
Protective Effect of Zeaxanthin from Lycium barbarum L. on Ultraviolet B-Induced Skin Photodamage in Mice Through Nrf2-Related Pathway


Abstract
1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Preparation of Zeaxanthin
2.3. Experimental Design
2.4. Histological Analyses
2.5. Assays and Kits
2.6. Immunofluorescence Staining
2.7. Western Blotting
2.8. Statistical Analysis
3. Results and Discussion
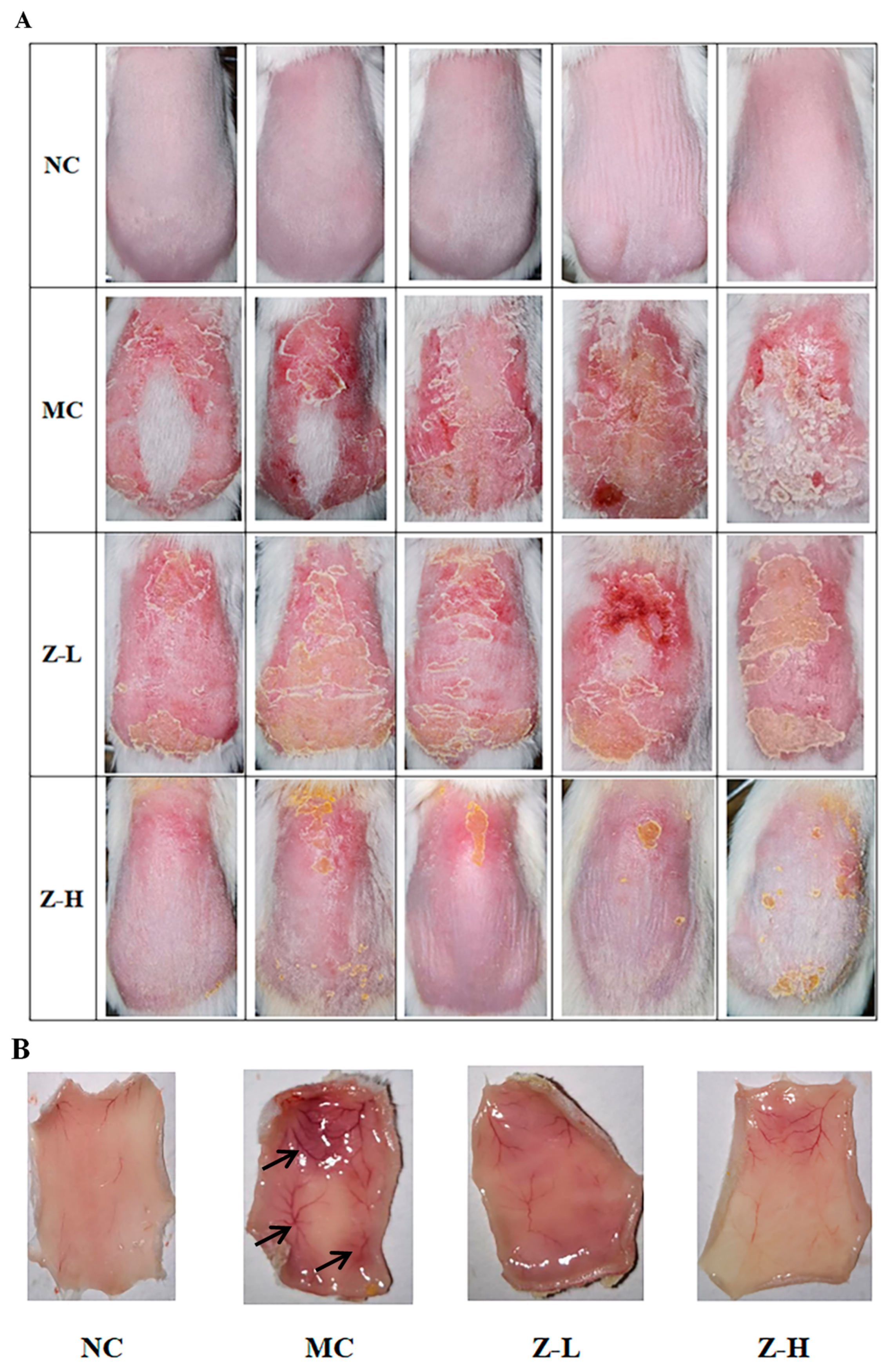
3.1. L. barbarum Zeaxanthin Alleviated UVB-Damaged Skin Appearance of Mice
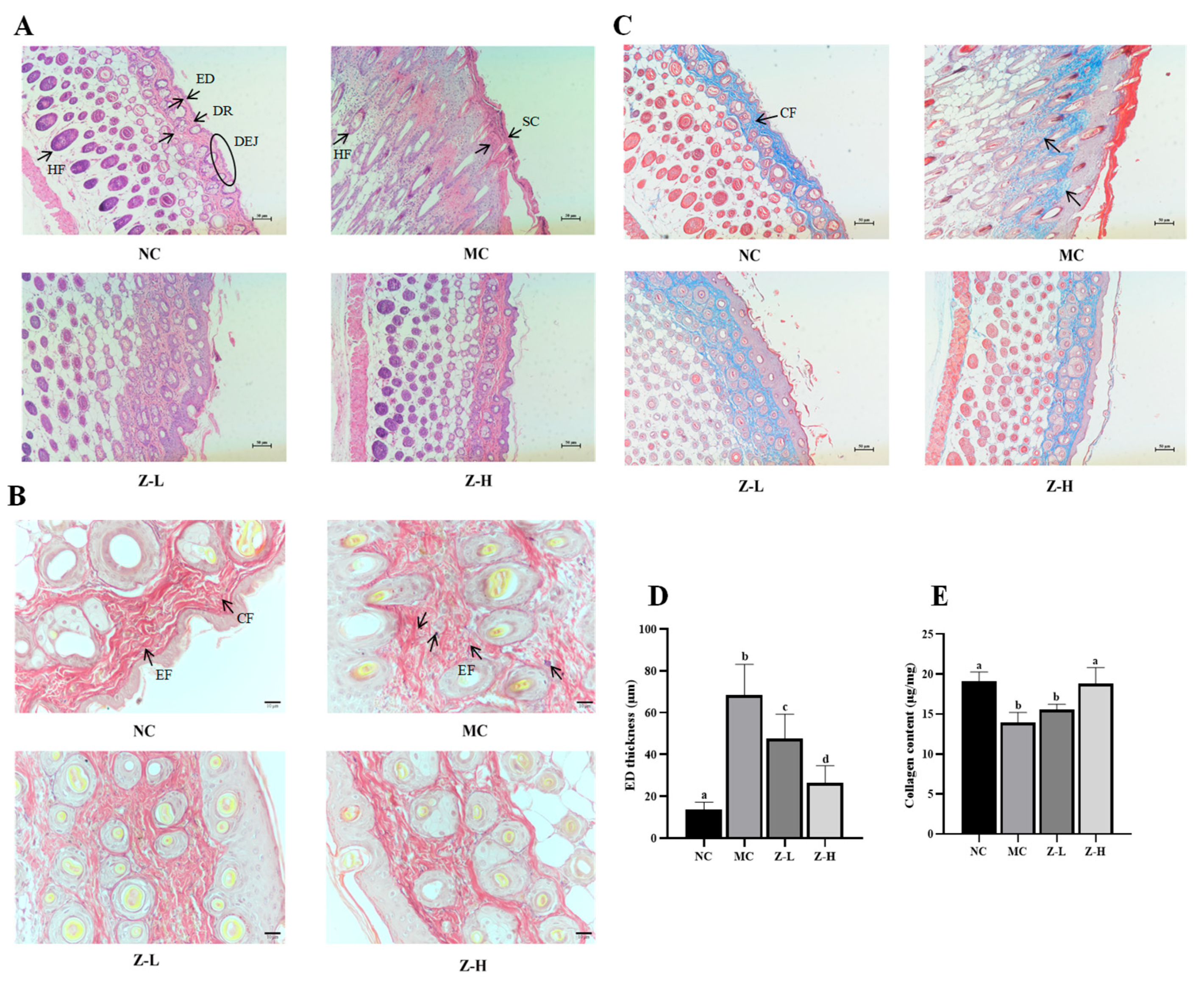
3.2. L. barbarum Zeaxanthin Improved UVB-Injured Skin Tissue Structure of Mice
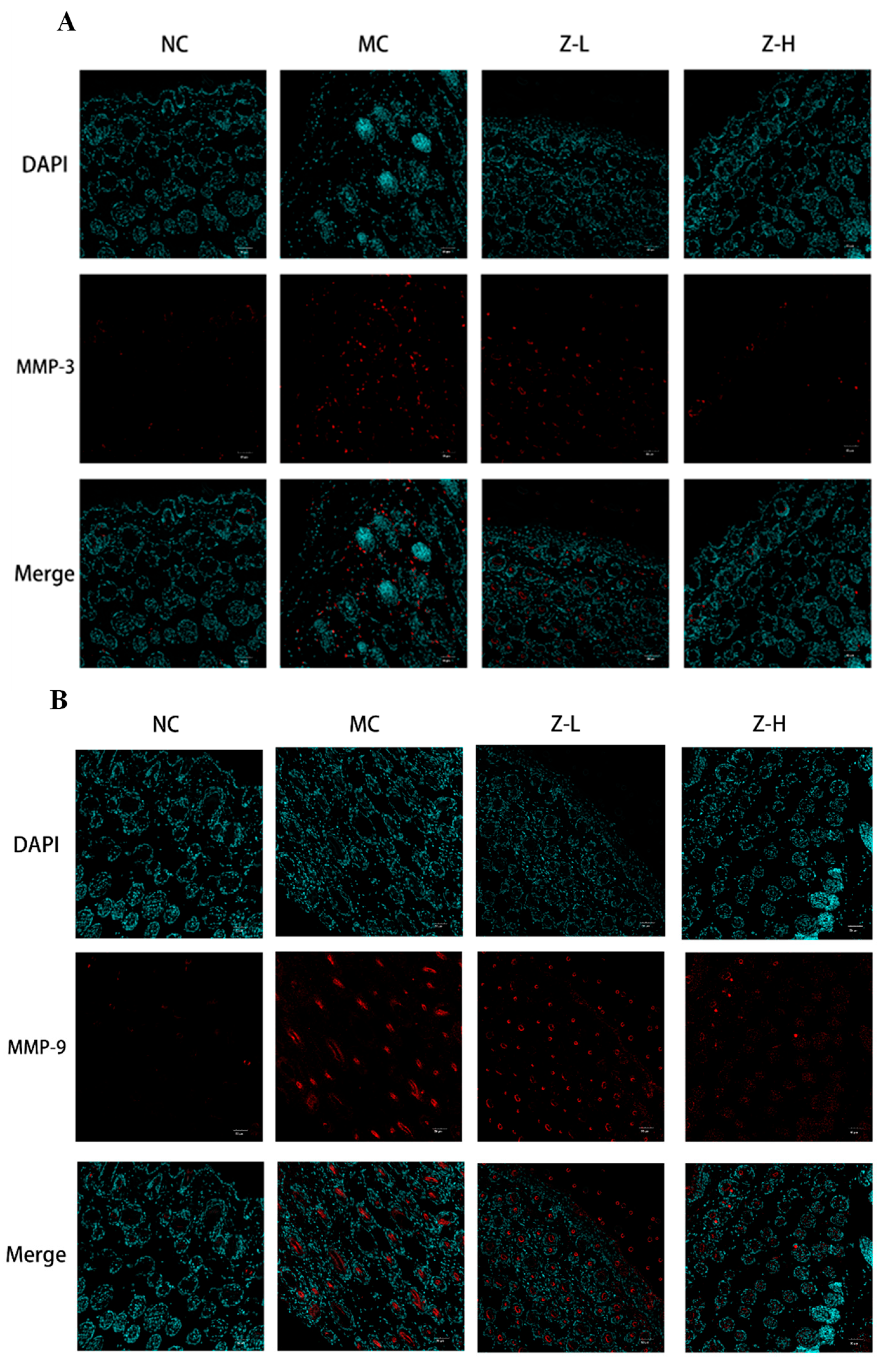
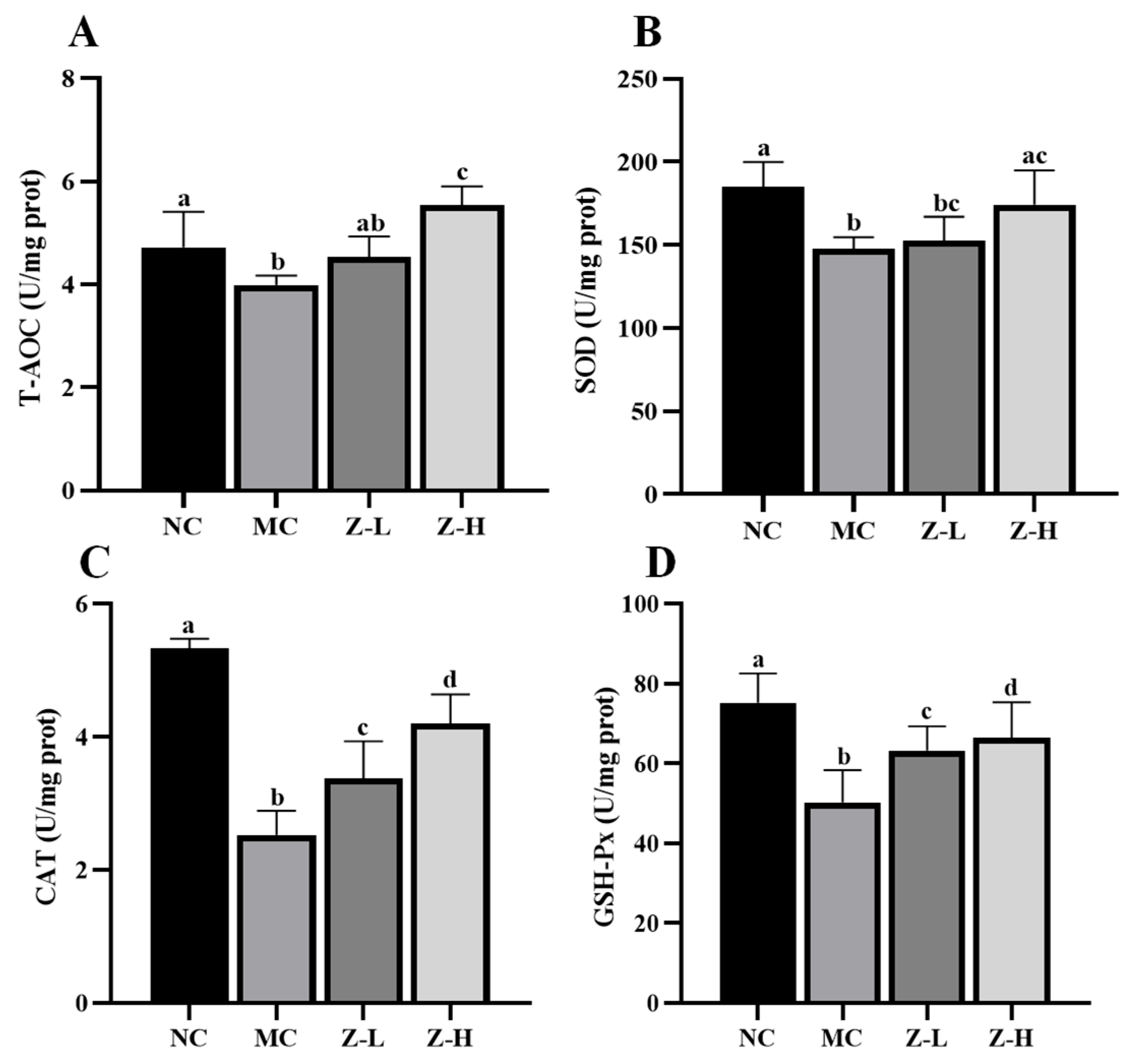
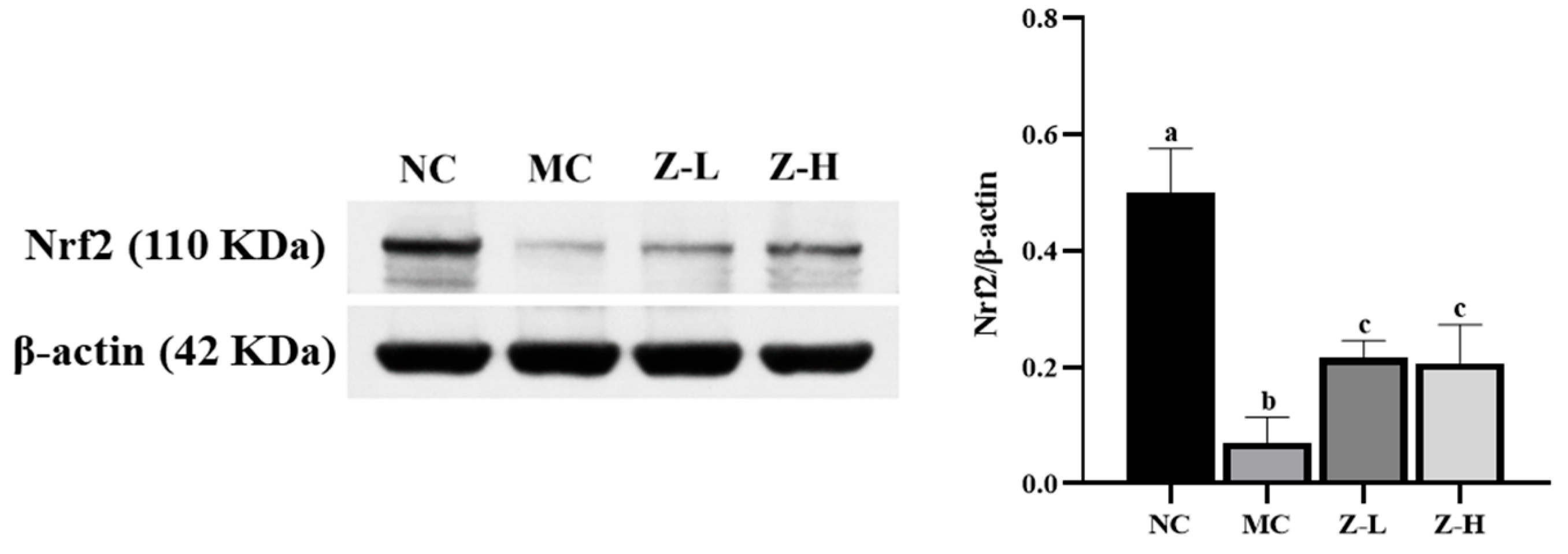
3.3. L. barbarum Zeaxanthin Inhibited UVB-Induced Oxidative Stress of Mouse Skin via Activating Nrf2
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huang, F.; Liu, Q.; Lu, Y. Magnolia biondii Flower Extract Attenuates UVB-Induced Skin Damage through High-Mobility Group Box Protein B1. Int. J. Cosmet. Sci. 2024, 46, 775–785. [Google Scholar] [CrossRef]
- Le Digabel, J.; Filiol, J.; Lauze, C.; Redoules, D.; Josse, G. In Vivo Method for Evaluating Sunscreen Protection against High-Energy Visible Light. J. Eur. Acad. Dermatol. Venereol. 2023, 37, 6–11. [Google Scholar] [CrossRef]
- Pasupathi, M.; Natarajan, B.; Kumar, T. Enhanced Sun Protection Factor of Octocrylene with Green Tea and Bhringraj Extracts. Cutan. Ocul. Toxicol. 2024, 43, 134–147. [Google Scholar] [CrossRef]
- Abou-Dahech, M.; Boddu, S.H.S.; Bachu, R.D.; Babu, R.J.; Shahwan, M.; Al-Tabakha, M.M.; Tiwari, A.K. A Mini-Review on Limitations Associated with UV Filters. Arab. J. Chem. 2022, 15, 104212. [Google Scholar] [CrossRef]
