; Wang, Z.C. Assimilation and translocation of dry matter and phosphorus in rice varieties affected by saline-alkaline stress. Sustainability 2016, 8, 568. [Google Scholar] [CrossRef]
- Lv, B.S.; Li, X.W.; Ma, H.Y.; Sun, Y.; Wei, L.X.; Jiang, C.J.; Liang, Z.W. Differences in growth and physiology of rice in response to different saline-alkaline stress factors. Agron. J. 2013, 105, 1119–1128. [Google Scholar] [CrossRef]
- Shaban, A.S.; Safhi, F.A.; Fakhr, M.A.; Pruthi, R.; Abozahra, M.S.; El-Tahan, A.M.; Subudhi, P.K. Comparison of the morpho-physiological and molecular responses to salinity and alkalinity stresses in rice. Plants 2023, 13, 60. [Google Scholar] [CrossRef]
- Lu, X.; Chen, G.; Ma, L.; Zhang, C.; Yan, H.; Bao, J.; Nai, G.; Wang, W.; Chen, B.; Ma, S.; et al. Integrated transcriptome and metabolome analysis reveals antioxidant machinery in grapevine exposed to salt and alkali stress. Physiol. Plant. 2023, 175, e13950. [Google Scholar] [CrossRef]
- Yang, Y.Q.; Guo, Y. Unraveling salt stress signaling in plants. J. Integr. Plant Biol. 2018, 60, 796–804. [Google Scholar] [CrossRef]
- Liu, X.L.; Xie, X.Z.; Zheng, C.K.; Wei, L.X.; Li, X.W.; Jin, Y.Y.; Zhang, G.H.; Jiang, C.J.; Liang, Z.W. RNAi-mediated suppression of the abscisic acid catabolism gene OsABA8ox1 increases abscisic acid content and tolerance to saline–alkaline stress in rice (Oryza sativa L.). Crop J. 2021, 10, 354–367. [Google Scholar] [CrossRef]
- Lu, X.; Ma, L.; Zhang, C.; Yan, H.; Bao, J.; Gong, M.; Wang, W.; Li, S.; Ma, S.; Chen, B. Grapevine (Vitis vinifera) responses to salt stress and alkali stress: Transcriptional and metabolic profiling. BMC Plant Biol. 2022, 22, 528. [Google Scholar] [CrossRef]
- Wei, L.X.; Lv, B.S.; Wang, M.M.; Ma, H.Y.; Yang, H.Y.; Liu, X.L.; Jiang, C.J.; Liang, Z.W. Priming effect of abscisic acid on alkaline stress tolerance in rice (Oryza sativa L.) seedlings. Plant Physiol. Biochem. 2015, 90, 50–57. [Google Scholar] [CrossRef]
- Feng, Z.H.; Lu, G.R.; Sun, M.; Jin, Y.Y.; Xu, Y.; Liu, X.L.; Wang, M.M.; Liu, M.; Yang, H.Y.; Guan, Y.; et al. Comparative study of the priming effect of abscisic acid on tolerance to saline and alkaline stresses in rice seedlings. Agronomy 2023, 13, 18. [Google Scholar] [CrossRef]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Raihan, M.R.H.; Masud, A.A.C.; Rahman, K.; Nowroz, F.; Rahman, M.; Nahar, K.; Fujita, M. Regulation of reactive oxygen species and antioxidant defense in plants under salinity. Int. J. Mol. Sci. 2021, 22, 9326. [Google Scholar] [CrossRef] [PubMed]
- Sachdev, S.; Ansari, S.A.; Ansari, M.I.; Fujita, M.; Hasanuzzaman, M. Abiotic stress and reactive oxygen species: Generation, signaling, and defense mechanisms. Antioxidants 2021, 10, 277. [Google Scholar] [CrossRef]
- Jia, X.M.; Zhu, Y.F.; Hu, Y.; Zhang, R.; Cheng, L.; Zhu, Z.L.; Zhao, T.; Zhang, X.Y.; Wang, Y.X. Integrated physiologic, proteomic, and metabolomic analyses of Malus halliana adaptation to saline-alkali stress. Hortic. Res. 2019, 6, 91. [Google Scholar] [CrossRef]