- Ciazynska, M.; Olejniczak-Staruch, I.; Sobolewska-Sztychny, D.; Narbutt, J.; Skibinska, M.; Lesiak, A. Ultraviolet Radiation and Chronic Inflammation-Molecules and Mechanisms Involved in Skin Carcinogenesis: A Narrative Review. Life 2021, 11, 326. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Li, Y.; Liu, M.; Li, Z.; Wang, X.; Zhang, K.; Zhao, X.; Li, G.; Bian, L. Complex Coacervate-Derived Hydrogel with Asymmetric and Reversible Wet Bioadhesion for Preventing UV Light-Induced Morbidities. Bioact. Mater. 2023, 30, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Zha, J.; Pan, Y.; Liu, X.; Zhu, H.; Liu, Y.; Zeng, W. Exosomes from Hypoxia-Pretreated Adipose-Derived Stem Cells Attenuate Ultraviolet Light-Induced Skin Injury via Delivery of Circ-Ash1l. Photodermatol. Photoimmunol. Photomed. 2023, 39, 107–115. [Google Scholar] [CrossRef]
- Heck, D.E.; Vetrano, A.M.; Mariano, T.M.; Laskin, J.D. UVB Light Stimulates Production of Reactive Oxygen Species—Unexpected Role for Catalase. J. Biol. Chem. 2003, 278, 22432–22436. [Google Scholar] [CrossRef]
- Chen, X.; Yang, C.; Jiang, G. Research Progress on Skin Photoaging and Oxidative Stress. Postep. Dermatol. I Alergol. 2021, 38, 931–936. [Google Scholar] [CrossRef]
- Chen, J.; Liu, Y.; Zhao, Z.; Qiu, J. Oxidative Stress in the Skin: Impact and Related Protection. Int. J. Cosmet. Sci. 2021, 43, 495–509. [Google Scholar] [CrossRef]
- Honda, M.; Kageyama, H.; Zhang, Y.; Hibino, T.; Goto, M. Oral Supplementation with Z-Isomer-Rich Astaxanthin Inhibits Ultraviolet Light-Induced Skin Damage in Guinea Pigs. Mar. Drugs 2022, 20, 414. [Google Scholar] [CrossRef]