- Ma, Q.; Wang, H.L.; Wu, E.G.; Yuan, Y.H.; Feng, Y.; Zhao, L.; Feng, B.L. Comprehensive physiological, transcriptomic, and metabolomic analysis of the response of Panicum miliaceum L. roots to alkaline stress. Land Degrad. Dev. 2023, 34, 2912–2930. [Google Scholar] [CrossRef]
- Sharma, M.; Mahajan, P.; Singh, H.P.; Batish, D.R.; Kohli, R.K. 24-Epibrassinolide pre-treatment reduces alkaline-induced oxidative stress in red rice seedlings. Environ. Sci. Pollut. Res. 2019, 26, 23192–23197. [Google Scholar] [CrossRef]
- Shen, T.; Yan, R.J.; Xu, F.J.; Wang, Q.W.; Chen, D.; Li, K.Y.; Ni, L.; Jiang, M.Y. The NADPH oxidase OsRbohD and OsRbohH negatively regulate saline-alkaline tolerance in rice. Environ. Exp. Bot. 2023, 213, 105445. [Google Scholar] [CrossRef]
- Li, N.; Liu, H.; Sun, J.; Zheng, H.L.; Wang, J.G.; Yang, L.M.; Zhao, H.W.; Zou, D.T. Transcriptome analysis of two contrasting rice cultivars during alkaline stress. Sci. Rep. 2018, 8, 9586. [Google Scholar] [CrossRef]
- Ren, X.N.; Fan, J.H.; Li, X.; Shan, Y.; Wang, L.L.; Ma, L.J.; Li, Y.Y.; Li, X.M. Application of RNA sequencing to understand the response of rice seedlings to salt-alkali stress. BMC Genom. 2023, 24, 21. [Google Scholar] [CrossRef]
- Wang, J.; Lin, C.; Han, Z.; Fu, C.; Huang, D.; Cheng, H. Dissolved nitrogen in salt affected soils reclaimed by planting rice: How is it influenced by soil physicochemical properties. Sci. Total Environ. 2022, 824, 153863. [Google Scholar] [CrossRef] [PubMed]
- Bipul, S.; Pratim, B.; Abir, D.; Sayan, P.; Mirza, H.; Malay, K.A. Abscisic acid priming confers salt tolerance in maize seedlings by modulating osmotic adjustment, bond energies, ROS homeostasis, and organic acid metabolism. Plant Physiol. Biochem. 2023, 202, 107980. [Google Scholar]
- Guo, R.; Zhou, Z.Y.; Cai, R.; Liu, L.; Wang, R.X.; Sun, Y.G.; Wang, D.; Yan, Z.; Guo, C.H. Metabolomic and physiological analysis of alfalfa (Medicago sativa L.) in response to saline and alkaline stress. Plant Physiol. Biochem. 2024, 207, 108338. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, Y.; Wang, Y.; Du, F.; Zhang, Y.; Yin, M.; Zhao, X.; Xu, J.; Yang, Y.; Wang, W. Transcriptome and Metabolome Analyses Reveal Complex Molecular Mechanisms Involved in the Salt Tolerance of Rice Induced by Exogenous Allantoin. Antioxidants 2022, 11, 2045. [Google Scholar] [CrossRef]
- Elstner, E.F.; Heupel, A. Inhibition of nitrite formation from hydroxyl ammonium chloride: A simple assay for superoxide dismutase. Anal. Biochem. 1976, 70, 616–620. [Google Scholar] [CrossRef]
- Gao, Y.; Dong, X.; Wang, R.J.; Hao, F.; Zhang, H.; Zhang, Y.Y.; Lin, G.L. Exogenous Calcium Alleviates Oxidative Stress Caused by Salt Stress in Peanut Seedling Roots by Regulating the Antioxidant Enzyme System and Flavonoid Biosynthesis. Antioxidants 2024, 13, 233. [Google Scholar] [CrossRef]
- Jia, Z.S.; Tang, M.C.; Wu, J.M. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar]
- Du, J.; Cullen, J.J.; Buettner, G.R. Ascorbic acid: Chemistry, biology and the treatment of cancer. Biochim. Biophy. Acta. 2012, 1826, 443–457. [Google Scholar] [CrossRef]