- Li, X.; Matsumoto, T.; Takuwa, M.; Saeed Ebrahim Shaiku Ali, M.; Hirabashi, T.; Kondo, H.; Fujino, H. Protective Effects of Astaxanthin Supplementation against Ultraviolet-Induced Photoaging in Hairless Mice. Biomedicines 2020, 8, 18. [Google Scholar] [CrossRef]
- Prasedya, E.S.; Martyasari, N.W.R.; Abidin, A.S.; Pebriani, S.A.; Ilhami, B.T.K.; Frediansyah, A.; Sunarwidhi, A.L.; Widyastuti, S.; Sunarpi, H. Macroalgae Sargassum Cristaefolium Extract Inhibits Proinflammatory Cytokine Expression in BALB/C Mice. Scientifica 2020, 2020, 9769454. [Google Scholar] [CrossRef]
- Luo, Q.; Mi, J.; Zhang, L.; Li, Y.; Li, X.; Huang, T.; Yan, Y.; Zhang, X. Research on Composition and Antioxidant Activity of Carotenoids from Lycium L. and Different Fruits & Vegetables. Food Res. Dev. 2015, 36, 39–42. [Google Scholar] [CrossRef]
- Inbaraj, B.S.; Lu, H.; Hung, C.F.; Wu, W.B.; Lin, C.L.; Chen, B.H. Determination of Carotenoids and Their Esters in Fruits of Lycium barbarum Linnaeus by HPLC-DAD-APCI-MS. J. Pharm. Biomed. Anal. 2008, 47, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Setiawan, F.; Nurdianti, L.; Sri, N. Formulation and Effectivity of the Antioxidant Gel Preparation Containing Zeaxanthin as Anti Aging. Int. J. Appl. Pharm. 2021, 13, 152–156. [Google Scholar] [CrossRef]
- He, M.; Deng, X.-Y.; Zhu, Y.-B.; Hao, J.; Kay, M.; Zhang, H.; Chen, J.J.; Chen, Z.-B. Mechanism of Astaxanthin Relieving Lipopolysaccharide (LPS)-Induced Acute Liver Injury in Mice. Cienc. Rural 2024, 54, e20230102. [Google Scholar] [CrossRef]
- Longoni, B.; Demontis, G.C. Polyunsaturated Lipids in the Light-Exposed and Prooxidant Retinal Environment. Antioxidants 2023, 12, 617. [Google Scholar] [CrossRef]