- Nishimoto, S.; Koike, S.; Inoue, N.; Suzuki, T.; Ogasawara, Y. Activation of Nrf2 attenuates carbonyl stress induced by methylglyoxal in human neuroblastoma cells: Increase in GSH levels is a critical event for the detoxification mechanism. Biochem. Biophy. Res. Commun. 2017, 483, 874. [Google Scholar] [CrossRef]
- Rangani, J.K.; Parida, A.K.; Panda, A.; Kumari, A. Coordinated Changes in Antioxidative Enzymes Protect the Photosynthetic Machinery from Salinity Induced Oxidative Damage and Confer Salt Tolerance in an Extreme Halophyte Salvadora persica L. Front. Plant Sci. 2016, 7, 50. [Google Scholar] [CrossRef]
- Muchate, N.S.; Nikalje, G.C.; Rajurkar, N.S.; Suprasanna, P.; Nikam, T.D. Plant Salt Stress: Adaptive Responses, Tolerance Mechanism and Bioengineering for Salt Tolerance. Bot. Rev. 2016, 82, 371–406. [Google Scholar] [CrossRef]
- Gao, Y.G.; Jin, Y.L.; Guo, W.; Xue, Y.W.; Yu, L.H. Metabolic and Physiological Changes in the Roots of Two Oat Cultivars in Response to Complex Saline-Alkali Stress. Front. Plant Sci. 2022, 13, 835414. [Google Scholar] [CrossRef] [PubMed]
- Musazade, E.; Zhao, Z.; Shang, Y.; He, J.X.; Wang, Z.H.; Wu, M.; Xu, M.; Guo, L.Q.; Feng, X.Z. Differential Responses of Rice Seedlings to Salt and Alkaline Stresses: Focus on Antioxidant Defense, Organic Acid Accumulation, and Hormonal Regulation. J. Soil Sci. Plant Nutr. 2025. [Google Scholar] [CrossRef]
- Su, X.; Yao, L.; Wang, X.; Zhang, Y.; Zhang, G.F.; Li, X.J. Mechanisms for cell survival during abiotic stress: Focusing on plasma membrane. Stress Biol. 2025, 5, 1. [Google Scholar] [CrossRef]
- Stoeva, N.; Berova, M.; Zlatev, Z. Physiological Response of Maize to Arsenic Contamination. Biol. Plant 2003, 46, 449–452. [Google Scholar] [CrossRef]
- Zhang, H.Z.; Guo, Q.J.; Yang, J.X.; Shen, J.X.; Chen, T.B.; Zhu, G.X.; Chen, H.; Shao, C.Y. Subcellular cadmium distribution and antioxidant enzymatic activities in the leaves of two castor (Ricinus communis L.) cultivars exhibit differences in Cd accumulation. Ecotox. Environ. Saf. 2015, 120, 184–192. [Google Scholar] [CrossRef]
- Genisel, M.; Erdal, S.; Kizilkaya, M. The mitigating effect of cysteine on growth inhibition in salt-stressed barley seeds is related to its own reducing capacity rather than its effects on antioxidant system. Plant Growth Regul. 2015, 75, 187–197. [Google Scholar] [CrossRef]
- Zhang, H.; Li, D.S.; Zhou, Z.G.; Zahoor, R.; Chen, B.L.; Meng, Y.L. Soil water and salt affect cotton (Gossypium hirsutum L.) photosynthesis, yield and fiber quality in coastal saline soil. Agric. Water Manag. 2017, 187, 112–121. [Google Scholar] [CrossRef]
- Luo, X.F.; Dai, Y.J.; Zheng, C.; Yang, Y.Z.; Chen, W.; Wang, Q.C.; Chandrasekaran, U.; Du, J.B.; Liu, W.G.; Shu, K. The ABI4-RbohD/VTC2 regulatory module promotes reactive oxygen species (ROS) accumulation to decrease seed germination under salinity stress. New Phytol. 2021, 229, 950–962. [Google Scholar] [CrossRef]
- Li, J.; Chen, L.; Chen, Q.; Miao, Y.H.; Peng, Z.; Huang, B.S.; Guo, L.P.; Liu, D.H.; Du, H.Z. Allelopathic effect of Artemisia argyi on the germination and growth of various weeds. Sci. Rep. 2021, 11, 4303. [Google Scholar] [CrossRef]