- Moreno, L.G.; Cesar, N.R.; Melo, D.S.; Figueiro, M.T.O.; dos Santos, E.C.; Evangelista-Silva, P.H.; Santos, C.; de Sousa Santos, C.; Costa, K.B.; Rocha-Vieira, E.; et al. A MUFA/Carotenoid-Rich Oil Ameliorated Insulin Resistance by Improving Inflammation and Oxidative Stress in Obese Rats. Mol. Cell. Endocrinol. 2024, 581, 112110. [Google Scholar] [CrossRef]
- Rani, E.A.; Janani, R.; Chonche, M.J.; Vallikannan, B. Lactucaxanthin Regulates the Cascade of Retinal Oxidative Stress, Endoplasmic Reticulum Stress and Inflammatory Signaling in Diabetic Rats. Ocul. Immunol. Inflamm. 2023, 31, 320–328. [Google Scholar] [CrossRef]
- Huang, Y.; Shi, C.; Li, J. The Protective Effect of Zeaxanthin on Human Limbal and Conjunctival Epithelial Cells against UV-Induced Cell Death and Oxidative Stress. Int. J. Ophthalmol. 2019, 12, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Lan, L.; Wang, S.; Duan, S.; Zhou, X.; Li, Y. Cordyceps militaris Carotenoids Protect Human Retinal Endothelial Cells against the Oxidative Injury and Apoptosis Resulting from H2O2. Evid.-Based Complement. Altern. Med. 2022, 2022, 1259093. [Google Scholar] [CrossRef]
- Palombo, P.; Fabrizi, G.; Ruocco, V.; Ruocco, E.; Fluhr, J.; Roberts, R.; Morganti, P. Beneficial Long-Term Effects of Combined Oral/Topical Antioxidant Treatment with the Carotenoids Lutein and Zeaxanthin on Human Skin: A Double-Blind, Placebo-Controlled Study. Skin Pharmacol. Physiol. 2007, 20, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Lee, Y.-M.; Lee, Y.Y.; Yeum, K.-J. Oat (Avena Sativa) Extract against Oxidative Stress-Induced Apoptosis in Human Keratinocytes. Molecules 2021, 26, 5564. [Google Scholar] [CrossRef]
- Shen, Q.; Zhu, T.; Wu, C.; Xu, Y.; Li, C. Ultrasonic-Assisted Extraction of Zeaxanthin from Lycium barbarum L. with Composite Solvent Containing Ionic Liquid: Experimental and Theoretical Research. J. Mol. Liq. 2022, 347, 118265. [Google Scholar] [CrossRef]
- Yang, Y.; Zhou, M.J.; Liu, R.C.; Xu, L.; Gao, S.S.; Tan, W.; Gu, Z.Y. Protective Mechanism of Total Flavonoids of Lavender Against Skin Photoaging in Mice. Chin. J. Exp. Tradit. Med. Formulae 2022, 28, 112–120. [Google Scholar]
- Bhattacharyya, T.K.; Barch, B.E.; Vargas, M.; Thomas, J.R. Cutaneous Injury Following Acute UV-B Radiation in a Mouse Model: A Pilot Histological Study. J. Histotechnol. 2013, 36, 37–44. [Google Scholar] [CrossRef]
- Khan, A.; Bai, H.L.; Khan, A.; Bai, Z.L. Neferine Prevents Ultraviolet Radiation-Induced Skin Photoaging. Exp. Ther. Med. 2020, 19, 3189–3196. [Google Scholar] [CrossRef]
- Neuman, R.E.; Logan, M.A. The Determination of Collagen and Elastin in Tissues. J. Biol. Chem. 1950, 186, 549–556. [Google Scholar] [CrossRef]
- Zhong, Q.-Y.; Luo, Q.-H.; Lin, B.; Lin, B.-Q.; Su, Z.-R.; Zhan, J.Y.-X. Protective Effects of Andrographolide Sodium Bisulfate on UV-Induced Skin Carcinogenesis in Mice Model. Eur. J. Pharm. Sci. 2022, 176, 106232. [Google Scholar] [CrossRef]
- Tohgasaki, T.; Nishizawa, S.; Kondo, S.; Ishiwatari, S.; Sakurai, T. Long Hanging Structure of Collagen VII Connects the Elastic Fibers and the Basement Membrane in Young Skin Tissue. J. Histochem. Cytochem. 2022, 70, 751–757. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Wang, M.; Zhang, X.; Chen, A.; Fei, Y.; Zhao, Q.; Guo, D.; Chen, H.; Zheng, S. Therapeutic Effect of Concentrated Growth Factor Preparation on Skin Photoaging in a Mouse Model. J. Int. Med. Res. 2020, 48, 300060520962946. [Google Scholar] [CrossRef]