- Wang, J.Y.; Hu, K.K.; Wang, J.E.; Gong, Z.Y.; Li, S.M.; Deng, X.X.; Li, Y.S. Integrated Transcriptomic and Metabolomic Analyses Uncover the Differential Mechanism in Saline–Alkaline Tolerance between Indica and Japonica Rice at the Seedling Stage. Int. J. Mol. Sci. 2023, 24, 12387. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.L.; Liu, J.; Meng, W.N.; Sun, Z.G.; Tan, Y.L.; Liu, Y.; Tan, M.P.; Wang, B.X.; Yang, J.C. Integrated Analysis of Transcriptome and Metabolome Reveals Molecular Mechanisms of Rice with Different Salinity Tolerances. Plants 2023, 12, 3359. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.C.; Pang, J.Y.; Zhang, F.H.; Sun, L.P.; Yang, L.; Zhao, Y.G.; Yang, Y.; Wang, Y.J.; Kadambot, H.M.; Siddique, K.H. Integrated transcriptomics and metabolomics analysis to characterize alkali stress responses in canola (Brassica napus L.). Plant Physiol. Biochem. 2021, 166, 605–620. [Google Scholar] [CrossRef] [PubMed]
- Mano, Y.; Nemoto, K. The pathway of auxin biosynthesis in plants. J. Exp. Bot. 2012, 63, 2853–2872. [Google Scholar] [CrossRef]
- Li, J.; Xu, H.H.; Liu, W.C.; Zhang, X.W.; Lu, Y.T. Ethylene Inhibits Root Elongation during Alkaline Stress through AUXIN1 and Associated Changes in Auxin Accumulation. Plant Physiol. 2015, 168, 1777–1791. [Google Scholar] [CrossRef]
- Ma, S.Q.; Lv, L.; Meng, C.; Zhang, C.S.; Li, Y.Q. Integrative Analysis of the Metabolome and Transcriptome of Sorghum bicolor Reveals Dynamic Changes in Flavonoids Accumulation under Saline−Alkali Stress. J. Agric. Food Chem. 2020, 68, 14781–14789. [Google Scholar] [CrossRef]
- Raghvendra, S.; Annapurna, S.; Ragunathan, D.; Mukesh, J. Over-Expression of a Rice Tau Class Glutathione S-Transferase Gene Improves Tolerance to Salinity and Oxidative Stresses in Arabidopsis. PLoS ONE 2024, 9, e92900. [Google Scholar]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, Y.; Hao, F.; Li, J.; Zhang, B.; Li, Z.; Liu, T.; Gao, Y.; Niu, X.; Liu, X.; Zhang, H.; et al. Integrated Transcriptome and Metabolome Analyses Reveal Complex Oxidative Damage Mechanisms in Rice Seedling Roots Under Different Carbonate Stresses. Antioxidants 2025, 14, 658. https://doi.org/10.3390/antiox14060658
Cao Y, Hao F, Li J, Zhang B, Li Z, Liu T, Gao Y, Niu X, Liu X, Zhang H, et al. Integrated Transcriptome and Metabolome Analyses Reveal Complex Oxidative Damage Mechanisms in Rice Seedling Roots Under Different Carbonate Stresses. Antioxidants. 2025; 14(6):658. https://doi.org/10.3390/antiox14060658
Chicago/Turabian StyleCao, Yang, Fei Hao, Jingpeng Li, Bolun Zhang, Zeming Li, Tiantian Liu, Yan Gao, Xuguang Niu, Xiaohu Liu, Hui Zhang, and et al. 2025. "Integrated Transcriptome and Metabolome Analyses Reveal Complex Oxidative Damage Mechanisms in Rice Seedling Roots Under Different Carbonate Stresses" Antioxidants 14, no. 6: 658. https://doi.org/10.3390/antiox14060658
APA StyleCao, Y., Hao, F., Li, J., Zhang, B., Li, Z., Liu, T., Gao, Y., Niu, X., Liu, X., Zhang, H., & Yang, L. (2025). Integrated Transcriptome and Metabolome Analyses Reveal Complex Oxidative Damage Mechanisms in Rice Seedling Roots Under Different Carbonate Stresses. Antioxidants, 14(6), 658. https://doi.org/10.3390/antiox14060658