- Lü, J.; Lin, H.; Qin, X.; Zhang, C.; Cao, W.; Gao, J.; Zheng, H. Preliminary Study on Resistance of Oyster and Its Enzymatic Hydrolysis Products to Skin Photoaging. South China Fish. Sci. 2021, 17, 91–100. [Google Scholar]
- Xu, C.; Xiong, Q.-W.; Li, Y.; Zhao, J.-N.; Zhang, L.; Li, X.-L. Explore the Multitarget Mechanism of Tetrahydrocurcumin Preventing on UV-Induced Photoaging Mouse Skin. Heliyon 2022, 8, e09888. [Google Scholar] [CrossRef]
- Zhi, Q.; Lei, L.; Li, F.; Zhao, J.; Yin, R.; Ming, J. The Anthocyanin Extracts from Purple-Fleshed Sweet Potato Exhibited Anti-Photoaging Effects on Ultraviolent B-Irradiated BALB/c-Nu Mouse Skin. J. Funct. Foods 2020, 64, 103640. [Google Scholar] [CrossRef]
- Kim, M.-K.; Shin, H.S.; Shin, M.H.; Kim, H.; Lee, D.H.; Chung, J.H. Dual Role of Enhancer of Zeste Homolog 2 in the Regulation of Ultraviolet Radiation-Induced Matrix Metalloproteinase-1 and Type I Procollagen Expression in Human Dermal Fibroblasts. Matrix Biol. 2023, 119, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-J.; Lee, K.B.; Hong, A.-Y.; Son, Y.H.; Lee, D.H.; Jeong, E.M.; Kim, I.-G. Transglutaminase 2 Mediates UVB-Induced Matrix Metalloproteinase-1 Expression by Inhibiting Nuclear P65 Degradation in Dermal Fibroblasts. Exp. Dermatol. 2022, 31, 743–752. [Google Scholar] [CrossRef]
- Shin, J.; Kim, J.-E.; Pak, K.-J.; Kang, J.I.; Kim, T.-S.; Lee, S.-Y.; Yeo, I.-H.; Park, J.H.Y.; Kim, J.H.; Kang, N.J.; et al. A Combination of Soybean and Haematococcus Extract Alleviates Ultraviolet B-Induced Photoaging. Int. J. Mol. Sci. 2017, 18, 682. [Google Scholar] [CrossRef]
- Tominaga, K.; Hongo, N.; Fujishita, M.; Takahashi, Y.; Adachi, Y. Protective Effects of Astaxanthin on Skin Deterioration. J. Clin. Biochem. Nutr. 2017, 61, 33–39. [Google Scholar] [CrossRef]
- Wang, X.-F.; Huang, Y.-F.; Wang, L.; Xu, L.-Q.; Yu, X.-T.; Liu, Y.-H.; Li, C.-L.; Zhan, J.Y.-X.; Su, Z.-R.; Chen, J.-N.; et al. Photo-Protective Activity of Pogostone against UV-Induced Skin Premature Aging in Mice. Exp. Gerontol. 2016, 77, 76–86. [Google Scholar] [CrossRef]
- Chen, N.; Hu, M.; Jiang, T.; Xiao, P.; Duan, J. Insights into the Molecular Mechanisms, Structure-Activity Relationships and Application Prospects of Polysaccharides by Regulating Nrf2-Mediated Antioxidant Response. Carbohydr. Polym. 2024, 333, 122003. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liang, Y.; Zhou, X.; Tian, Y.; Miao, Z.; Ko, C.-C.; Hu, X. Nrf2 Differentially Regulates Osteoclast and Osteoblast Differentiation for Bone Homeostasis. Biochem. Biophys. Res. Commun. 2023, 674, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Zheng, S.; Jin, J.; Wu, T.; Shi, Y.; Yang, K.; Zhang, H.; Li, Y.; Sun, Y.; Lv, Y.; et al. Polydopamine-Loaded Prunetin Nanomaterials Activate DRD2 to Reduce UV-Induced Inflammation by Stabilizing and Promoting Nrf2 Nuclear Translocation. ACTA Biomater. 2023, 169, 556–565. [Google Scholar] [CrossRef]
- Zhu, S.; Qin, W.; Liu, T.; Ma, H.; Hu, C.; Yue, X.; Yan, Y.; Lv, Y.; Wang, Z.; Zhao, Z.; et al. Modified Qing’e Formula Protects against UV-Induced Skin Oxidative Damage via the Activation of Nrf2/ARE Defensive Pathway. Front. Pharmacol. 2022, 13, 976473. [Google Scholar] [CrossRef] [PubMed]
- Fazekas, Z.; Saladi, R.N.; Gao, D.; Lu, Y.; Lebwohl, M.; Wei, H. Immunohistochemical Evaluation of Lycopene’s Protective Effect against Ultraviolet B (UVB)-Induced Photodamage in Vivo. J. Investig. Dermatol. 2003, 121, 207. [Google Scholar]
- Rodriguez-Luna, A.; Avila-Roman, J.; Luisa Gonzalez-Rodriguez, M.; Cozar, M.J.; Rabasco, A.M.; Motilva, V.; Talero, E. Fucoxanthin-Containing Cream Prevents Epidermal Hyperplasia and UVB-Induced Skin Erythema in Mice. Mar. Drugs 2018, 16, 378. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carotenoids | Protective Effect of Carotenoids on UVB-Damaged Mouse Skin |
|---|---|
| lycopene | reduces epidermal ornithine decarboxylase and caspase-3; inhibits tumor and apoptosis |
| β-carotene | enhances Nrf2 and HO-1; inhibits oxidative stress |
| fucoxanthin | enhances Nrf2 and HO-1; reduces COX-2; inhibits oxidative stress and inflammation |
| zeaxanthin | enhances Nrf2; reduces MMPs; inhibits oxidative stress |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, L.; Shen, Q.; Xu, Y.; Li, C. Protective Effect of Zeaxanthin from Lycium barbarum L. on Ultraviolet B-Induced Skin Photodamage in Mice Through Nrf2-Related Pathway. Antioxidants 2025, 14, 632. https://doi.org/10.3390/antiox14060632
Zhu L, Shen Q, Xu Y, Li C. Protective Effect of Zeaxanthin from Lycium barbarum L. on Ultraviolet B-Induced Skin Photodamage in Mice Through Nrf2-Related Pathway. Antioxidants. 2025; 14(6):632. https://doi.org/10.3390/antiox14060632
Chicago/Turabian StyleZhu, Lin, Qiruonan Shen, Yujuan Xu, and Chunmei Li. 2025. "Protective Effect of Zeaxanthin from Lycium barbarum L. on Ultraviolet B-Induced Skin Photodamage in Mice Through Nrf2-Related Pathway" Antioxidants 14, no. 6: 632. https://doi.org/10.3390/antiox14060632
APA StyleZhu, L., Shen, Q., Xu, Y., & Li, C. (2025). Protective Effect of Zeaxanthin from Lycium barbarum L. on Ultraviolet B-Induced Skin Photodamage in Mice Through Nrf2-Related Pathway. Antioxidants, 14(6), 632. https://doi.org/10.3390/antiox14060632